





Фузаріозна сажка або парша дрібних зерен викликається грибком Fusarium graminearum (Schwabe), хоча Нідерланди та інші регіони Центральної Європи повідомляють F. culmorum як найбільш поширений вид (Snidjers, 1989). У Польщі F. culmorum, F. graminearum і F. nivale показали схожі рівні вірулентності від помірного до сильного, тоді як F. avenaceum виявився середньо- та помірно вірулентним. Однак у кількох дослідженнях, спрямованих на ідентифікацію збудника, аж 18 Fusarium spp. були виділені та ідентифіковані (Mihuta-Grimm and Foster, 1989; Reis, 1985).
Стаття від L. ГілкрістПарша Х.Я. Дубіна поширена в теплих, вологих регіонах, де цвітіння збігається з періодами дощів. Захворюваність на цю хворобу зростала протягом останніх десяти років з різних причин. Мабуть, найважливішою причиною є збільшення площ, на яких пшениця сіється з кукурудзою або іншими зерновими культурами. Іншими причинами є зміни в системі землеробства з метою захисту ґрунтів та переміщення посівів пшениці з традиційних на більш вологі, нетрадиційні райони (Gilchrist et al., 1997).
Фузаріозна парша спричиняє значні виробничі втрати в усьому світі, які можуть сягати 50 відсотків. Аналогічно, в Парагваї погодні умови в 1972 і 1975 роках сприяли епідеміям фузаріозу і септоріозу, які разом призвели до втрат до 70 відсотків (Viedma, 1989). Польща, Нідерланди, Великобританія, колишня Чехословаччина, Російська Федерація, Франція та Австрія є деякими з європейських країн, які повідомляють про захворюваність паршею. Території Канади, США, Мексики, Гватемали, Бразилії, Еквадору, Уругваю та Аргентини в Америці сильно уражені фузаріозом (Ireta and Gilchrist, 1994).
Парша може спричинити значні втрати врожаю та якості, а також токсикози у тварин і людей (Ireta and Gilchrist, 1994; Baht et al., 1989; Luo, 1988; Snidjers, 1989; Marasas et al., 1988). Збитки від парші в Сполучених Штатах оцінювалися більш ніж в 1 млрд. доларів США в 1993 році і 1 млрд. 500 млн. доларів США в 1994 році. У Китаї, за оцінками, парша може вражати до 7 млн. га, і в роки епідемії може бути втрачено 2,5 млн. тонн зерна. Захворювання, пов'язані з фузаріозним мікотоксином у людей, були зареєстровані в Китаї, Індії та Японії, тоді як захворювання у тварин були зареєстровані в багатьох частинах світу (Dubin et al., 1997).
На сьогоднішній день у всьому світі існують правила щодо мікотоксинів (Van Egmond та Dekker, 1995). Однак у багатьох країнах ці норми не застосовуються, і багато людей, особливо в сільській місцевості, вживають зернові без будь-якого контролю, або у вигляді зерен, або опосередковано у вигляді м'яса, отриманого від тварин, яких годували зараженим зерном. У таблиці 16.1 показано основні ефекти на свиней і птицю, спричинені найбільш важливими токсинами, що виробляються деякими Фузаріоз видів.
Вологі та теплі погодні умови в період від колосіння до дозрівання посилюють ураженість паршею. Точкою проникнення F. graminearum є колос, особливо квіткові органи. Це впливає на зав'язування насіння та налив зерна. Заражені колоски швидко втрачають хлорофіл і стають блідими. Пізніше вони набувають рожевого або персикового кольору, особливо біля основи та країв колосків (Рис. 41). Якщо умови навколишнього середовища залишаються сприятливими, інфекція поширюється на сусідні колоски, а в деяких випадках може вразити весь колос, включаючи рахіс і його квітконіжку. За сильного розвитку інфекції пошкоджені зерна покриваються міцелієм і набувають вигляду рожевої ватоподібної маси. За помірного рівня розвитку хвороби зерно може бути зморщеним, малої ваги та білуватого кольору (надгробне зерно).
Основні токсичні ефекти на свиней та птицю, спричинені найбільш важливими токсинами, що виробляються деякими видами Fusarium
| Мікотоксин | Клінічні ознаки |
| Зеараленон | Набрякла червона вульва; випадіння піхви у свиней; у поросят-сисунів може спостерігатися збільшення вульви; проблеми з фертильністю |
| Ботоксин (дезоксиніваленол, DON) | Зниження споживання корму та приросту ваги у свиней з концентрацією ДОН >2 мг/кг корму; блювота; відмова від корму при дуже високих концентраціях ДОН (>20 мг/кг корму)a |
| Інші трихотецени Т-2 токсин HT-2 токсин Діацетоксісцирпенол | Більш токсичний, ніж DON; зниження споживання корму; блювота; подразнення шкіри та шлунково-кишкового тракту; нейротоксичність; ненормальне потомство; підвищена чутливість до хвороб; кровотечі |
| Охаратоксин | Вражає переважно проксимальні канальці нирок у свиней та птиці; нирки сильно збільшені та бліді; у птиці - жирова дистрофія печінки |
амг/кг = частин на мільйон (ppm).
Джерело: Тренгольм та ін., 1984.
Fusarium graminearum (Plate 42) може вражати рослини пшениці на всіх стадіях росту, викликаючи хвороби проростків, стебла та кореневі гнилі. Первинна інфекція може виникати через аскоспори або макроконідії, що осідають на колосках та видавлених пиляках. Для успішного зараження колосків макроконідіями зазвичай достатньо температури від 10° до 30°C і відносної вологості вище 95% протягом 40-60 годин (Ireta, 1989).
Fusarium graminearum є одним з небагатьох Фузаріоз вид, який утворює перитеції в польових умовах (Платівка 43, Платівка, 44). Перитеції представляють собою статеву стадію гриба Gibberella zeae і утворюються на пшеничній клейковині. Перитеції відіграють важливу роль у виживанні патогена з року в рік (Khonga and Sutton, 1988) і співіснують з міцелієм у залишках попередньої культури, утворюючи початкове джерело інокуляту для розвитку парші. Дослідження, проведені в Китаї, показали, що найнижча температура для розвитку перитеціїв становить від 7° до 10°C, а найбільш придатна - від 15° до 20°C. Найнижча вологість ґрунту для утворення перитеціїв становила 50-60%, а найбільш придатна - 70-80% (Wang, 1997).
Фактори, що визначають розвиток хвороби, включають клімат, рівень посівного матеріалу та стадію росту пшениці. Епідемії парші пшениці залежать в основному від кількості первинного, а не вторинного посівного матеріалу. Інвазія відбувається переважно в період цвітіння. Вторинне зараження конідіями (вклейка 45) відбувається після того, як на полі з'являються хворі колоски.
Пожнивні рештки та культурні практики відіграють важливу роль у збереженні F. graminearum і, як наслідок, епідемії. Інфекції на пшениці, висіяній на полях з кукурудзяними рештками, можуть бути в два-три рази сильнішими (Teich and Nelson, 1984). Загалом, патоген був сапрофітом лише на стерні рису в районах рисово-пшеничної сівозміни та на стеблах кукурудзи в посушливих районах Китаю (Wang, 1997). Якщо залишки заорюються, виживання перитеції знижується і зменшується первинне джерело інокуляту (Reis, 1989).
Fusarium graminearum є факультативним паразитом і патогенним для багатьох інших трав, включаючи звичайні бур'яни та зернові культури (жито, рис, ячмінь і тритикале). Якщо такі практики, як сівозміна з не-господарями або поводження з пожнивними рештками, не є ефективними окремо, вони можуть у комплексі зменшити джерело первинного збудника хвороби. Контроль хвороб ефективно ґрунтується на інтегрованому управлінні, що включає належні агрономічні практики, використання стійких або толерантних сортів і застосування хімічних засобів.
Біологія фузаріозної сажки
Випадкові патогени
ВГК викликається грибками роду Fusarium. Найпоширенішими видами, що викликають FHB, є Fusarium graminearum (сексуальна стадія - Gibberella zeae). Це той самий грибок, який часто асоціюється з стебловою гниллю кукурудзи. Ще один Фузаріоз видами, що викликають FHB, є Fusarium culmorum. Обидва. F. graminearum і F. culmorum також може викликати кореневу гниль дрібних зерен. На ячмені, два інших Фузаріоз видів, F. poae і F. avenaceumтакож може спричинити загнивання ядра.
Виживання та поширення
Гриб зберігається і розмножується на заражених пожнивних рештках дрібних зернових і кукурудзи. У вологу погоду спори грибів переносяться вітром або розбризкуються на качани зернових культур. Спори можуть потрапляти зсередини посіву або заноситися вітром з сусідніх посівів, іноді на значні відстані. Пшениця і тверда пшениця сприйнятливі до інфекції з періоду цвітіння (запилення) до стадії твердого тіста розвитку ядра. Спори гриба-збудника можуть потрапляти на відкриті пиляки під час цвітіння, а потім проростати в зерно, остюки або інші частини колоса. Для ярого ячменю, який цвіте, коли качан знаходиться в колосі, зараження найчастіше відбувається після періоду цвітіння, коли качан пробивається крізь листкову піхву. Інфекція в обох культурах може тривати до дозрівання зерна за сприятливих умов навколишнього середовища для організму (організмів).
Найсприятливішими умовами для інфікування є тривалі періоди (від 48 до 72 годин) високої вологості та високої температури (від 75 до 85 градусів за Фаренгейтом (від 24°C до 30°C)). Однак інфікування відбувається і при більш низьких температурах, коли висока вологість зберігається довше, ніж 72 години. Ранні інфекції можуть утворювати спори, що переносяться повітрям, які спричиняють вторинне поширення хвороби, особливо якщо культура має нерівномірне цвітіння через пізні кущіння.
Оскільки розвиток FHB залежить від сприятливих умов навколишнього середовища від цвітіння (поява качанів у ячменю) до розвитку ядра, частота виникнення та інтенсивність хвороби змінюється з року в рік. Комбінація факторів, які можуть призвести до значних втрат врожаю та якості, включає в себе: надлишок посівного матеріалу, тривалі або повторювані періоди вологості та високої вологості в період цвітіння (поява колоса у ячменю) до формування ядра, а також використання дуже сприйнятливого сорту.
Модель інфекції фузаріозної сажки
Відомо, що грибкові агенти, які належать до комплексу фузаріозної сажки пшениці, полюбляють теплі температури від 20°C до 30°C та тривалі вологі періоди. Кількаденні періоди вологості листя призводять до появи ранніх видимих симптомів. Але симптоми можуть з'явитися і після більш тривалого латентного періоду, якщо інфекція супроводжується періодом зволоження листя протягом 18 годин або навіть коротшого часу. Симптоми можуть з'явитися і після штучної інокуляції при температурі 15°C.
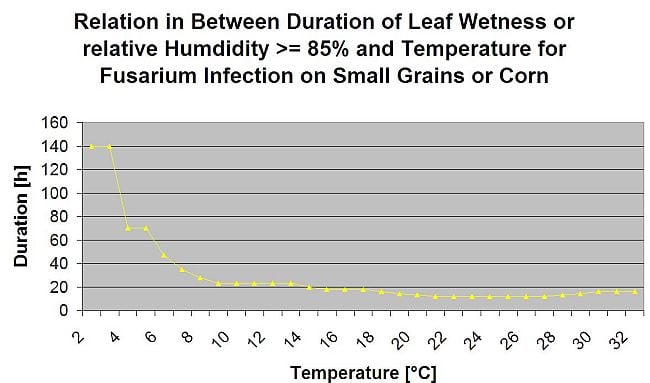

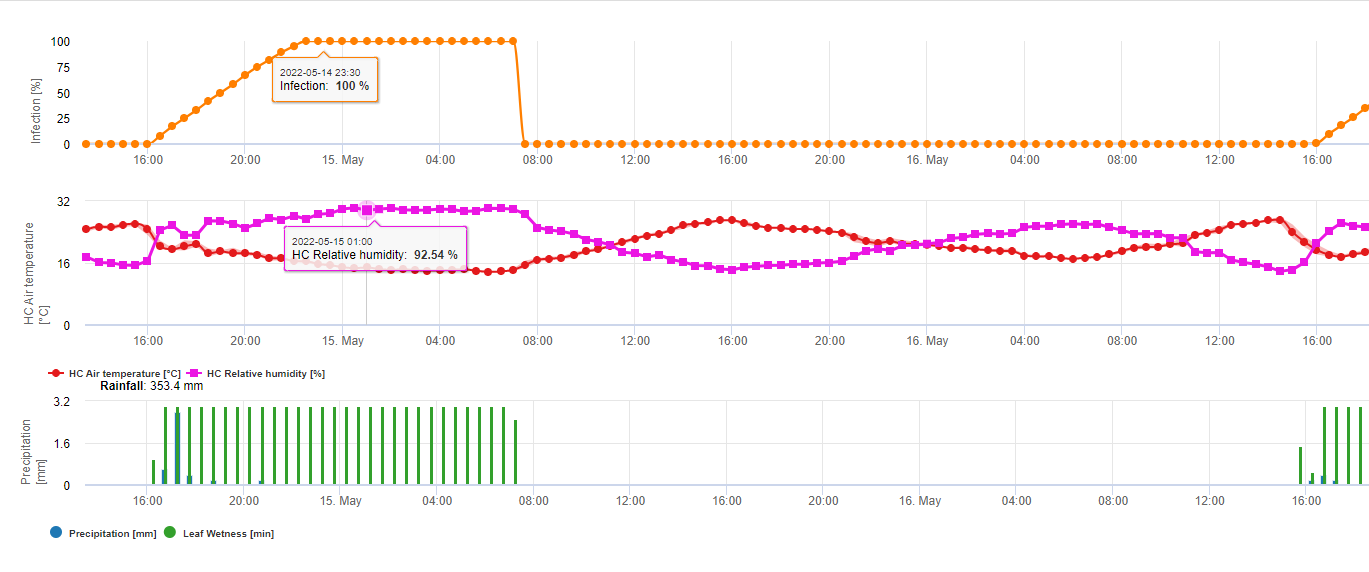
Підсумовуючи всі різні комбінації температур і вологості, які ми знайшли в літературі, ми вирішили вказати на інфекцію фузаріозної сажки, якщо температура і період вологості листя або періоди з відносною вологістю понад 85% перевищують значення, наведені на наступному графіку.
Інфекції починають розвиватися при дощі з опадами 2 мм. Зараження фузаріозом можна вважати, якщо показник розвитку інфекції досягає 100%. Розрахунок цього показника розвитку відбувається за співвідношенням між тривалістю вологих умов і температурою, що показано на графіку вище.
Ця модель використовується для візуалізації днів інфікування та кліматичних умов під час клімату. Знання фермерів про стадію розвитку різних сортів пшениці дає можливість прийняти рішення про застосування лікувального спрею одразу після зараження.
Попередження про мікотоксин фузаріозу
Випробування на зараження фузаріозною сажкою з подовженим періодом вологості листя призводять до високого вмісту мікотоксинів. З цієї інформації випливає, що період намокання листя протягом 48 годин або більше на стадіях 61 і 69 призводить до високого ризику накопичення мікотоксинів. Досліди, проведені під час аналізу ДОН у комерційно вирощуваній пшениці, показали, що періоди вологості листя, достатньо тривалі для інфікування після первинної інфекції на стадії 61-69, також можуть підвищити значення ДОН. У випадку більш тривалих періодів вологості листя мікотоксини можуть збільшуватися до стадії 85. FieldClimate накопичує показник ризику, пропорційний прогресу інфекції, для кожного успішного періоду інфікування протягом періоду, який був обраний для цих розрахунків. 6 щойно завершених періодів інфікування призведуть до ризику 100%. Зазвичай період вологості листя, що призводить до фузаріозної інфекції, довший, ніж мінімально необхідний. Тому більшість фузаріозних інфекцій призводять до збільшення ризику більш ніж на 17%. Значення ризику, що вказує на проблемну ситуацію з мікотоксинами, залежить від історії поля. Пшениця, вирощена після необробленої кукурудзи або необроблена пшениця після необробленої кукурудзи, може нести лише невеликий ризик, якщо вона не була обприскана в оптимальній ситуації. У необприсканій пшениці слід очікувати підвищених значень DON вже після 35% ризику. Пшениця після необробленої пшениці після будь-якої іншої культури, крім кукурудзи або пшениці, може нести вищий ризик 50%. Якщо ми маємо пшеницю після кукурудзи або пшениці з обробітком ґрунту, ризик може бути збільшений до 70%. Пшениця першого року повинна бути протестована на DON, якщо ризик перевищує 100%.
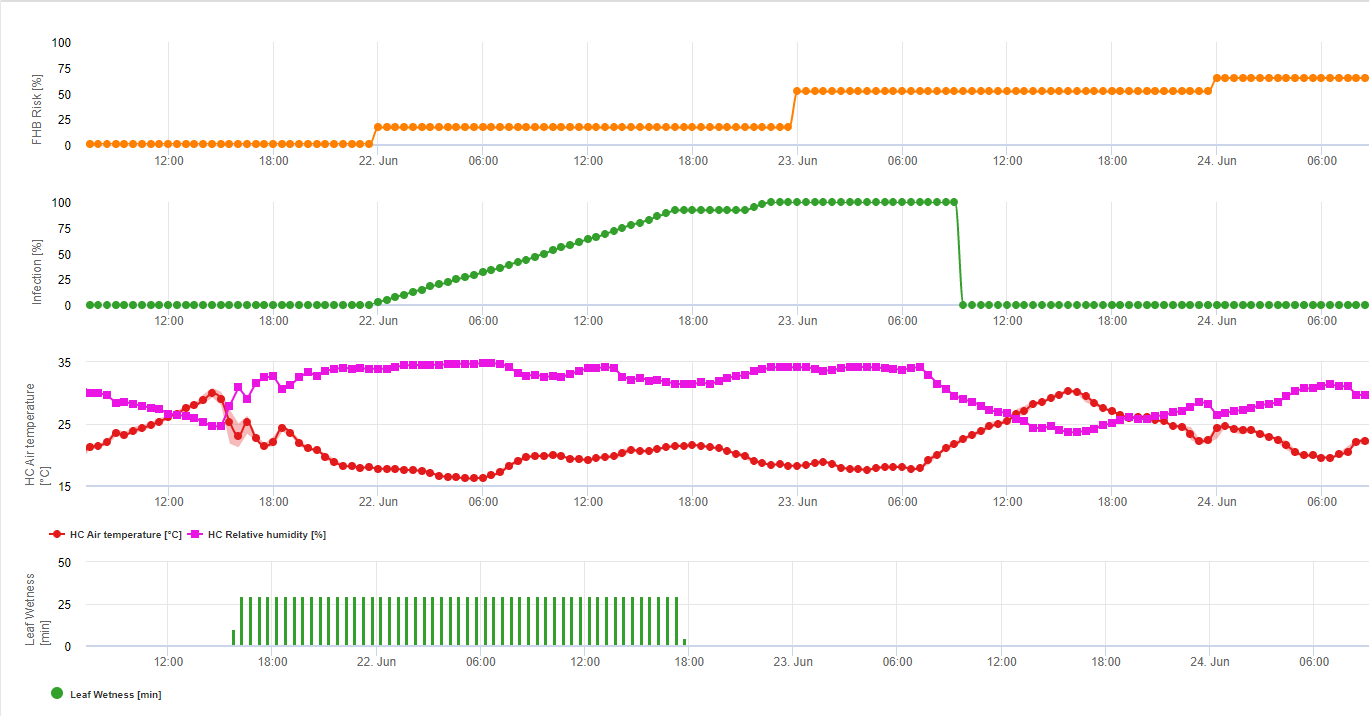
1) Модель ризику фузаріозної сажки який показує ризиковані періоди часу для інфікування. При досягненні інфікування 100% (зелена лінія) ризик (синя лінія) є дуже високим і умови для грибка є сприятливими для інфікування. Залежно від методу застосування (лікувальний, профілактичний) ризикований період часу показаний синьою лінією.
2) Фузаріозна сажка: У цій моделі інфікування FHB розраховується за допомогою опадів (необхідно 2 мм), відносної вологості (вище 85%) або вологості листя, температури під час процесу. Якщо інфікованість досягає 100%, то оптимальні умови для розвитку грибкового патогену досягнуті. Далі модель розраховує ризик мікотоксину FHB.
Існує дві основні септоріозні хвороби пшениці. Це септоріозна плямистість, спричинена грибком Septoria tritici Septoria tritici (телеоморф: Mycophaerella graminicola), та Септорія вузлувата (Septoria nodorum) плямистості, спричиненої грибком Septoria nodorum (телеоморф: Leptosphaeria nodorum). Обидві хвороби спричиняють серйозні втрати врожаю, які коливаються від 31 до 53% (Eyal, 1981; Babadoost and Herbert, 1984; Polley and Thomas, 1991). У всьому світі уражено понад 50 млн га пшениці, яка переважно вирощується в районах з великою кількістю опадів. Протягом останніх 25 років ці хвороби зростали і стали основним обмежувальним фактором для виробництва пшениці в певних регіонах. Під час сильних епідемій зерна сприйнятливих сортів пшениці стають щуплими і не придатними для помелу. Епідемії септоріозної та нодорозної плямистостей пшениці пов'язані зі сприятливими погодними умовами (часті дощі та помірні температури), специфічними культуральними практиками, доступністю посівного матеріалу та наявністю сприйнятливих сортів пшениці (Eyal et al., 1987).
Septoria spp. Біологія
За Еріком де Вольфом, пляма Septoria Tritici, Університет штату Канзас, квітень 2008 року Septoria tritici плямистість, відома як крапчаста плямистість листя, спричинена грибком Septoria tritici. Вона поширена в усіх регіонах світу, де вирощують пшеницю, і є серйозною проблемою в багатьох регіонах. Найбільшої шкоди септоріозна плямистість завдає, коли хвороба вражає верхні листки і качани сприйнятливих сортів наприкінці сезону.
Симптоми
Septoria tritici Симптоми плямистості вперше з'являються восени. Початковими симптомами є невеликі жовті плями на листі. З віком ці ураження часто набувають світло-коричневого відтінку, а плодові тіла гриба можна помітити на остюках, вбудовані в уражені ділянки. Ураження мають неправильну форму і варіюються від еліптичних до довгих і вузьких (рис. 1). Ураження містять маленькі, круглі, чорні цятки, які є плодовими тілами гриба. Чорні плодові тіла схожі на зерна чорного перцю і зазвичай їх можна побачити без допомоги лупи. Хвороба починається з нижніх листків і поступово переходить на прапорцевий листок. Листові піхви також схильні до ураження. У вологі роки крапчаста плямистість листя може переходити на качани і викликати коричневі ураження на колоссях і остюках, відомі як плямистість колосся. З віком ці ураження часто набувають світло-коричневого кольору, а плодові тіла гриба часто можна побачити вбудованими в ураження на остюках.
Фаза плямистості може спричинити значні втрати врожаю, але взаємозв'язок між тяжкістю хвороби та втратами врожаю недостатньо вивчений. Septoria tritici плямистість можна сплутати з іншими хворобами листя, які мають дуже схожі симптоми: плямистістю та Stagonspora nodorum наприклад, плямистості. Зазвичай рослини уражуються більш ніж однією з цих хвороб листя, і для точного діагностування найбільш поширених хвороб може знадобитися лабораторне обстеження. Лабораторне дослідження майже завжди потрібне, щоб визначити причину появи плямистостей. Знання виду не є важливим для прийняття рішення щодо обприскування, оскільки всі три хвороби однаково реагують на фунгіциди. Однак знання того, які хвороби найбільш поширені, є важливою частиною селекції сортів, оскільки різні гени контролюють стійкість до хвороб.
Найнадійніший спосіб відрізнити плямистість Septoria tritici від інших захворювань - це наявність чорних плодових тіл гриба. Гриб, що спричиняє засмагу, не утворює такого типу репродуктивних структур. Однак у вологих умовах гриб, що викликає плямистість Stagonospora nodorum, утворює світло-коричневі плодові тіла. Крім різниці в кольорі, ці структури також менші, ніж ті, що утворює Septoria tritici.
Життєвий цикл
Septoria tritici виживає влітку на залишках попереднього врожаю пшениці та ініціює інфікування восени. Існують певні докази того, що гриб здатний виживати в асоціації з іншими рослинами-господарями та насінням пшениці. Ці джерела гриба, ймовірно, є найбільш важливими за відсутності пшеничних решток. Незалежно від сівозміни або практики управління пожнивними рештками, зазвичай є достатньо посівного матеріалу, щоб ініціювати осінню інфекцію. Septoria tritici плямистості сприяє прохолодна волога погода. Оптимальний температурний діапазон - від 16 до 21 °C, проте інфікування може відбуватися і взимку за температури до 5 °C. Для зараження потрібно, щоб листя було вологим щонайменше 6 годин, а для максимального зараження - до 48 годин. Після зараження грибу потрібно від 21 до 28 днів, щоб розвинути характерні чорні плодові тіла і виробити нове покоління спор. Спори, що утворюються в цих плодових тілах, виділяються у вигляді липкої маси і потребують дощу, щоб потрапити на верхні листки і качани.
Зараження через Septoria tritici
Пікнідіоспори S. tritici проростають у вільній воді з обох кінців спори або з інтеркалярних клітин (Weber, 1922). Проростання спор починається приблизно через 12 годин після контакту з листком. Зародкові трубки ростуть хаотично по поверхні листка. Вебер (1922) спостерігав лише пряме проникнення між клітинами епідермісу, але інші дослідники дійшли висновку, що проникнення як через відкриті, так і через закриті продихи є основним способом проникнення в хазяїна (Benedict, 1971; Cohen and Eyal, 1993; Hilu and Bever, 1957). Кема та ін. (1996) спостерігали лише продихове проникнення. Гіфи, що проростають через продихи, звужуються до діаметру близько 1 мкм, а потім стають ширшими після досягнення підпродихової порожнини.
Гіфи ростуть паралельно поверхні листка під клітинами епідермісу, потім через мезофіл до клітин нижнього епідермісу, але не в епідерміс. Гаусторії не утворюються, а ріст гіф обмежується клітинами склеренхіми навколо судинних пучків, за винятком випадків, коли гіфи дуже щільні. Судинні пучки не інвазуються. Гіфи ростуть міжклітинно вздовж клітинних стінок через мезофіл, розгалужуючись на перегородці або в середині клітини. Макроскопічні симптоми не з'являються протягом приблизно 9 днів, за винятком випадкових мертвих клітин, але клітини мезофілу швидко гинуть через 11 днів. Пікніди розвиваються в субтоматичних камерах. Гіфи рідко проростають у клітини хазяїна (Hilu and Bever, 1957; Kema et al, 1996; Weber, 1922).
Успішне зараження відбувається лише після щонайменше 20 годин високої вологості. Лише кілька коричневих плям розвиваються, якщо листя залишається вологим протягом 5-10 годин після осадження спор (Holmes and Colhoun, 1974) або до 24 годин (Kema et al., 1996). Відносини хазяїн-паразит є однаковими як на стійких, так і на сприйнятливих пшеницях. Проростання спор на поверхні листя однакове незалежно від сприйнятливості. Кількість успішних проникнень приблизно однакова, але ріст гіф відбувається швидше на сприйнятливих сортах, що призводить до більшого ураження. Гіфи поширюються 44 Сесія 2 - Б.М. Кунфер за межі некротичних ділянок у всіх сортів. Токсин може відігравати певну роль у патогенезі (Cohen and Eyal, 1993; Hilu and Bever, 1957). На противагу цьому, колонізація була значно знижена на стійкій лінії (Kema et al., 1996).
Стагоноспори (септоріоз) та збудники септоріозу зернових: Інфекційний процес
Б.М. Канфер, кафедра патології рослин, Університет Джорджії, Гріффін, штат Джорджія
Процес зараження найбільш інтенсивно вивчався для Stagonospora (Septoria) nodorum та Septoria tritici. Одне поглиблене дослідження на тему Septoria passerinii є доступною. Майже вся інформація стосується інфікування пікнідоспорами. Однак процес інфікування іншими споровими формами досить схожий. Представлена інформація здебільшого стосується інфікування листків за оптимальних умов. Деякі дослідження були проведені з інтактними проростками, тоді як інші - з відокремленими листками. Інфікування колеоптиля та проростка пшениці S. nodorum був детально описаний Бейкером (1971) і переглянутий Кунфером (1983). Хоча точних порівнянь не проводилося, схоже, що інфекційний процес має багато спільного в кожній системі хазяїн-паразит і є типовим для багатьох некротрофних патогенів. Інформація про фактори, що впливають на розвиток симптомів і вираженість хвороби, не включена, але була розглянута іншими авторами (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Включено короткий опис факторів, що впливають на тривалість життя спор на поверхні листка.
Роль циркуляції та виживання спор на поверхні листка Найбільш детальна інформація про функцію кільця, що оточує пікнідоспори, які виділяються з пікнідія, міститься у S. nodorum. Циррус - це гель, що складається з білкових і сахаридних сполук. Його склад і функції подібні до складу та функцій інших грибів Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths and Peverett, 1980). Основними функціями компонентів цирруса є захист пікнідіоспор від десикації та запобігання передчасному проростанню.
Циррус захищає пікнідоспори так, що деякі з них залишаються життєздатними щонайменше 28 днів (Fournet, 1969). Коли циррус розводили водою, якщо концентрація розчину цирруса була >20%, проростало менше 10% пікнідіоспор. При меншій концентрації компоненти забезпечують поживні речовини, які стимулюють проростання спор і подовження зародкових трубок. Довжина зародкових трубок збільшувалася до концентрації цирруса 15%, а потім помірно зменшувалася за вищих концентрацій (Harrower, 1976). Бреннан та ін. (Brennan et al., 1986) повідомили про більшу схожість у розведеній рідині цирруса. Компоненти вихору знижували схожість при відносній вологості 10-60%. Після того, як спори дисперговані, стимулюючий вплив циррусної рідини, ймовірно, є незначним (Гріффітс і Певеретт, 1980).
При відносній вологості 35-45% спори S. tritici в циррі залишалися життєздатними щонайменше 60 днів (Gough and Lee, 1985). Компоненти цирруса можуть діяти як інгібітори проростання спор, або ж високий осмотичний потенціал цирруса може перешкоджати проростанню. Пікнідіоспори S. nodorum не виживали протягом 24 годин при відносній вологості вище 80% при 20 C. Спори виживали два тижні або більше при відносній вологості <10% (Griffiths and Peverett, 1980). Коли рідина цирруса S. nodorum розбавляли водою, близько двох третин пікнідоспор втрачали життєздатність протягом 8 годин, а через 30 годин при денному світлі проростала лише 5%. Коли спори зберігалися в темряві, 40% залишалися життєздатними через 30 годин (Brennan et al., 1986).
Сухі конідії S. nodorumу затінку та під прямими сонячними променями виживали на відкритому повітрі щонайменше 56 годин (Fernandes and Hendrix, 1986a). Проростання пікнідоспор S. nodorum пригнічувалося безперервним УФ-В (280-320 нм), тоді як проростання S. tritici не було. Подовження зародкових трубок під впливом безперервного ультрафіолетового випромінювання пригнічувалося для обох грибів порівняно з темрявою (Rasanayagam et al., 1995).
Зараження Septoria nodorum
Процес проникнення в організм хазяїна і розвиток S. nodorum в листі було детально вивчено кількома дослідниками (Baker and Smith, 1978, Bird and Ride 1981, Karjalainen and Lounatmaa, 1986; Keon and Hargreaves, 1984; Straley, 1979; Weber, 1922). Пікнідіоспори, як правило, розташовуються в заглибленнях між двома епідермальними клітинами, і багато спроб проникнення в листки починаються саме там. Спори проростають на поверхні листка у відповідь на вільну вологу (Fernandes and Hendrix, 1986b). Вони починають проростати через 2-3 години після осадження, а через 8 годин проростання може досягати 90%. Проникнення в листя починається приблизно через 10 годин після осадження спор (Bird and Ride, 1981; Brönnimann et al., 1972; Holmes and Colhoun, 1974).
На початку проростання зародкова трубка оточена аморфним матеріалом, який прикріплюється до листка. Зародкові трубки, що ростуть з обох кінців спори та з інтеркалярних клітин, мають тенденцію рости вздовж заглиблень між клітинами і часто орієнтовані вздовж довгої осі листка (O'Reilly and Downes, 1986). Гіфи зі спор, що не знаходяться в заглибленнях, ростуть хаотично з рідкісним розгалуженням (Straley, 1979). Утворюється аппресорій з інфекційним кілочком, який проникає крізь кутикулу і периклінальну стінку епідермальних клітин безпосередньо в клітинний просвіт, що призводить до швидкої загибелі клітини.
У багатьох випадках проникнення спочатку відбувається субкутикулярно або бічний ріст гіфи всередині клітинної стінки, перш ніж вона проростає в цитоплазму (Bird and Ride, 1981; O'Reilly and Downes, 1986). Проникнення як через відкриті, так і через закриті продихи також відбувається і може бути швидшим, ніж пряме проникнення (Harrower, 1976; Jenkins, 1978; O'Reilly and Downes, 1986; Straley, 1979). Зародкові трубки розгалужуються на продихах і з'єднаннях клітин епідермісу. Проникнення зародкової трубки в продихи може відбуватися без утворення продиху. Іноді проникнення відбувається через трихоми (Straley, 1979). Очевидно, більшість спроб проникнення є невдалими, оскільки в клітинах на місці спроби проникнення утворюються щільні сосочки (Karjalainen and Lounatmaa, 1986; Bird and Ride, 1981).
Після проникнення клітини епідермісу швидко відмирають і здерев'яніють, а гіфи вростають у мезофіл. Клітини мезофілу деформуються, і здерев'янілий матеріал відкладається поза деякими клітинами, які потім руйнуються. Здерев'яніння відбувається до того, як гіфи досягнуть клітини. Процес однаковий у всіх сортів, але у стійких сортів розвивається повільніше. Гіфи ростуть міжклітинно між клітинами епідермісу, а потім у мезофіл. Після проникнення в мезофіл руйнування хлоропластів починається через 6-9 днів (Karjalainen and Lounatmaa, 1986).
Однак швидкість фотосинтезу починає знижуватися вже через добу після зараження і до появи симптомів (Krupinsky et al, 1973). Склеренхімна тканина навколо судинних пучків запобігає інфікуванню судинної тканини. Судинні пучки блокують поширення гіфів через мезофіл, за винятком випадків, коли тканина склеренхіми молода і не повністю сформована (Baker and Smith, 1978).
Stagonospora nodorum виділяє широкий спектр ферментів, що руйнують клітинну стінку, включаючи амілазу, пектинметилестеразу, полігалактуронази, ксиланази та целюлазу in vitro та під час зараження листя пшениці (Baker, 1969; Lehtinen, 1993; Magro, 1984). Інформація, пов'язана з деградацією клітинної стінки ферментами, узгоджується з гістологічними спостереженнями. Ці ферменти можуть діяти разом з токсинами. Чутливість до ферментів може бути пов'язана зі стійкістю та швидкістю колонізації грибів (Magro, 1984). Як і багато некротрофів, збудники септоріозу та стагоноспори продукують фітотоксичні сполуки in vitro. Погіршення стану і загибель клітин до початку росту гіфів у тканину мезофілу (Bird and Ride, 1981) узгоджується з продукуванням токсинів. Однак, остаточна роль токсинів в інфекційному процесі та їх зв'язок зі стійкістю хазяїна не встановлені (Bethenod et al, 1982; Bousquet et al, 1980; Essad and Bousquet, 1981; King et al, 1983). Відмінності в діапазоні господарів між адаптованими до пшениці та ячменю штамами S. nodorum може бути пов'язана з виробленням токсинів (Bousquet і Kollmann, 1998). Ініціація проростання спор і відсоток пророслих спор не залежать від сприйнятливості хазяїна (Bird and Ride, 1981; Morgan 1974; Straley, 1979; Straley and Scharen, 1979; Baker and Smith, 1978).
Bird і Ride (1981) повідомили, що розширення зародкових трубок на поверхні листя відбувалося повільніше на стійких, ніж на сприйнятливих сортах. Цей механізм, що проявляється щонайменше через 48 годин після осадження спор, вказує на стійкість до проникнення до подовження зародкових трубочок. На стійких сортах було менше успішних проникнень, і проникнення відбувалося повільніше на стійких сортах (Baker and Smith, 1978; Bird and Ride, 1981). Було запропоновано лігніфікацію для обмеження інфекції як у стійких, так і у сприйнятливих сортів, але інші фактори сповільнювали розвиток гриба на стійких лініях. У сприйнятливих лініях гіфи, що швидше ростуть, можуть уникати лігніфікації клітин хазяїна. Через чотири дні після інокуляції ячменю ізолятом пшеничного біотипу гриба S. nodorumгіфи проростали крізь кутикулу, а іноді і в зовнішні целюлозні шари клітинних стінок епідермісу. Товсті сосочки відкладалися під проникними гіфами, і клітини не проникали (Keon and Hargreaves, 1984).
Зараження Septoria passerinii
Грін і Діксон (Green and Dickson, 1957) представили детальний опис процесу зараження S. passerinii на ячмені. Процес зараження схожий на S. tritici. Як і для S. tritici, час, необхідний для проникнення в листя, значно довший, ніж для S. nodorum. Зародкові трубки розгалужуються і ростуть по поверхні листка хаотично, але іноді вздовж заглиблень між клітинами епідермісу. Проникнення в листки відбувається майже виключно через продихи. Проростаючі гіфи набухають, і якщо проникнення не вдається, гіфи продовжують витягуватися. Через 48 годин після осадження спор проникнення не відбувається. Через 72 години зародкові трубки потовщуються над продихами, ростуть між охоронними клітинами, на поверхнях додаткових клітин і проникають у підпродихові порожнини. Пряме проникнення між клітинами епідермісу спостерігається дуже рідко.
Проростання спор і проникнення в рослину є однаковими на стійких і сприйнятливих сортах. На стійких сортах спостерігається значно менше поширення гіф всередині листя, а сосочки спостерігаються на багатьох, але не на всіх клітинних стінках. Гіфи ростуть під епідермою від одного продиху до іншого, але не проникають між клітинами епідерми. Мезофіл колонізується, але гаусторії не утворюються. Після того, як клітини мезофілу некротизуються, клітини епідермісу руйнуються. Розвиток міцелію в листі рідкісний і зазвичай блокується судинними пучками. У молодих листках, якщо судинна оболонка менш розвинена, гіфи проходять між пучком і епідермісом. Пікніди утворюються в субтоматних порожнинах, переважно на верхній поверхні листка (Green and Dickson, 1957).
Фактори, що впливають на довговічність спор на листковій поверхні Серед збудників стагоноспорозу та септоріозу зернових культур достовірна інформація про інфекційний процес була отримана лише для S. nodorum, S. triticiі S. passerinii. Як і багато інших некротрофних патогенів, жодна з груп збудників не викликає гіперчутливої реакції. Істотна відмінність в інфекційному процесі між збудниками септоріозу та стагоноспори полягає в тому, що проростання і проникнення спор відбувається набагато швидше для збудників септоріозу S. nodorum ніж для S. tritici і S. passerinii. Це має значний вплив на епідеміологію захворювань.
Збудники септоріозу проникають у рослину переважно через продихи, тоді як S. nodorum проникає як безпосередньо, так і через продихи. S. nodorum швидко проникає і вбиває клітини епідермісу, але S. tritici та S. passerinii не вбивають клітини епідермісу, поки гіфи не проростуть через мезофіл листка і не почнеться швидкий некроз. Гістологічні дослідження росту гриба після проникнення в організм хазяїна збігаються з даними епідеміологічних досліджень резистентності хазяїна. Резистентність сповільнює швидкість колонізації хазяїна, але не має помітного впливу на процес розвитку ураження.
Механізми, що контролюють реакцію хазяїна, пов'язані з ферментами, токсинами або іншими метаболітами, що вивільняються патогенами під час інфікування, все ще залишаються незрозумілими. Існує мало інформації про інфікування аскоспорами. Процес зараження, ймовірно, дуже схожий на процес зараження пікнідоспорами. Аскоспори Phaeosphaeria nodorum проростають в широкому діапазоні температур, а їхні зародкові трубки проникають безпосередньо в листок. Однак, за даними Rapilly та ін. (1973), аскоспори, на відміну від пікнідоспор, не проростають у вільній воді.
Модель інфекції, спричиненої септоріозом
Зараження септоріозом можливе при низьких температурах, тоді як температура нижче 7°C може не призвести до інфікування протягом 2 днів. Оптимальна температура для розвитку хвороби досягається в діапазоні від 16 до 21°C. Зараження можливе в період високої відносної вологості повітря або вологості листя протягом 14 годин або довше. Щоб відповідати цим умовам, ми вирішили розділити моделі на слабкі, помірні та сильні інфекції. Слабкі інфекції можуть бути задані, якщо патоген може інфікувати тканину хазяїна. Це означає, що слабкі інфекції можуть мати місце, якщо температури мінімальні, а періоди зволоження листя мають критичну тривалість. Помірне інфікування відбувається за умов, коли більшість випробувань на інфекційність призводять до задовільних результатів, а сильне інфікування відбувається за умов, коли патоген має оптимальні умови для інфікування.
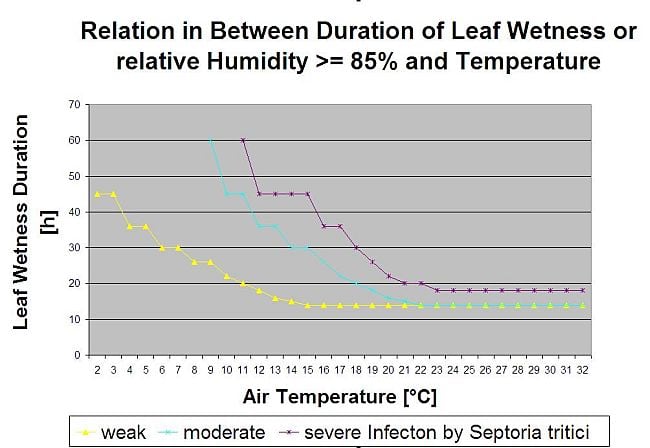

Починається інфікування після дощу 0,5 мм. Ми вирішили не використовувати модель утворення пікнід. Умовою, необхідною для утворення пікнід, вважається період з відносною вологістю повітря вищою за 85%. Очікується, що час життя пікнід становить 24 години. У всіх кліматичних зонах, де Septoria tritici має можливість інфікувати, ми знайдемо 2 години, що відповідають цим умовам, майже кожного дня на сході сонця.
Оцінка тяжкості інфекції
Для того, щоб мати змогу оцінити інфекційний тиск Septoria tritici між стадіями 10 (колеоптиль першого листкового жолоба) і 32 (другий вузол принаймні на 2 см вище вузла 1), а також між 32 і 51 (початок колосіння), ми повинні оцінити ступінь тяжкості інфекції, виходячи з кліматичних умов. Ця оцінка проводиться за шкалою від 1 до 5. Ступінь тяжкості 1 ставиться, якщо умови для слабкої інфекції виконані, а дощ випав менше 5 мм, в іншому випадку відповідний показник тяжкості буде дорівнювати 2. Ступінь важкості 3 присвоюється, якщо виконані умови для помірного зараження і випало менше 5 мм опадів. Якщо під час помірного зараження випало більше 5 мм опадів, а під час сильного зараження - менше 5 мм опадів, то ступінь вираженості інфекції дорівнює 4.
Сильна інфекція з кількістю опадів понад 5 мм відповідає значенню ступеня тяжкості 5.
Septoria tritici оцінка тиску хвороби
Клімат - це лише один з факторів, що впливає на розвиток хвороб на полях. Іншими двома факторами є історія поля та сприйнятливість вирощуваного сорту. Якщо ми можемо накопичити значення тяжкості хвороби від стадії 10 до стадії 32 до значення 4, ми можемо очікувати слабкого тиску хвороби з боку клімату. Якщо це значення досягає 6, ми можемо очікувати помірного тиску хвороб, а якщо воно досягає 10, ми можемо очікувати високого тиску хвороб з боку клімату. Знаючи сприйнятливість сорту та історію поля, ми зможемо прийняти рішення про те, обприскувати чи не обприскувати за слабкого або помірного тиску хвороб у цій ситуації. Накопичене значення 10 може призвести до обприскування на стадії 32 в будь-якому випадку. Рішення про обприскування на більш пізніх стадіях більше залежить від весняного клімату. Якщо ми зможемо накопичити значення ураженості, починаючи зі стадії 10, до значення 6, ми можемо очікувати слабкого тиску хвороби. Якщо це значення досягає 10, ми можемо очікувати помірного тиску хвороби, а якщо це значення досягає 15, ми можемо очікувати високого тиску хвороби, що залежить від кліматичної ситуації.
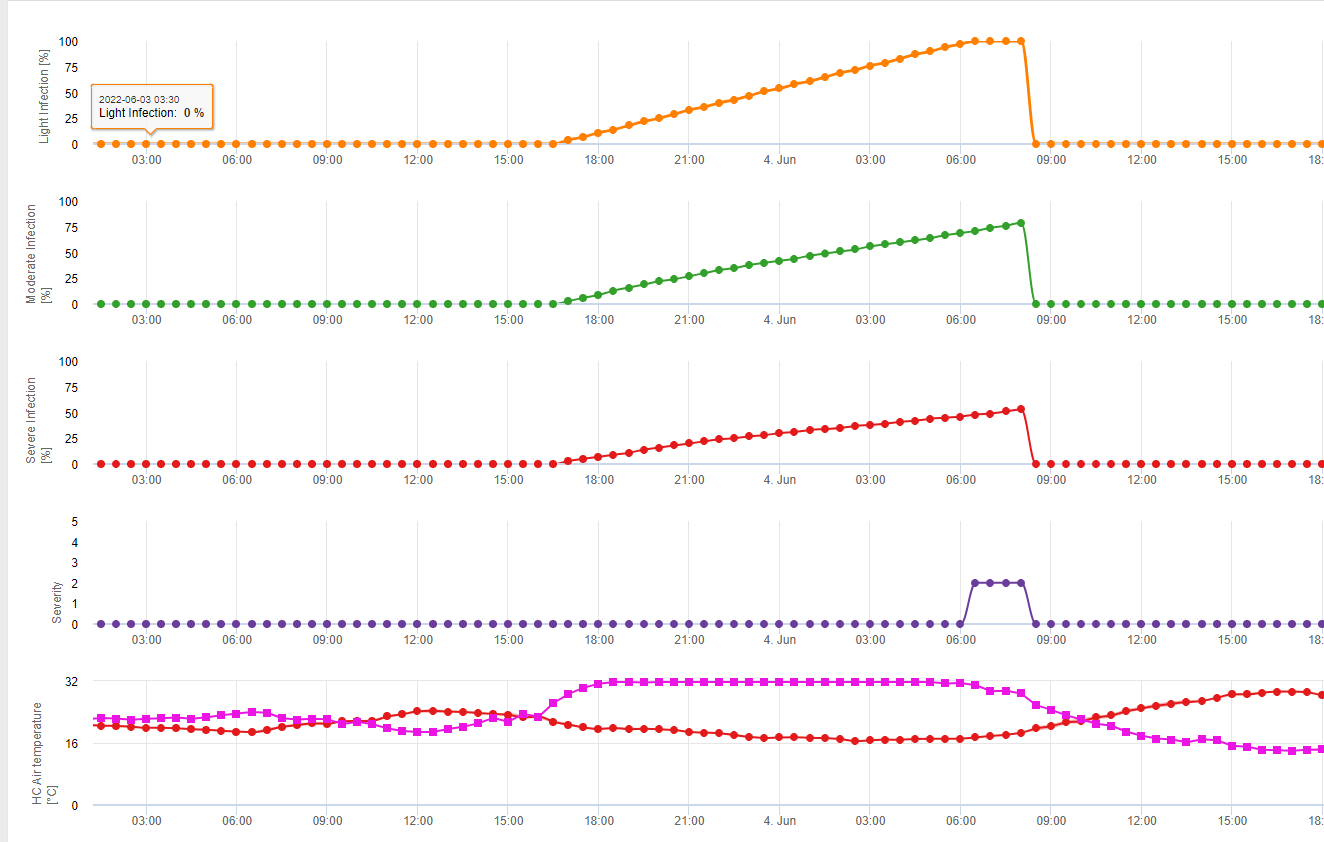
У FieldClimate ми показуємо Septoria tritici Тяжкість разом з трьома різними ступенями тяжкості інфекції на одному графіку (див. вище). Надмірна кількість опадів і тривалий період вологості листя створюють умови для розвитку важкої інфекції через S. tritici були повними 14 та 16 травня. Рівні тяжкості досягли найвищого значення 5 14 травня, що означає високий ризик інфікування.
Біологія інфекції Stagonospora nodorum дещо відрізняється від біології S. tritici, але ця різниця не є достатньо великою для окремої моделі. Тому ми пропонуємо використовувати цю модель для всього комплексу стагноспорозних і септоріозних хвороб зернових культур, включаючи S. passerinii. S. tritici і S. passerinii як правило, потребують більш тривалого періоду зволоження листя, ніж S. nodorum. У районах з високим тиском S. nodorum інфекції, класифіковані як слабкі, що мають ступінь тяжкості 2, слід лікувати більш серйозно, ніж в інших областях.
Для Септорія вузлувата (Septoria nodorum) модель ризику показана в FieldClimate (див. вище). Високий ризик був визначений 17 червня та 7 липня (100%). Залежно від сприйнятливої до інфекції стадії рослини, необхідно вжити заходів захисту рослин, якщо ризик досягне 80% (також див. прогноз погоди, захист рослин). Якщо ризик становить 100% і інфекція вже виявлена, для захисту рослин необхідно вжити системних заходів захисту рослин (лікувальне застосування).