Koşullar:
- Sabah güneş yükseldiğinde ve bağıl nem azaldığında, optimum sıcaklıklar 20°C ile 32°C arasındadır - iyi uçuş gösterilir.
- Sıcaklıklar optimum aralıkta değilse (çok soğuk/sıcak) veya çok ıslaksa (yaprak ıslaklığı) risk azalır.
- Çıktı günlük risktir.
Yani sabahları optimum sıcaklıklar ve düşen bağıl nem iyi bir uçuş gününe işaret eder. Gece boyunca ıslak olduğunda ve sıcaklıklar çok düşük olduğunda bu yayılma için kötüdür. Aynı şey gün boyunca sıcak ve nemli olduğunda da geçerlidir.
Külleme, birçok bitki türünde yaygın olarak görülen bir hastalıktır. Birkaç külleme mantarı farklı bitkilerde benzer hastalıklara neden olur (örneğin Podosphaera türleri elma ve sert çekirdekli meyvelerde; Sphaerotheca türleri çilek ve sert çekirdekli meyvelerde; Erysiphe necator üzüm asmalarında). Külleme mantarları genellikle nemli koşullar Kışlayan sporların serbest kalması ve bu sporların çimlenip bitki dokusunu enfekte etmesi için. Ancak mantarın bitkiyi enfekte ettikten sonra yerleşmesi ve büyümesi için neme ihtiyaç yoktur. Külleme normalde şunları tercih eder sıcakAkdeniz tipi iklimler.
Külleme çoğu bitkide kolayca fark edilebilir beyazdan griye değişen tozlu miselyum tarafından ve yaprakların, çiçeklerin, meyvelerin her iki tarafında ve sürgünlerde oluşan spor büyümesi. Ağaç meyvelerinde, enfeksiyonun meydana geldiği yerde kabuk üzerinde pürüzlü mantarımsı bir nokta gelişecektir.

Tüm külleme mantarları büyümek için canlı bitki dokusuna ihtiyaç duyar. Asma, ahududu ve meyve ağaçları gibi yaprak döken çok yıllık konukçularda külleme hayatta kalır bir sezondan diğerine enfekte tomurcuklar veya meyve veren gövdeler olarak kordonlar, dallar ve gövdelerin kabuklarında bulunan chasmothecia olarak adlandırılır.
Çoğu külleme mantarı, etkilenen bitki parçasının yüzeyinde ince miselyum tabakası olarak büyür. Birincil yayılma aracı olan sporlar, tozumsu büyümenin büyük kısmını oluşturur ve el merceği ile görülebilen zincirler halinde üretilir. Buna karşın, tüylü küf sporları küçük ağaçlara benzeyen dallı saplar üzerinde büyür. Ayrıca tüylü küf kolonileri beyaz yerine gridir ve çoğunlukla alt yaprak yüzeyinde görülür.
Külleme sporları aşağıdakiler tarafından taşınır rüzgar Konukçu bitkilere. Çimlenme için nem gereksinimleri farklılık gösterse de, birçok külleme türü su yokluğunda çimlenir ve enfekte olur. Aslında, bazı külleme mantarlarının sporları öldürülür ve çimlenme ve misel büyümesi bitki yüzeylerindeki su ile engellenir. Ilımlı sıcaklıklar ve gölge genellikle külleme gelişimi için en uygun koşullardır, çünkü sporlar ve miselyum aşırı sıcağa ve doğrudan güneş ışığına karşı hassastır.
Bu mantar tomurcuk pullarının içinde misel olarak kışı geçirir, birincil enfeksiyon bu enfekte tomurcuklardan yapraklar çıkarken meydana gelir. İkincil enfeksiyonlar, birincil ve sonraki ikincil enfeksiyonlar tarafından üretilen konidilerin üflenmesi veya yağmurla duyarlı dokulara sıçramasıyla meydana gelir. Meyve (çukur sertleşmeden önce) ve etli terminal büyüme enfeksiyona duyarlıdır.
S. pannosa için ortalama minimum, optimum ve maksimum sıcaklıklar yaklaşık 5°, 24° ve 24°C'dir. Tüm sıcaklıklarda nemli havaya kıyasla kuru havada çok daha fazla konidi oluşur (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli; 1954)
Külleme, kiraz küllemesi ile benzer bağıl nem ve sıcaklıklar altında yaygındır.
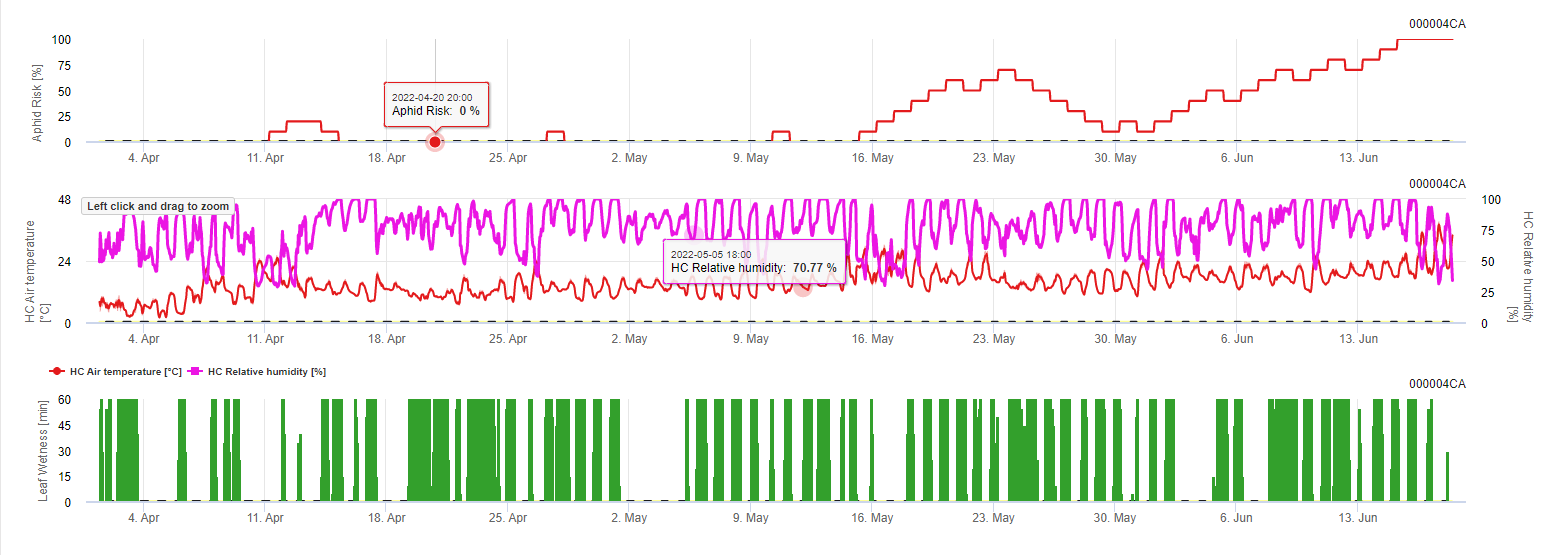
Saha İklim Modeli: Mantar hastalığı sıcaklık ve yaprak ıslaklık süresi faktörleri ile modellenmiştir. Örneğin 11 Mayıs'ta ılımlı sıcaklıklar altındaki yaprak ıslaklık süresi hastalığın gelişimini desteklemiş ve 100% riski belirlenebilmiştir.
Edebiyat:
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954): Külleme Hastalığının Sıcaklıkla İlişkisi. HILGARDIA. Kaliforniya Tarımsal Deney İstasyonu tarafından yayınlanan bir Tarım Bilimleri Dergisi. Kaliforniya Üniversitesi. Cilt 22/Sayı 17.
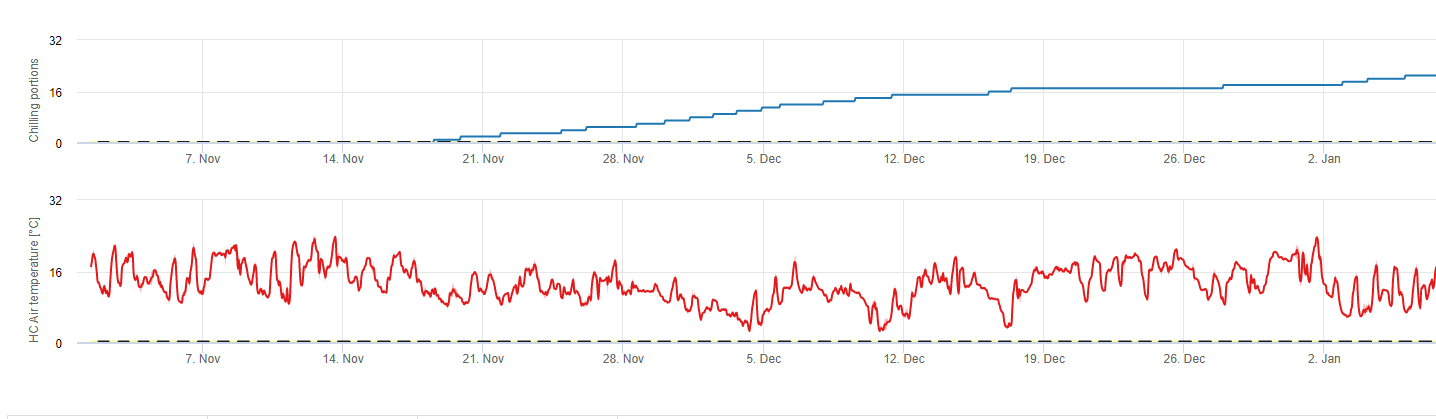
Chilling
Sert çekirdekli meyve ağaçları vejetatif ve meyve veren tomurcuklarını yazın geliştirir ve kış yaklaştıkça, zaten gelişmiş olan tomurcuklar hem daha kısa gün uzunluklarına hem de daha düşük sıcaklıklara tepki olarak uykuya geçer. Bu uyku hali ya da uyku aşaması, bu tomurcukları yaklaşan soğuk havadan korur. Tomurcuklar uyku haline girdikten sonra, donma noktasının çok altındaki sıcaklıklara karşı toleranslı olacak ve kış ortasındaki sıcak dönemlere tepki olarak büyümeyeceklerdir. Bu tomurcuklar yeterli soğuk hava birimi (CU) biriktirene kadar uykuda kalırlar. Ne zaman yeterince soğuk biriktiğinde, tomurcuklar ılık sıcaklıklara tepki olarak büyümeye hazırdır. Yeterli CU olduğu sürece çiçek ve yaprak tomurcukları normal şekilde gelişir. Eğer tomurcuklar kış aylarında yeterli soğutma sıcaklığı almazlar ağaçlar yetersiz soğuklama ile ilişkili fizyolojik semptomlardan bir veya daha fazlasını geliştirecektir: 1) yapraklanmanın gecikmesi, 2) meyve tutumunun azalması ve iliklenmenin artması ve 3) meyve kalitesinin düşmesi.
Yetersiz Üşüme Belirtileri
Gecikmiş Yapraklanma:
Yetersiz soğutmanın klasik bir belirtisi yapraklanmanın gecikmesidir. Bir ağaçta gövde uçlarına yakın küçük bir tutam yaprak olabilir ve uçların 12 ila 20 inç altında yaprak bulunmayabilir. Alt tomurcuklar sonunda kırılır ancak tam yapraklanma önemli ölçüde gecikir, meyve tutumu azalır ve ağaç zayıflar. Ayrıca, ağacın alt kısımlarından yoğun emilim yönetim sorunlarına neden olur ve gelecek yılın meyve tomurcuklarının normal gelişimi bozulabilir.
Azaltılmış Meyve Tutumu ve Düğmeleme:
Yetersiz soğuklamaya tepki olarak çiçeklenme, genellikle yaprak gelişiminde görülen modeli takip eder. Çiçeklenme gecikir, uzar ve pistil ve polen gelişimindeki anormallikler nedeniyle meyve tutumu azalır. Birçok şeftali çeşidinde çiçekler kabuk yarılmasından önce veya kabuk yarılması civarında dökülür, ancak 'Jersey Queen' ve 'Harvester' gibi diğerlerinde düğmeler oluşur. Düğmeler, görünüşte açan ama asla tam boy meyveye dönüşmeyen çiçeklerden kaynaklanır. Meyveler olgunlaştıkça küçük ve şekilsiz kalır. Eğer bu meyveleri kesip açarsanız, tohum ölür. Düğümlenme sezonun başlarında belirgin olmadığından, yetiştiriciler anormal meyveleri seyreltemez ve gelişen düğmeler böcekler ve hastalıklar için bir besin kaynağı ve kışlama alanı olarak hizmet eder.
Meyve Kalitesinde Azalma:
Yetersiz soğuklamanın meyve kalitesi üzerindeki etkileri muhtemelen en az tartışılan konudur ancak özellikle orta ve güney Teksas'ta çok yaygın olduğu görülmektedir. Yaprak büyümesi ve meyve tutumu üzerindeki etkiler dramatiktir ancak yetersiz soğuklamanın meyve kalitesi üzerindeki etkileri hafiftir ve diğer belirtiler görülmediğinde ortaya çıkabilir. Yetersiz soğuklama birçok çeşidin ucunun genişlemesine ve sertliğinin azalmasına neden olur. Ayrıca, meyve zemin rengi normalden daha yeşil olabilir, bunun nedeni muhtemelen zemin rengi yeşilden sarıya tam olarak değişmeden önce meyvenin sertliğini kaybetmesidir. Bu kalite sorunlarının kapsamı çeşide ve soğuklama eksikliğinin derecesine bağlıdır.
Modeller
Soğutmayı hesaplamak için kullanılan ve her biri soğutma biriminin ne olduğunu tanımlayan çeşitli modeller vardır. En yaygın üç model 45 derece F (7°C) altındaki saat sayısı modeli, 32 ile 45 derece F (2 ile 7°C) arasındaki saat sayısı modeli ve Utah modelidir. İlk iki model basittir ve bir soğutma birimini belirli sıcaklıkların altında veya arasında bir saat olarak tanımlar. Utah yöntemi daha karmaşıktır çünkü göreceli soğutma etkinliği ve negatif soğutma birikimi (veya soğutma olumsuzlaması) kavramlarını ortaya koymaktadır.
fieldClimate.com'da şunları kullanırız soğuk porsiyonların hesaplanması için model (CP). Soğutma birikimleri, 2 ila 7°C'lik bir sıcaklık aralığı kullanılarak soğutma kısımları olarak hesaplanır. Soğutma oranlarının hesaplanması 96 saatlik eşit veya daha fazla sürenin ardından >15°C' de sona erer (7 ila 15°C arasında tutar)
Hesaplamalar Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) Şeftali tomurcuklarında dinlenme tamamlanması için dinamik model çalışmasına dayanmaktadır. Acta Hortic 276: 165-174.