





Раннее поражение картофеля и томата
Рэндалл К. Роу, Салли А. Миллер, Ричард М. Ридель, Служба расширения Университета штата Огайо
Ранняя пятнистость - очень распространенное заболевание картофеля и томатов. Оно вызывает пятнистость листьев и поражение клубней у картофеля, а также пятнистость листьев, плодовую гниль и поражение стеблей у томата. Болезнь может развиваться в широком диапазоне климатических условий и при отсутствии контроля может быть очень разрушительной, часто приводя к полной дефолиации растений. В отличие от названия, болезнь редко развивается в ранние сроки, а обычно появляется на зрелой листве.
Симптомы
На листьях обеих культур первые симптомы обычно появляются на старых листьях и представляют собой мелкие, неправильной формы, темно-коричневые или черные мертвые пятна размером от точечки до 1/2 дюйма в диаметре. По мере увеличения пятен могут образовываться концентрические кольца в результате неравномерного роста организма в ткани листа. Это придает поражению характерный вид "пятна-мишени" или "бычьего глаза". Вокруг каждого пятна часто имеется узкий желтый ореол, а поражения обычно окаймлены жилками. Если пятен много, они могут разрастаться вместе, в результате чего зараженные листья желтеют и отмирают. Обычно сначала заражаются самые старые листья, которые засыхают и опадают с растения по мере продвижения болезни вверх по главному стеблю.
На томатах поражение стеблей может произойти в любом возрасте и привести к появлению небольших, темных, слегка впалых участков, которые увеличиваются и образуют круглые или удлиненные пятна с центрами более светлого цвета. На пораженных стеблях часто образуются концентрические пятна, похожие на пятна на листьях. Если для пересадки томатов используются зараженные семена, то вскоре после появления всходов рассада может засохнуть. Если на стеблях пересаженных растений или рассады появляются крупные поражения у линии почвы, растения могут оказаться опоясанными, что называется "воротничковой гнилью". Такие растения могут погибнуть при высадке в поле или, если стебли ослаблены, могут переломиться в начале сезона. Некоторые растения могут выжить с уменьшенной корневой системой, если части стеблей, расположенные над раковой опухолью, развивают корни там, где они соприкасаются с почвой. Однако такие растения обычно дают мало плодов или не дают их вовсе. Поражение стеблей гораздо менее распространено и разрушительно для картофеля.
Опадание цветков и пятнистость плодоножек, а также гибель молодых плодов могут произойти, когда раннее заболевание поражает томаты на стадии цветения. На более старых плодах ранняя пятнистость вызывает темные, кожистые впалые пятна, обычно в месте прикрепления плодоножки. Эти пятна могут увеличиваться и охватывать всю верхнюю часть плода, часто с концентрическими отметками, как на листьях. Пораженные участки могут быть покрыты бархатистыми черными массами спор. Плоды также могут быть заражены в зеленой или спелой стадии через трещины роста и другие раны. Зараженные плоды часто опадают, не достигнув зрелости.
На клубнях картофеля раннее поражение приводит к появлению поверхностных поражений, которые выглядят немного темнее, чем прилегающая здоровая кожица. Поражения обычно слегка впалые, круглые или неправильной формы, их размер варьируется до 3/4 дюйма в диаметре. Между здоровой и пораженной тканью обычно имеется четко очерченная и иногда слегка приподнятая граница. Внутри ткани видна коричневая или черная пробковая сухая гниль, обычно не более 1/4-3/8 дюйма глубиной. В старых поражениях могут образовываться глубокие трещины. В условиях Огайо заражение клубней встречается редко.
Патоген
Ранняя пятнистость вызывается грибком Alternaria solani, который выживает в инфицированных тканях листьев или стеблей на почве или в почве. Этот грибок повсеместно присутствует на полях, где выращивались эти культуры. Он также может переноситься на семенах томатов и в клубнях картофеля. Споры образуются на зараженных растительных остатках на поверхности почвы или на активных поражениях в довольно широком диапазоне температур, особенно при чередовании влажных и сухих условий. Они легко переносятся воздушными потоками, ветром, брызгами дождя и поливной водой. Заражение восприимчивых тканей листьев или стеблей происходит в теплую, влажную погоду с обильными росами или дождями. Раннее поражение может развиваться довольно быстро в середине и конце сезона и проявляется сильнее, когда растения испытывают стресс из-за плохого питания, засухи или других вредителей. Заражение клубней картофеля происходит через естественные отверстия на кожице или через повреждения. Клубни могут контактировать со спорами во время уборки, и поражения могут продолжать развиваться во время хранения.
TOMCAST (Джим Ясински, координатор TOMCAST в штатах Огайо, Индиана и Мичиган)
Справочная информация
TOMCAST (TOMato disease foreCASTing) - это компьютерная модель, основанная на полевых данных, которая пытается предсказать развитие грибковых заболеваний, а именно ранней пятнистости, септориозной листовой пятнистости и антракноза на томатах. Полевые регистраторы данных регистрируют ежечасные данные о влажности листьев и температуре. Эти данные анализируются в течение 24 часов и могут привести к следующим результатам формирование значения тяжести заболевания (DSV); по сути, это приращение развития болезни. По мере накопления DSV давление болезни на культуру продолжает нарастать. Когда количество накопленных DSV превышает интервал опрыскивания, рекомендуется применение фунгицида для ослабления давления болезни.
ТОМКАСТ
Сроки применения фунгицидов против ранней пятнистости, листовой пятнистости Septoria и антракноза
Система прогнозирования заболеваний на основе погодных условий под названием TOMCAST, разработанная доктором Роном Питбладо в Риджтаунском колледже сельскохозяйственных технологий в Онтарио, Канада, может быть использована для своевременного применения фунгицидов против трех грибковых заболеваний: ранней пятнистости (вызываемой Alternaria solani), листовой пятнистости Septoria (вызываемой Septoria lycopersici) и антракноза плодов (вызываемого Colletotrichum coccodes). Если поздняя пятнистость присутствует в вашем или соседних округах, или имеются условия для перемещения спор в ваш регион, используйте систему прогнозирования поздней пятнистости Simcast для определения времени применения фунгицидов.
ВАЖНОЕ ПРЕДОСТЕРЕЖЕНИЕ:
TOMCAST не стоит применять на фермах, где уже были случаи бактериальных заболеваний. Если вы часто сталкиваетесь с проблемами бактериальной пятнистости, пятнистости или рака на томатах, вам не следует использовать ТОМКАСТ, так как рекомендуемые интервалы опрыскивания будут недостаточны для борьбы с бактериальными заболеваниями, если вы смешиваете медь с фунгицидами. Информацию о борьбе с бактериальными заболеваниями можно найти здесь: http://extension.psu.edu/plants/vegetable-fruit/news/2015/farming-like-you-expect-bacterial-diseases
TOMCAST использует данные о влажности листьев и температуре для расчета значений тяжести заболевания (DSV), как показано в таблице 1.
TOMCAST является производной от оригинальной модели F.A.S.T. (прогнозирование Alternaria solani на томатах), разработанной докторами Мадденом, Пеннипакером и Макнабом в Университете штата Пенсильвания (PSU). Модель PSU F.A.S.T. была далее модифицирована доктором Питбладо в Риджтаунском колледже в Онтарио в модель TOMCAST, используемую в Университете штата Огайо.
DSV Значение тяжести заболевания (DSV) - это единица измерения, присвоенная определенному приросту развития болезни (раннего поражения). Другими словами, DSV - это числовое представление того, насколько быстро или медленно болезнь (раннее поражение) развивается на томатном поле. DSV определяется двумя факторами: влажностью листьев и температурой в часы "влажности листьев". По мере увеличения количества часов влажности листьев и температуры DSV накапливается быстрее. См. таблицу оценки тяжести заболевания ниже.
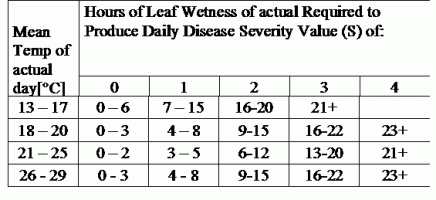
И наоборот, когда влажных часов для листьев меньше, а температура ниже, DSV накапливаются медленно, если вообще накапливаются. Когда общее количество накопленных DSV превышает установленный предел, называемый интервалом опрыскивания или порогом, рекомендуется опрыскивание фунгицидом для защиты листвы и плодов от развития болезни.

Интервал опрыскивания (определяющий время опрыскивания) может варьироваться в пределах 15-20 DSV. Точное количество DSV, которое должен использовать садовод, обычно предоставляется переработчиком и зависит от качества плодов. Интервал опрыскивания 15 DSV - это консервативное использование системы TOMCAST, то есть вы будете опрыскивать чаще, чем садовод, который использует интервал опрыскивания 19 DSV с системой TOMCAST. Компромисс заключается в количестве опрыскиваний, применяемых в течение сезона, и потенциальной разнице в качестве плодов.
ИСПОЛЬЗОВАНИЕ ТОМКАСТА
Томаты, выращенные в радиусе 10 миль от станции отчетности, должны воспользоваться функцией управления болезнями TOMCAST, которая помогает прогнозировать раннее заболевание, септориоз и антракноз. Если вы решите попробовать TOMCAST в этом сезоне, пожалуйста, помните о трех очень важных понятиях.
1) Если вы впервые используете систему, рекомендуется включить в программу только часть ваших площадей, чтобы понять, насколько она соответствует вашим стандартам качества и стилю работы.
2) Используйте ТОМКАСТ как руководство для более точного определения времени применения фунгицидов, понимая, что в некоторые сезоны вы можете применить больше препарата, чем того требует установленный график.
3) Чем дальше поле находится от места составления отчета, тем больше вероятность искажения накопления DSV, т.е. отчетное значение может быть на несколько DSV выше или ниже, чем в месте расположения поля. Это следует учитывать, когда применение фунгицидов возможно через несколько дней. Прослушайте сообщения о накоплении DSV на близлежащих станциях и сделайте триангуляцию к своему местоположению - это лучший способ приблизительно оценить накопление DSV.
ПЕРВЫЙ СПРЕЙ С ИСПОЛЬЗОВАНИЕМ TOMCAST
В течение многих лет велись дискуссии по поводу применения первого опрыскивания после ТОМКАСТа. Правило, изложенное в Руководстве по производству овощей 1997 года, касается даты посадки.
Растения томата, высаженные в поле до 20 мая, должны быть опрысканы в первый раз, когда DSV для этой зоны превысит 25 или когда наступит безопасная дата 15 июня. Безопасная дата используется только в том случае, если вы не проводили обработку с 20 мая, и является средством устранения начальной инокуляции болезни. После первого опрыскивания эти томаты в дальнейшем обрабатываются при превышении выбранного интервала опрыскивания (диапазон 15-20 DSV).
Томаты, высаженные после 20 мая, обрабатываются, когда они превышают выбранный интервал опрыскивания (диапазон 15-20 DSV) или когда они не были обработаны к дате отказа - 15 июня. Поэтому очень важно сравнить дату посадки томатов с датой начала регистрации DSV в этом районе, чтобы руководствоваться процессом принятия решения об опрыскивании).
Существует два основных заболевания пшеницы, вызываемых септориозом. Это пятнистость Septoria tritici, вызываемая грибком Septoria tritici (телеоморф: Mycophaerella graminicola), и Septoria nodorum пятнистость, вызываемая грибком Septoria nodorum (телеоморф: Leptosphaeria nodorum). Обе болезни вызывают серьезные потери урожая, которые, по имеющимся данным, составляют от 31 до 53 процентов (Eyal, 1981; Babadoost and Herbert, 1984; Polley and Thomas, 1991). Во всем мире поражено более 50 миллионов гектаров пшеницы, растущей в основном в районах с высоким уровнем осадков. За последние 25 лет эти болезни усилились и стали основным ограничивающим фактором производства пшеницы в некоторых районах. При сильных эпидемиях зерно восприимчивых сортов пшеницы сморщивается и становится непригодным для помола. Эпидемии пятнистости Septoria tritici и пятнистости Septoria nodorum на пшенице связаны с благоприятными погодными условиями (частые дожди и умеренные температуры), специфическими методами культуры, наличием инокулята и присутствием восприимчивых сортов пшеницы (Eyal et al., 1987).
Septoria spp. Биология
После Эрика де Вольфа, пятнистость Septoria Tritici, Университет штата Канзас, апрель 2008 г. Septoria tritici пятнистость, известная как пятнистость листьев, вызывается грибком Septoria tritici. Она распространена во всех пшеничных областях мира и представляет собой серьезную проблему во многих регионах. Пятнистость Septoria tritici наносит наибольший ущерб, когда болезнь поражает верхние листья и головки восприимчивых сортов в конце сезона.
Симптомы
Septoria tritici Симптомы пятнистости впервые проявляются осенью. Начальные симптомы - небольшие желтые пятна на листьях. По мере старения эти поражения часто становятся светло-коричневыми, а в поражениях на ости можно увидеть плодовые тела гриба. Поражения имеют неправильную форму, от эллиптической до длинной и узкой (рис. 1). Поражения содержат маленькие, круглые, черные крапинки, которые являются плодовыми телами гриба. Черные плодовые тела похожи на зерна черного перца и обычно видны без помощи лупы. Болезнь начинается на нижних листьях и постепенно переходит на флаговый лист. Листовые влагалища также подвержены поражению. Во влажные годы грибок пятнистости листьев может переходить на головки и вызывать коричневые поражения на ботве и ости, известные как пятнистость ботвы. Эти поражения часто становятся светло-коричневыми по мере старения, а плодовые тела гриба часто видны в поражениях на ости.
Фаза пятнистости ботвы может вызвать значительные потери урожая, однако взаимосвязь между тяжестью заболевания и потерями урожая изучена недостаточно хорошо. Septoria tritici пятнистость можно спутать с другими заболеваниями листьев, имеющими очень похожие симптомы: пятнистостью загара и Stagonspora nodorum например, пятнистость. Обычно растения поражаются более чем одной из этих болезней листьев, и для точного определения того, какие болезни наиболее распространены, может потребоваться лабораторное исследование. Лабораторные исследования почти всегда требуются для выявления причины пятнистости ботвы. Знание вида не имеет значения для принятия решения об опрыскивании, поскольку все три болезни одинаково реагируют на фунгициды. Однако знание того, какие болезни наиболее распространены, является важной частью селекции сортов, поскольку различные гены контролируют устойчивость к болезням.
Самый надежный способ отличить пятнистость Septoria tritici от других заболеваний - это наличие черных плодовых тел гриба. Грибок, вызывающий пятнистость, не производит такого типа репродуктивных структур. Однако во влажных условиях грибок, вызывающий пятнистость Stagonospora nodorum, образует светло-коричневые плодовые тела. Помимо разницы в цвете, эти структуры также меньше, чем те, которые производит Septoria tritici.
Жизненный цикл
Septoria tritici выживает в течение лета на остатках предыдущего урожая пшеницы и начинает заражение осенью. Есть некоторые свидетельства того, что гриб способен выживать в ассоциации с другими травянистыми хозяевами и семенами пшеницы. Эти источники грибка, вероятно, наиболее важны, когда остатки пшеницы отсутствуют. Независимо от севооборота или практики управления остатками, обычно имеется достаточно инокулята для инициирования осенних инфекций. Septoria tritici пятнистости благоприятствует прохладная, влажная погода. Оптимальный температурный диапазон - от 16 до 21 °C; однако заражение может происходить в зимние месяцы при температуре до 5 °C. Для заражения необходимо не менее 6 часов влажности листьев, а для максимального заражения требуется до 48 часов влажности. После заражения грибку требуется от 21 до 28 дней, чтобы сформировать характерные черные плодовые тела и произвести новое поколение спор. Споры, образующиеся в этих плодовых телах, выделяются в виде липкой массы, и для их разбрызгивания на верхние листья и головки требуется дождь.
Инфекция от Septoria tritici
Пикнидиоспоры S. tritici прорастают в свободной воде с обоих концов споры или из интеркалярных клеток (Weber, 1922). Прорастание спор начинается не ранее чем через 12 часов после контакта с листом. Зародышевые трубки растут беспорядочно на поверхности листа. Вебер (1922) наблюдал только прямое проникновение между эпидермальными клетками, но другие исследователи пришли к выводу, что проникновение через открытые и закрытые стоматы является основным способом проникновения в организм хозяина (Benedict, 1971; Cohen and Eyal, 1993; Hilu and Bever, 1957). Кема и др. (1996) наблюдали только стоматальное проникновение. Гифы, растущие через стоматы, сжимаются до диаметра около 1 мкм, затем становятся шире после достижения субстоматальной полости.
Гифы растут параллельно поверхности листа под эпидермальными клетками, затем через мезофилл к клеткам нижней части эпидермиса, но не в эпидермис. Гаустории не образуются, а рост гиф ограничен клетками склеренхимы вокруг сосудистых пучков, за исключением случаев, когда гифы очень плотные. Сосудистые пучки не инвазируются. Гифы растут межклеточно вдоль клеточных стенок через мезофилл, ветвясь на перегородке или в середине клетки. Макроскопические симптомы не проявляются в течение примерно 9 дней, за исключением отдельных мертвых клеток, но клетки мезофилла быстро погибают через 11 дней. Пикнидии развиваются в субстоматальных камерах. Гифы редко прорастают в клетки хозяина (Hilu and Bever, 1957; Kema et al, 1996; Weber, 1922).
Успешное заражение происходит только после как минимум 20 часов высокой влажности. Если листья оставались влажными в течение 5-10 часов после осаждения спор (Holmes and Colhoun, 1974) или до 24 часов (Kema et al., 1996), развивалось лишь несколько коричневых пятен. Отношения хозяин-паразит одинаковы на устойчивых и восприимчивых пшеницах. Прорастание спор на поверхности листьев одинаково независимо от восприимчивости. Количество успешных проникновений примерно одинаково, но рост гиф быстрее у восприимчивых сортов, что приводит к большему количеству поражений. Гифы распространяются 44 Заседание 2 - Б.М. Канфер за пределы некротической зоны у всех сортов. Токсин может играть определенную роль в патогенезе (Cohen and Eyal, 1993; Hilu and Bever, 1957). Напротив, колонизация была значительно снижена на устойчивой линии (Kema et al., 1996).
Стагоноспоры (септории) и возбудители септориоза зерновых культур: Процесс заражения
Б.М. Канфер, кафедра патологии растений, Университет Джорджии, Гриффин, штат Джорджия
Инфекционный процесс наиболее интенсивно изучался для Stagonospora (Septoria) nodorum и Septoria tritici. Одно углубленное исследование по Septoria passerinii имеется. Почти вся представленная информация относится к заражению пикнидиоспорами. Однако процесс заражения другими споровыми формами довольно схож. Представленная информация в основном относится к заражению листьев в оптимальных условиях. Некоторые исследования проводились с неповрежденными проростками, в то время как другие - с отделенными листьями. Заражение колеоптиля и проростков пшеницы спорами S. nodorum был подробно описан Бейкером (1971) и рассмотрен Канфером (1983). Хотя точных сравнений не проводилось, представляется, что инфекционный процесс имеет много общих черт в каждой системе хозяин-паразит и типичен для многих некротрофных патогенов. Информация о факторах, влияющих на развитие симптомов и проявление болезни, исключена, но была рассмотрена другими авторами (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Включена краткая информация о факторах, влияющих на продолжительность жизни спор на поверхности листьев.
Роль цирруса и выживаемость спор на поверхности листьев Наиболее подробная информация о функции цирруса, заключающего пикнидиоспоры, выделяющиеся из пикнидий, имеется для S. nodorum. Циррус представляет собой гель, состоящий из протеинатных и сахаридных соединений. Его состав и функции сходны с таковыми у других грибов из отряда Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths and Peverett, 1980). Основная роль компонентов цирруса - защита пикнидиоспор от высыхания и предотвращение преждевременного прорастания.
Циррус защищает пикнидиоспоры так, что некоторые из них остаются жизнеспособными по крайней мере 28 дней (Fournet, 1969). При разбавлении цирруса водой, если концентрация раствора цирруса составляла >20%, прорастало менее 10% пикнидиоспор. При более низкой концентрации компоненты обеспечивают питательные вещества, которые стимулируют прорастание спор и удлинение зародышевых трубок. Длина зародышевых трубок увеличивалась до 15% концентрации цирруса, затем умеренно снижалась при более высоких концентрациях (Harrower, 1976). Бреннан и др. (1986) сообщили о большей всхожести в разбавленной циррусовой жидкости. Компоненты цирруса снижали всхожесть при относительной влажности 10-60%. После рассеивания спор стимулирующее воздействие циррусовой жидкости, вероятно, незначительно (Griffiths and Peverett, 1980).
При относительной влажности 35-45% споры S. tritici в циррусе оставались жизнеспособными не менее 60 дней (Gough and Lee, 1985). Компоненты цирруса могут действовать как ингибитор прорастания спор, или высокий осмотический потенциал цирруса может препятствовать прорастанию. Пикнидиоспоры из S. nodorum не выживали в течение 24 часов при относительной влажности выше 80% при 20 C. Споры выживали две недели и более при относительной влажности <10% (Griffiths and Peverett, 1980). Когда циррусовая жидкость из S. nodorum разбавляли водой, около двух третей пикнидиоспор теряли жизнеспособность в течение 8 часов, а через 30 часов при дневном свете прорастали только 5%. Когда споры хранились в темноте, через 30 часов жизнеспособными оставались 40% (Brennan et al., 1986).
Сухие конидии S. nodorumв тени и под прямыми солнечными лучами, выживали на открытом воздухе не менее 56 часов (Fernandes and Hendrix, 1986a). Прорастание пикнидиоспор S. nodorum было подавлено непрерывным УФ-B (280-320 нм), тогда как прорастание S. tritici не было. Удлинение зародышевой трубки под непрерывным УФ-B было подавлено для обоих грибов, по сравнению с темнотой (Rasanayagam et al., 1995).
Инфекция, вызванная Septoria nodorum
Процесс проникновения в организм хозяина и развития S. nodorum внутри листа было подробно изучено несколькими исследователями (Baker and Smith, 1978, Bird and Ride 1981, Karjalainen and Lounatmaa, 1986; Keon and Hargreaves, 1984; Straley, 1979; Weber, 1922). Пикнидиоспоры обычно располагаются в углублениях между двумя эпидермальными клетками, и многие попытки проникновения в лист начинаются именно там. Споры прорастают на поверхности листа в ответ на свободную влагу (Fernandes and Hendrix, 1986b). Они начинают прорастать через 2-3 часа после осаждения, а через 8 часов прорастание может достигать 90%. Проникновение в листья начинается примерно через 10 часов после осаждения спор (Bird and Ride, 1981; Brönnimann et al., 1972; Holmes and Colhoun, 1974).
В начале прорастания зародышевая трубка окружена аморфным материалом, который прикрепляется к листу. Зародышевые трубки, растущие с обоих концов споры и из интеркалярных клеток, обычно растут вдоль углублений между клетками и часто ориентированы вдоль длинной оси листа (O'Reilly and Downes, 1986). Гифы из спор, не находящихся в углублениях, растут беспорядочно, иногда ветвясь (Straley, 1979). Аппрессорий образуется с инфекционным стержнем, который проникает через кутикулу и периклинальные стенки эпидермальных клеток непосредственно в просвет клетки, что приводит к быстрой гибели клеток.
Многие проникновения сначала являются субкутикулярными или боковой рост гифы происходит внутри клеточной стенки перед ростом в цитоплазму (Bird and Ride, 1981; O'Reilly and Downes, 1986). Проникновение через открытые и закрытые стоматы также происходит и может быть быстрее, чем прямое проникновение (Harrower, 1976; Jenkins, 1978; O'Reilly and Downes, 1986; Straley, 1979). Зародышевые трубки ветвятся на стоматах и стыках эпидермальных клеток. Проникновение зародышевой трубки в стомату может происходить без образования аппрессория. Иногда проникновение происходит через трихомы (Straley, 1979). По-видимому, большинство попыток проникновения не удается, при этом в клетках на месте попытки проникновения образуются плотные сосочки (Karjalainen and Lounatmaa, 1986; Bird and Ride, 1981).
После проникновения эпидермальные клетки быстро погибают и лигнифицируются, а гифы прорастают в мезофилл. Клетки мезофилла приобретают неправильную форму, а лигнифицированный материал откладывается снаружи некоторых клеток, которые затем разрушаются. Лигнификация происходит до того, как гифы достигают клетки. Этот процесс одинаков у всех сортов, но у устойчивых сортов развивается медленнее. Гифы растут межклеточно среди клеток эпидермиса, а затем проникают в мезофилл. Когда гифы проникают в мезофилл, через 6-9 дней начинается разрушение хлоропластов (Karjalainen and Lounatmaa, 1986).
Однако скорость фотосинтеза начинает снижаться в течение суток после заражения и до появления симптомов (Krupinsky et al, 1973). Ткань склеренхимы вокруг сосудистых пучков предотвращает заражение сосудистой ткани. Сосудистые пучки блокируют распространение гиф через мезофилл, за исключением тех случаев, когда ткань склеренхимы молодая и не полностью сформирована (Baker and Smith, 1978).
Stagonospora nodorum выделяет широкий спектр ферментов, разрушающих клеточную стенку, включая амилазу, пектинметилэстеразу, полигалактуроназы, ксиланазы и целлюлазу in vitro и при заражении листьев пшеницы (Baker, 1969; Lehtinen, 1993; Magro, 1984). Информация, касающаяся деградации клеточной стенки ферментами, согласуется с гистологическими наблюдениями. Эти ферменты могут действовать совместно с токсинами. Чувствительность к ферментам может быть связана с устойчивостью и скоростью колонизации грибами (Magro, 1984). Как и многие некротрофы, патогены Septoria и Stagonospora производят фитотоксичные соединения in vitro. Разрушение и гибель клеток до начала роста гиф в ткани мезофилла (Bird and Ride, 1981) согласуется с производством токсинов. Однако окончательная роль токсинов в инфекционном процессе и их связь с устойчивостью хозяина не установлена (Bethenod et al, 1982; Bousquet et al, 1980; Essad and Bousquet, 1981; King et al, 1983). Различия в ареале обитания между адаптированными к пшенице и ячменю штаммами S. nodorum может быть связано с производством токсинов (Bousquet and Kollmann, 1998). Начало прорастания спор и процент проросших спор не зависят от восприимчивости хозяина (Bird and Ride, 1981; Morgan 1974; Straley, 1979; Straley and Scharen, 1979; Baker and Smith, 1978).
Bird и Ride (1981) сообщили, что удлинение зародышевых трубочек на поверхности листьев происходило медленнее на устойчивых, чем на восприимчивых сортах. Этот механизм, проявляющийся по крайней мере через 48 часов после осаждения спор, указывает на предшествующую проникновению устойчивость к удлинению зародышевых трубок. На устойчивых сортах было меньше успешных проникновений, и проникновение происходило медленнее на устойчивых сортах (Baker and Smith, 1978; Bird and Ride, 1981). Было предложено, что лигнификация ограничивает инфекцию как на устойчивых, так и на восприимчивых сортах, но другие факторы замедляют развитие гриба на устойчивых линиях. В восприимчивых линиях более быстрорастущие гифы могут избежать лигнификации клеток хозяина.Через четыре дня после инокуляции ячменя пшеничным биотипом изолята S. nodorumгифы прорастали сквозь кутикулу и иногда в наружных целлюлозных слоях клеточных стенок эпидермиса. Под гифами проникновения откладывались толстые сосочки, которые не проникали в клетки (Keon and Hargreaves, 1984).
Инфекция, вызванная Septoria passerinii: Грин и Диксон (1957) представили подробное описание процесса заражения S. passerinii на ячмене. Процесс заражения похож на S. tritici. Как и S. tritici, для проникновения в листья требуется значительно больше времени, чем для S. nodorum. Зародышевые трубки ветвятся и растут по поверхности листа беспорядочно, но иногда вдоль углублений между эпидермальными клетками. Проникновение в лист происходит почти исключительно через стоматы. Гифы зародыша набухают, и если проникновение не удается, гифы продолжают удлиняться. Через 48 часов после отложения спор проникновение не происходит. Через 72 часа зародышевые трубки утолщаются над стоматами, прорастают между защитными клетками и на поверхностях аксессорных клеток, а также в субстоматальные полости. Прямое проникновение между эпидермальными клетками наблюдается крайне редко.
Прорастание спор и проникновение в организм хозяина одинаково на устойчивых и восприимчивых сортах. На устойчивых сортах гифы гораздо меньше разрастаются внутри листьев, а сосочки наблюдаются на многих, но не на всех клеточных стенках. Гифы растут под эпидермисом от одной стомы к другой, но не проникают между клетками эпидермиса. Мезофилл колонизируется, но гаустории не образуются. После того как клетки мезофилла становятся некротическими, клетки эпидермы разрушаются. Развитие мицелия в листе редкое и обычно блокируется сосудистыми пучками. В молодых листьях, если сосудистая оболочка менее развита, гифы проходят между пучком и эпидермисом. Пикнидии образуются в субстоматальных полостях, в основном на верхней поверхности листа (Green and Dickson, 1957).
Факторы, влияющие на продолжительность жизни спор на поверхности листьев Среди патогенов Stagonospora и Septoria зерновых культур окончательная информация о процессе заражения была получена только для S. nodorum, S. triticiи S. passerinii. Как и многие другие некротрофные патогены, обе группы патогенов вызывают реакцию гиперчувствительности. Существенное различие в инфекционном процессе между патогенами Septoria и Stagonospora заключается в том, что прорастание спор и их проникновение происходит гораздо быстрее для S. nodorum чем за S. tritici и S. passerinii. Это оказывает значительное влияние на эпидемиологию заболеваний.
Патогены Septoria проникают в растение в основном через стоматы, в то время как S. nodorum проникает как напрямую, так и через стоматы. S. nodorum быстро проникает и убивает эпидермальные клетки, но S. tritici и S. passerinii не убивают эпидермальные клетки до тех пор, пока гифы не прорастут через мезофилл листа и не начнется быстрый некроз. Гистологические исследования роста грибов после проникновения в организм хозяина соответствуют данным, полученным в результате эпидемиологических исследований устойчивости хозяина. Устойчивость замедляет скорость колонизации хозяина, но не оказывает заметного влияния на процесс развития поражения.
Механизмы, контролирующие реакцию хозяина, связанные с ферментами и токсинами или другими метаболитами, выделяемыми патогенами во время инфекции, до сих пор неясны. Существует мало информации о заражении аскоспорами. Вероятно, процесс заражения очень похож на процесс заражения пикнидиоспорами. Аскоспоры Phaeosphaeria nodorum прорастают в широком диапазоне температур, а их зародышевые трубки проникают непосредственно в лист. Однако, согласно Rapilly et al. (1973), аскоспоры, в отличие от пикнидиоспор, не прорастают в свободной воде.
Модель инфекции Septoria spp.
Инфекция септориозом возможна при низких температурах, в то время как при температуре ниже 7°C инфекция может не проявиться в течение 2 дней. Оптимальная температура для развития болезни достигается в области от 16 до 21°C. Инфекции возможны в период высокой относительной влажности или влажности листьев в течение 14 часов или дольше. Для соблюдения этих условий мы решили разделить на модели для слабых, умеренных и сильных инфекций. Слабые инфекции могут быть получены, если патоген способен инфицировать ткани хозяина. Это означает, что слабая инфекция может иметь место, если температура находится на минимальном уровне, а периоды увлажнения листьев имеют критическую продолжительность. Умеренное заражение будет происходить в условиях, когда большинство испытаний на зараженность приводят к приемлемым результатам, а сильное заражение происходит в условиях, когда патоген имеет оптимальные условия для заражения.

Заражение начинается после дождя в 0,5 мм. Мы решили не использовать модель для образования пикнидий. Условием, необходимым для образования пикнидий, предполагается период с относительной влажностью выше 85%. Время жизни пикнидий предположительно составляет 24 часа. Во всех климатических условиях, где Septoria tritici имеет шанс заразиться, мы найдем 2 часа, удовлетворяющие этим условиям, почти каждый день после восхода солнца.
Оценка тяжести инфекции: Чтобы иметь возможность оценить инфекционную нагрузку Septoria tritici в период между стадией 10 (первый лист в корешке колеоптиля) и стадией 32 (второй узел не менее чем на 2 см выше первого узла) и между 32 и 51 (начало колошения), мы должны оценить тяжесть инфекции в зависимости от климатических условий. Эта оценка производится по шкале от 1 до 5. Если условия для слабой инфекции выполнены и выпало менее 5 мм дождя, то ставится 1 балл, в противном случае соответствующий балл будет равен 2. Степень тяжести 3 присваивается, если условие умеренной инфекции выполнено и дождь прошел менее 5 мм. Если во время умеренной инфекции выпало более 5 мм дождя или менее 5 мм дождя во время тяжелой инфекции, присваивается степень тяжести 4.
Сильное заражение при выпадении более 5 мм осадков соответствует значению тяжести 5.
Septoria tritici оценка давления при заболевании: Климат является лишь одним из факторов, определяющих давление болезней на поле. Два других фактора - это история поля и восприимчивость выращиваемого сорта. Если мы можем накопить значения тяжести болезни от стадии 10 до стадии 32 до значения 4, мы можем ожидать слабое давление болезни со стороны климата. Если это значение достигнет 6, то можно ожидать умеренного давления болезни, а если оно достигнет 10, то можно ожидать сильного давления болезни со стороны климата. Зная восприимчивость сорта и историю поля, мы можем решить, опрыскивать или нет при слабом или умеренном давлении болезней в данной ситуации. Накопленное значение 10 может привести к опрыскиванию на стадии 32 в любом случае. Решение об опрыскивании на более поздней стадии в большей степени зависит от весеннего климата. Если нам удастся накопить значения тяжести с этапа 10 до значения 6, мы можем ожидать слабого давления болезни. Если это значение достигнет 10, мы можем ожидать умеренного давления болезни, а если это значение достигнет 15, мы можем ожидать высокого давления болезни в зависимости от климатической ситуации.
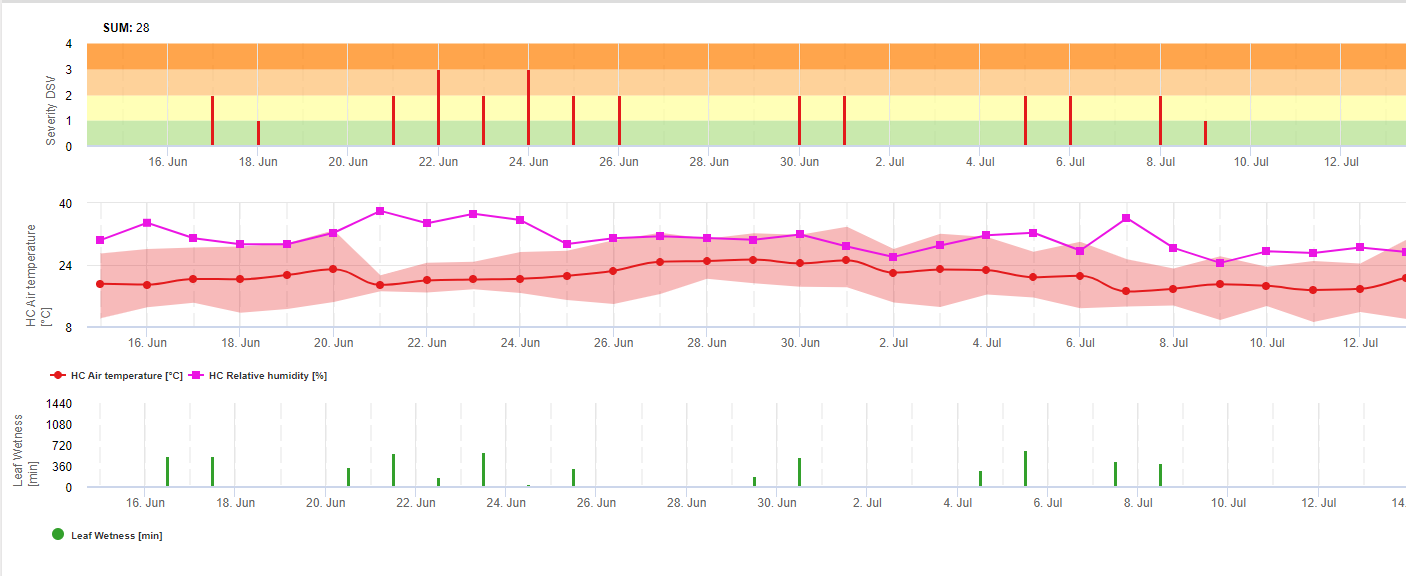
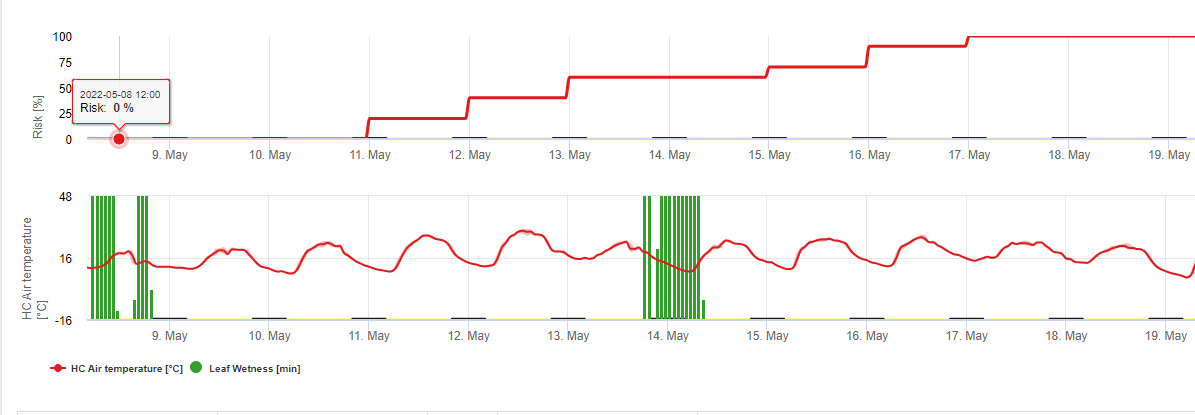
В FieldClimate мы показываем Septoria tritici Тяжесть вместе с тремя различными тяжестями инфекции на одном графике (см. выше). Из-за слишком сильных дождей и длительного периода увлажнения листьев создаются условия для тяжелой инфекции от S. tritici были полнопольными 14 и 16 мая. Уровни тяжести достигают наивысшего значения 5 14 мая, что означает высокий риск заражения.
Биология инфекции Stagonospora nodorum в некоторой степени отличается от таковой S. tritici, но эта разница недостаточно велика для создания отдельной модели. Поэтому мы предлагаем использовать эту модель для всего комплекса болезней Stagnospora и Septoria на зерновых культурах, включая S. passerinii. S. tritici и S. passerinii как правило, нуждаются в более длительных периодах увлажнения листьев, чем S. nodorum. В районах с высоким давлением S. nodorum инфекции, отнесенные к слабым и имеющие степень тяжести 2, должны рассматриваться более серьезно, чем в других областях.
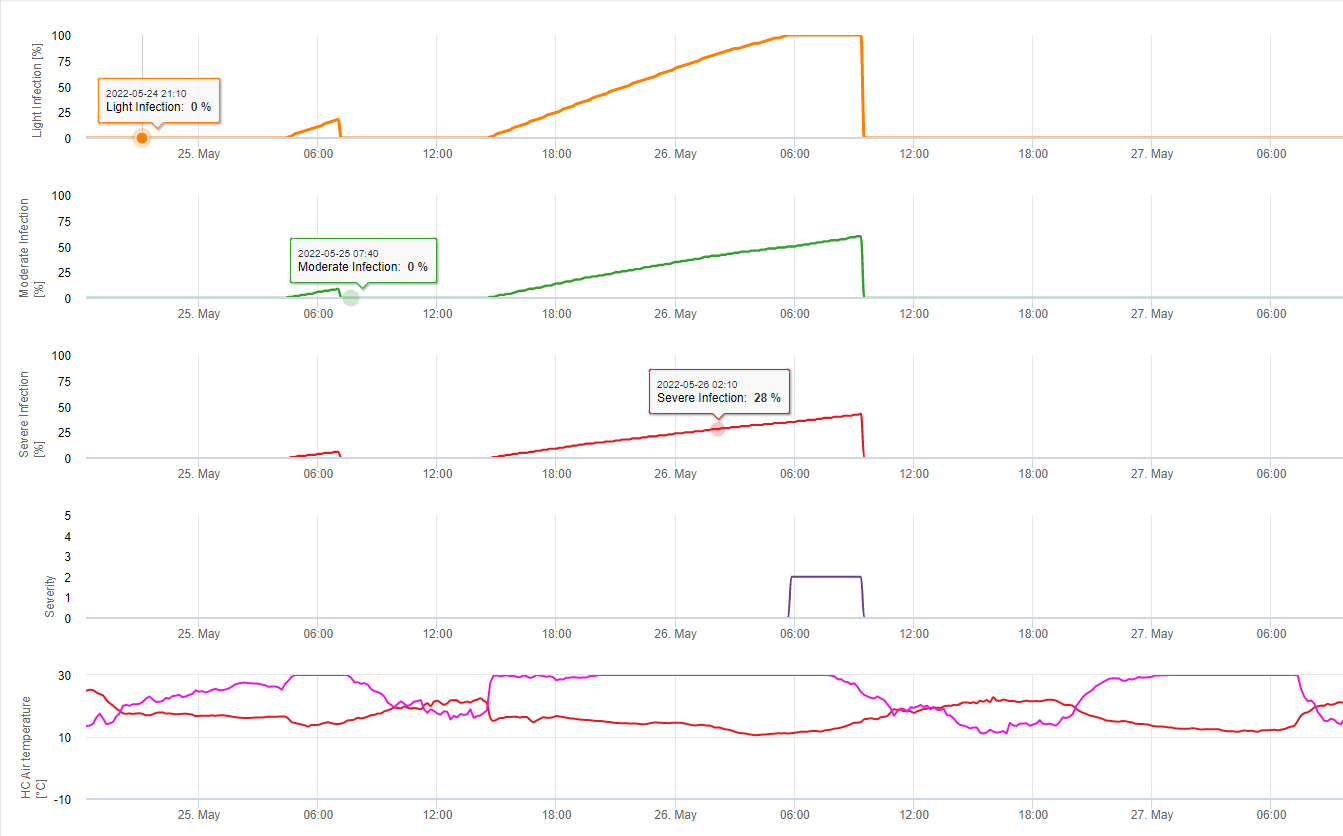
Для Septoria nodorum модель риска показана в FieldClimate (см. выше). Высокий риск был определен на 17 июня и 7 июля (100%). В зависимости от стадии восприимчивости растений к инфекции необходимо учесть меры защиты растений, если риск достигнет 80% (см. также прогноз погоды, защита растений). Если риск составляет 100% и инфекция уже установлена, то для защиты растений необходимо провести системные мероприятия по защите растений (лечебное применение).
Несколько видов патогенных для растений грибов из рода Colletotrichum вызывают антракноз у перцев и многих других овощей и фруктов. До конца 1990-х годов антракноз перцев и томатов ассоциировался только со спелыми или созревающими плодами. С тех пор появилась более агрессивная форма заболевания. Эта форма поражает перцы на любой стадии развития плодов и может угрожать рентабельности урожая перца в тех районах, где она развивается. Эта болезнь также может поражать томаты, клубнику и, возможно, другие плодовые и овощные культуры.
Симптомы
Круглые или угловатые впалые поражения развиваются на незрелых плодах любого размера. Часто на отдельных плодах образуются множественные поражения. При тяжелом течении болезни поражения могут сливаться. Часто розовые или оранжевые массы грибковых спор образуют концентрические кольца на поверхности поражений. В старых поражениях могут наблюдаться черные структуры, называемые ацервулами. С помощью ручной линзы они выглядят как маленькие черные точки; под микроскопом они похожи на пучки крошечных черных волосков. Патоген быстро и обильно образует споры и может быстро распространиться по всей культуре перца, что приводит к потере урожая до 100%. Поражения могут также появляться на стеблях и листьях в виде коричневых пятен неправильной формы с темно-коричневыми краями.
Патоген
Эта форма антракноза перца вызывается грибком Colletotrichum acutatum. Патоген выживает на растительных остатках зараженных культур и на других восприимчивых видах растений. В отсутствие зараженных растительных остатков гриб не переносится в почве в течение длительного времени. Грибок также может быть занесен в культуру с зараженными семенами. Во время теплые и влажные периоды, споры разбрызгиваются дождем или поливочной водой от больных плодов к здоровым. Заболевшие плоды служат источником инокулята, что позволяет болезни распространяться от растения к растению в пределах поля. Грибок выживает в семенах и на них. Антракноз заносится в поле с зараженными растениями при пересадке или может выживать в межсезонье в растительных остатках или на сорняках. Альтернативные хозяева включают сорняки и другие растения семейства пасленовых (томат, картофель, баклажан), хотя заражение этих хозяев крайне редко встречается во Флориде. Плоды заражаются, когда споры гриба или зараженный мусор попадают на растения перца. Новые споры образуются в зараженной ткани и затем распространяются на другие плоды. Рабочие также могут переносить споры с оборудованием или во время обработки зараженных растений. Заражение обычно происходит в теплую, влажную погоду. Температура около 80° F (27° C) являются оптимальными температурами для развития болезни, хотя заражение происходит как при более высоких, так и при более низких температурах. Серьезные потери происходят во время дождливой погоды, поскольку споры вымываются или разбрызгиваются на другие плоды, что приводит к увеличению числа заражений. Болезнь чаще развивается на зрелых плодах, которые долгое время находятся на растении, хотя она может развиваться как на незрелых, так и на зрелых плодах. Антракноз может поражать плоды при температуре от 15 °C до 30 °C. Но влажность длинных листьев необходимо для удовлетворения потребностей инфекции. При оптимальной температуре от 20°C до 25°C еще 12 часов после увлажнения листьев необходимы. Более высокая или холодная температура потребует еще более длительных периодов увлажнения листьев (для расчета не требуется линейная функция/массив). FieldClimate рассчитывает возможные события заражения на основе влажности листьев и температуры во время этого события.
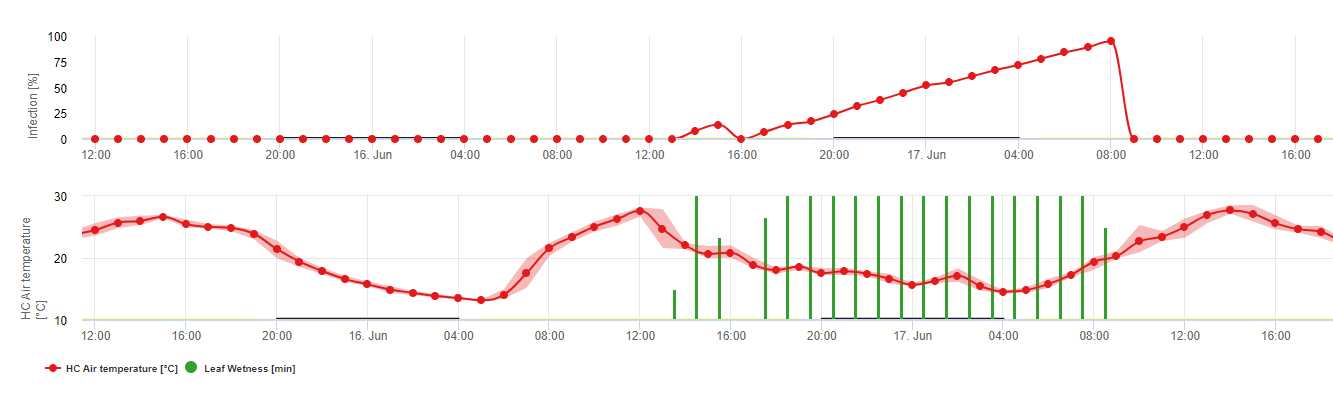
Биология Plasmopara viticola (Downy Mildew)
P. viticola является облигатным паразитом. Это означает, что для его роста необходимы зеленые, свежие органы лозы. В свободный от вегетации период он сохраняется, образуя плодовые тела, так называемые ооспоры. Ооспоры оомицетов могут очень долго сохраняться в почве. Поэтому мы можем обнаружить пушистую росу в местах, где заражение возможно не каждый год. Весной, когда верхний слой почвы достаточно влажный и теплый, ооспоры образуют так называемые макроспорангии, которые могут выпустить до 200 зооспор в свободную воду. Зооспоры переносятся ветром в капельках воды на листья и гроздья. Они имеют два жгутика и перемещаются в водной пленке на нижней стороне листьев, гроздей и молодых ягод, чтобы найти стому для проникновения в ткани растения. Они проникают и прорастают в стому, в которую переносят всю свою плазму менее чем за один час. При микроскопических исследованиях обнаружение стомы, энцистирование и прорастание в стому завершалось в течение 90 минут.
P. viticola растет в межклеточном пространстве и питается за счет гаусторий, проникающих в клетки эпидермиса и паренхимы. В зависимости от температуры и относительной влажности развивается достаточный межклеточный рост с достаточным количеством гаусторий, чтобы сформировать субстоматоловое тело, которое заполняет всю субстоматоловую область и поднимает эпидермальную ткань от паренхимальной ткани. Это приводит к появлению видимого симптома - масляного пятна.
Оомицеты являются споруляция в отсутствие света при относительной влажности воздуха очень высока. На сайте P. viticola споруляция отсутствует, если температура ниже 12°C и относительная влажность ниже 95%. Спорангиафоры образуются из везикул субстоматела и выходят из стомы. Свежие спорангии липкие и могут быть удалены из спорангиафоров только водой. При снижении относительной влажности спорангии становятся липкими и могут быть удалены ветром.
Спорангии выпускают до 20 зооспор в свободную воду. Чтобы попасть на свежие листья, эти зооспоры должны быть распространены ветром в капельках воды, или спорангии могут быть распространены дождем или ветром. Процесс заражения при первичной и вторичной инфекции одинаков.
Ввиду большого значения половой стадии для спячки патогена мы можем предположить, что типы спаривания подходят для всех виноградников, где встречается пуховая роса виноградной лозы. Образование зооспор происходит на старых листьях в течение лета и ранней осени.
Первичная инфекция в деталях
Инфекции, исходящие от ооспор, называются первичными инфекциями. Этот термин вводит в заблуждение, поскольку в начале лета может произойти несколько первичных инфекций. В эпидемиологических событиях первичная инфекция не играет важной роли, если на виноградниках достаточно маслянистых пятен и инфекционный потенциал летних спор (спорангиев) превышает потенциал ооспор.
Перезимовавшие ооспоры развиваются в так называемые макроспорангии при наличии достаточной относительной влажности в течение 24 часов. Макроспорангии выпускают свои зооспоры в свободную воду и под сильный дождь (как во время грозы). Споры переносятся на листья и/или зеленые побеги винограда. Первичные инфекции, таким образом, нуждаются в более продолжительном дожде периоды или несколько последовательных гроз. Первый дождь насыщает опавшие листья водой, а сильный дождь в последующие дни заставляет макроспорангии выпустить свои зооспоры, которые попадают на листья или побеги винограда. Для успешного заражения необходим достаточно длительный период увлажнения листьев, чтобы зооспоры достигли стомы листьев или побегов и заразили их (на рисунке выше показан цикл развития пушистой плесени виноградной лозы (Plasmopara viticola).
Модель для первичной инфекции Downy Mildew сначала проверяет, подходит ли погода для развития макроспорангиев. Это происходит при условии, что листья влажные или относительная влажность после дождя не опускается ниже 70%. В зависимости от температуры, макроспорангии могут развиваться в течение 16-24 часов. Если имеются зрелые макроспорангии, это будет показано графически на дисплее первичной инфекции Downy Mildew. Если макроспорангии присутствуют, сильный дождь может распространить их зооспоры. Непрерывный дождь в 5 мм считается сильным дождем и зооспоры распространяются, первичное заражение возможно, если увлажнение листьев длилось достаточно долго.
Вторичные инфекции
Вторичные инфекции Plasmopara viticola возможны только в том случае, если на вашем винограднике или поблизости от него уже имеются зрелые маслянистые пятна. Масляные пятна являются зрелыми, когда они способны спорулировать (производить спорангии). Спорангии образуются только в следующих случаях ночь. Солнечный свет подавляет споруляцию. Спорангии образуются, если теплее 12°C и относительная влажность очень высока. Скорость образования спорангиев увеличивается с ростом температуры вплоть до 24°C. Оптимальная температура для споруляции на европейских сортах винограда (Vitis vinifera) составляет около 24°C. Если температура превышает 29°C, споруляция не происходит. В нашей модели мы проверяем, если влажность более 95% происходит в течение ночи. Если это состояние длится в течение накопленной часовой температуры более 50°C, споруляция заканчивается, и новые спорангии из Plasmopara существуют в винограднике. Например, часы с температурой 50°C соответствуют 4 часам с температурой 13°C или 3 часам с температурой 17°C.
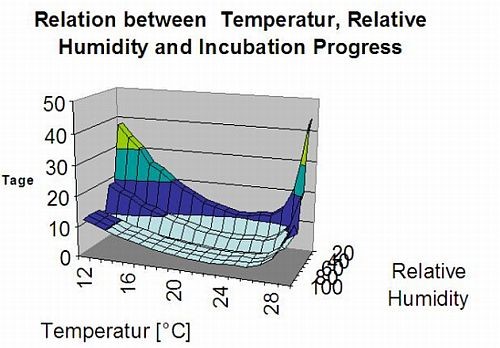
На следующем графике показана связь между температурой и влажными условиями (относительная влажность, влажность листьев, осадки), которые используются для моделирования инфекционных событий в fieldclimate.com.
Мучнистая роса томатов может вызываться тремя патогенами во всем мире.
Leveillula taurica (Oidiopsis taurica) является патогеном широкого спектра видов хозяев в теплом засушливом и полузасушливом климате Азии, Средиземноморья, Африки, а в последнее время и юго-запада США.
Erysiphe orontii (E. cichoracearum и E. polyphaga) - еще один вид, распространенный на многих растениях-хозяевах как в умеренных, так и в тропических регионах.
И как третий вид Odium lycopersicum.
Факторы развития заболевания
- уровень относительной влажности > 50% (оптимальная относительная влажность > 90%)
- свободная вода на поверхности листьев необязательна
- диапазон температур: 10-35 °C (лучше всего ниже 30 °C)
Мучнистая роса - это болезнь, управляемая инокулятом. Поэтому можно определить только рискованные периоды, основным фактором для повреждения, например, вспышки является начальный инокулят (который активен в широком диапазоне температур). Поэтому для стратегий контроля: объедините моделирование рискованного периода с мониторингом грибковой инокуляции (болезни) в поле!
Томат открытого грунта
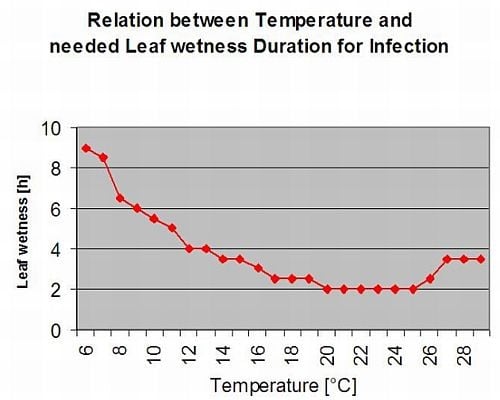
Ржавчина спаржи Puccinia asparagi благоприятствует влажный и теплый климат. Небольшие дожди и ветер распространяют уредиаспоры, в то время как сильные дожди могут смыть их в почву. По этой причине период увлажнения листьев с количеством осадков более 10 мм не принимается за период заражения. Периоды увлажнения листьев, вызванные небольшими дождями и росами продолжительностью от 3 до 8 часов при температуре от 25°C до 10°C, могут быть периодами заражения уредиаспорами летом и в конце лета.
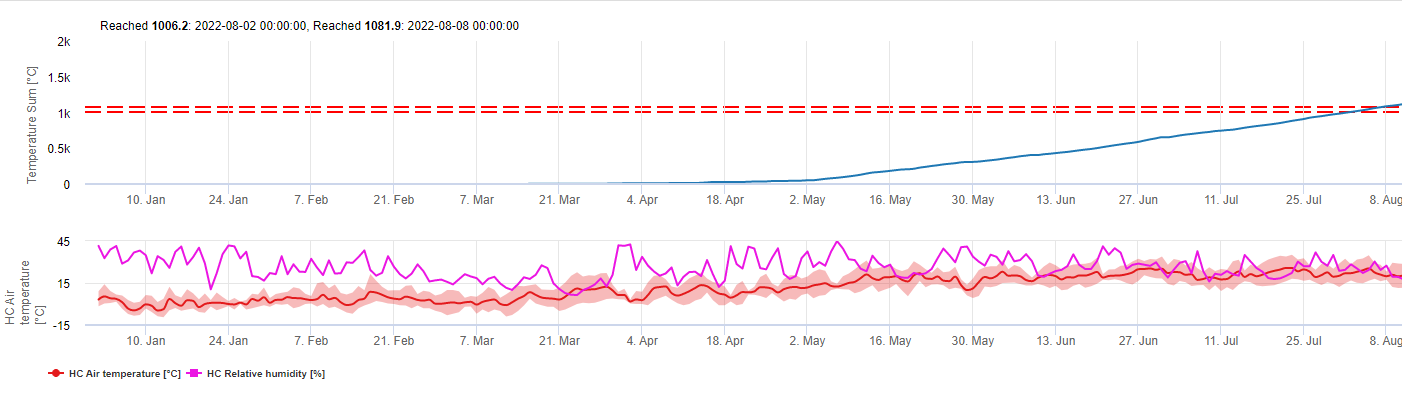
Эта модель оценивает дату первого появления C. beticola на основе накопленных среднесуточных температур с первого января выше 5°C. В периоды с относительной влажностью выше 60% принимается порог 1006.2°C, а если бы относительной влажности не было, то действует порог 1081.9°C.
Эта модель была разработана для Италии и используется в Италии и Германии.
Для 2010 года в Штирии CercoPrim указывает 19 июня для первого опрыскивания. Это примерно та же дата, которую модель риска Pessl Instruments или модель DIV указали бы для опрыскивания восприимчивых сортов.
Патоген
Болезнь пурпурной пятнистости на спаржевых копьях и папоротнике вызывается грибком Stemphylium vesicarium. Гриб выживает зимой в виде половых спор (аскоспор) в мешочке (аске), образующемся в зимующих структурах (псевдотециях), которые появляются в виде маленьких черных точек на остатках спаржи предыдущего сезона. Сайт аскоспоры высвобождаются из аска под воздействием дождя и могут переноситься ветром на только что появившиеся растения спаржи, где они вызывают первичную инфекцию вегетационного периода. Эти новые инфекции приводят к спорам (конидиям), образующимся бесполым путем, которые, в свою очередь, могут вызвать вторичные инфекции, и этот процесс повторяется до тех пор, пока температура и осадки благоприятны.
Болезнь
Появление пурпурной пятнистости как значительной проблемы при выращивании спаржи в Мичигане может быть связано с принятием системы культуры no- till, при которой высушенный папоротник предыдущего сезона измельчается в апреле и остается на поверхности почвы. Эти остатки сохраняются в течение сезона сбора урожая (середина июня) и видны в период роста папоротника (с конца июня по сентябрь) и являются источником аскоспор, которые вызывают инфекцию в начале вегетационного периода. Симптомы заболевания следующие: Болезнь проявляется в виде многочисленных, слегка впалых, пурпурных пятен с коричневым центром, возникающих на собранных копьецах и папоротнике. Поражения на копьях часто встречаются с наветренной стороны, так как песок наносит раны, что благоприятствует инфекции. В эпидемические годы пятнистость может наблюдаться на 60-90% копьях и может привести к отбраковке урожая, особенно для продажи на свежем рынке. Пятна также появляются на спаржевых папоротниках, поражая главный стебель, вторичные ветви и иглы (кладофиллы). Сильное заражение папоротника может привести к преждевременной дефолиации растения. Увеличение тяжести заболевания пурпурной пятнистостью связано с продолжительными периодами дождей, тумана или росы.
Текст предоставлен: Мэри К. Хаусбек, профессор и специалист по распространению, Мичиганский государственный университет, факультет патологии растений, г. Э. Лансинг
ТомКаст для спаржи
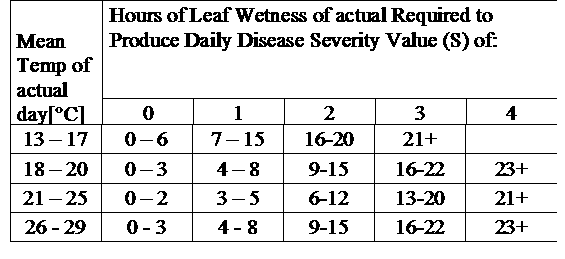
Общие сведения: TOMCAST (TOMato disease foreCASTing) - это компьютерная модель, основанная на полевых данных, которая пытается предсказать развитие грибковых заболеваний, а именно Ранняя пятнистость, септориозная пятнистость листьев и антракноз на томатах. Полевые регистраторы данных записывают ежечасные данные о влажности листьев и температуре. Эти данные анализируются в течение 24 часов и могут привести к формированию показателя тяжести заболевания (DSV); по сути, это прирост развития болезни. По мере накопления DSV давление болезни на урожай продолжает нарастать. Когда количество накопленных DSV превышает интервал опрыскивания, рекомендуется применение фунгицида для ослабления давления болезни.
TOMCAST является производной от оригинальной модели F.A.S.T. (прогнозирование Alternaria solani на томатах), разработанной докторами Мэдденом, Пеннипакером и Макнабом в Пенсильванском государственном университете (PSU). Модель PSU F.A.S.T. была далее модифицирована доктором Питбладо в Риджтаунском колледже в Онтарио в модель TOMCAST, которая в настоящее время используется службой расширения Университета штата Огайо.
DSV являются: Значение тяжести заболевания (DSV) - это единица измерения, присвоенная определенному приросту развития болезни (раннего поражения).
Другими словами, DSV - это числовое представление того, насколько быстро или медленно болезнь (раннее поражение) накапливается на томатном поле. DSV определяется двумя факторами: влажностью листьев и температурой в часы "влажности листьев". По мере увеличения количества часов влажности листьев и температуры DSV накапливается быстрее. См. таблицу оценки тяжести заболевания ниже.
И наоборот, когда влажных часов для листьев меньше, а температура ниже, DSV накапливаются медленно, если вообще накапливаются. Когда общее количество накопленных DSV превышает установленный предел, называемый интервалом опрыскивания или порогом, рекомендуется опрыскивание фунгицидом для защиты листвы и плодов от развития болезни.
Интервал опрыскивания (определяющий время опрыскивания) может варьироваться в пределах 15-20 DSV. Точное количество DSV, которое должен использовать садовод, обычно предоставляется переработчиком и зависит от качества плодов. Интервал опрыскивания 15 DSV - это консервативное использование системы TOMCAST, то есть вы будете опрыскивать чаще, чем садовод, который использует интервал опрыскивания 19 DSV с системой TOMCAST. Компромисс заключается в количестве опрыскиваний, применяемых в течение сезона, и потенциальной разнице в качестве плодов.
 ТомКаст со спаржей
ТомКаст со спаржей
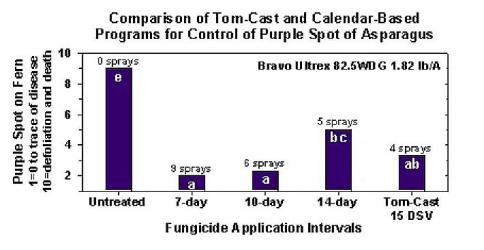
Исследования показали, что система прогнозирования заболеваний Tom-Cast является перспективной альтернативой опрыскиванию папоротника на коммерческих полях спаржи, основанному на календаре. Tom-Cast предупреждает садоводов, когда условия окружающей среды благоприятны для развития болезни пурпурной пятнистости (продолжительные периоды росы или дождей, сопровождаемые теплыми температурами). Эффективные фунгициды, применяемые в соответствии с прогнозом развития болезни Tom-Cast, позволяют садоводам справиться с пурпурной пятнистостью спаржи, сэкономив при этом деньги и сохранив окружающую среду.
Текст зачетный: Джим Ясински, координатор TOMCAST в штатах Огайо, Индиана и Мичиган
Модель заражения пурпурной пятнистостью
Сайт Stemphylium versicarium инфекционная модель основана на работе SUHERI и PRICE по луку и LLorent, VILARDELL, BUGIANI, GHERARDI и MONTESINO по груше.
Рассчитываются кривые заражения для легких, умеренных и тяжелых инфекций.
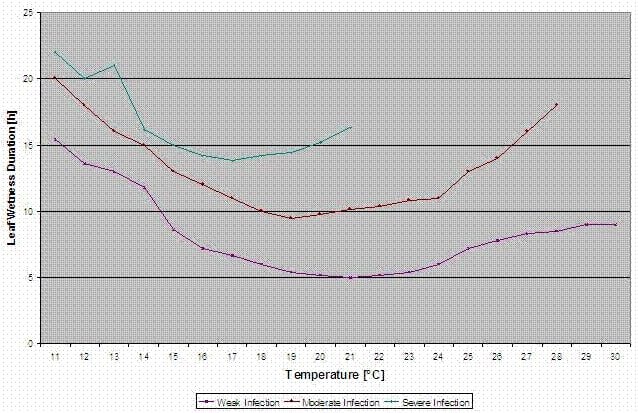
При таком разделении на благоприятные, умеренно благоприятные и очень благоприятные ситуации именно сельхозпроизводитель должен решить, насколько сильным будет давление болезней на конкретном поле и нужно ли ему покрывать конкретную инфекцию.
Условия:
Температура: 11-30°C
Влажность листьев > 0 (начало инфекции) или относительная влажность > 90% (только если инфекция уже была до расчета и все еще держится)
Легкая инфекция (11-30°C), умеренная инфекция (13-30°C), тяжелая инфекция (16-30°C), массив с продолжительностью увлажнения листьев (см. выше)
Сброс: если относительная влажность ниже 70%.