





Bacteria timpurie a cartofului și a tomatelor
Randall C. Rowe, Sally A. Miller, Richard M. Riedel, Ohio State University Extension Service
Bacteria timpurie este o boală foarte frecventă atât la cartof, cât și la tomate. Aceasta provoacă pete pe frunze și bubița tuberculilor la cartof, iar la tomate pete pe frunze, putregaiul fructelor și leziuni ale tulpinii. Boala poate apărea într-o gamă largă de condiții climatice și poate fi foarte distructivă dacă nu este controlată, ducând adesea la defolierea completă a plantelor. Spre deosebire de denumire, rareori se dezvoltă timpuriu, ci apare de obicei pe frunzele mature.
Simptomele
Pe frunzele ambelor culturi, primele simptome apar, de obicei, pe frunzele mai bătrâne și constau în pete moarte mici, neregulate, de culoare maro închis până la negru, cu diametrul de la un vârf de ac până la 1/2 inch. Pe măsură ce petele se măresc, se pot forma inele concentrice ca urmare a modelelor neregulate de creștere a organismului în țesutul frunzelor. Acest lucru conferă leziunii un aspect caracteristic de "punct-țintă" sau "ochi de taur". În jurul fiecărei pete există adesea un halo îngust, galben, iar leziunile sunt de obicei mărginite de nervuri. Atunci când petele sunt numeroase, ele pot crește împreună, ceea ce face ca frunzele infectate să se îngălbenească și să moară. De obicei, cele mai vechi frunze sunt infectate primele, iar acestea se usucă și cad de pe plantă pe măsură ce boala avansează pe tulpina principală.
La tomate, infecțiile tulpinii pot apărea la orice vârstă, rezultând în zone mici, întunecate, ușor înfundate, care se extind și formează pete circulare sau alungite cu centrul mai deschis la culoare. Pe leziunile de pe tulpină apar adesea marcaje concentrice, similare celor de pe frunze. În cazul în care semințele infestate sunt folosite pentru a începe transplanturile de tomate, răsadurile pot să se umezească la scurt timp după răsărire. Atunci când se dezvoltă leziuni mari la linia solului pe tulpinile transplanturilor sau răsadurilor, plantele pot fi încolțite, o afecțiune cunoscută sub numele de "putregaiul de guler". Astfel de plante pot muri atunci când sunt așezate în câmp sau, dacă tulpinile sunt slăbite, se pot rupe la începutul sezonului. Unele plante pot supraviețui cu un sistem radicular redus, dacă porțiuni de tulpini aflate deasupra cancerozității dezvoltă rădăcini acolo unde intră în contact cu solul. Cu toate acestea, astfel de plante produc de obicei puține sau deloc fructe. Leziunile tulpinii sunt mult mai puțin frecvente și mai puțin distructive la cartof.
În cazul în care tomatelor în faza de înflorire sunt atacate de arsura timpurie, pot apărea căderea florilor și pătarea tulpinilor fructelor, precum și pierderea fructelor tinere. Pe fructele mai bătrâne, arsura timpurie provoacă pete întunecate, cu aspect de piele, înfundate, de obicei în punctul de fixare a tulpinii. Aceste pete se pot mări până la a cuprinde întreaga porțiune superioară a fructului, prezentând deseori marcaje concentrice ca cele de pe frunze. Zonele afectate pot fi acoperite cu mase negre catifelate de spori. De asemenea, fructele pot fi infectate în stadiul verde sau copt prin fisuri de creștere și alte răni. Fructele infectate cad adesea înainte de a ajunge la maturitate.
Pe tuberculii de cartof, arsura timpurie are ca rezultat leziuni de suprafață care apar puțin mai întunecate decât pielea sănătoasă adiacentă. Leziunile sunt, de obicei, ușor înfundați, circulare sau neregulate și au un diametru de până la 3/4 inch. Există de obicei o margine bine definită și uneori ușor ridicată între țesutul sănătos și cel bolnav. La nivel intern, țesutul prezintă o putregai uscat, cornos, de culoare brună până la neagră, de obicei nu mai mult de 1/4 până la 3/8 de centimetru adâncime. În leziunile mai vechi se pot forma fisuri adânci. Infecția tuberculilor este neobișnuită în condițiile din Ohio.
Agenți patogeni
Ciuperca timpurie este cauzată de ciuperca Alternaria solani, care supraviețuiește în țesuturile infectate ale frunzelor sau tulpinilor, pe sau în sol. Această ciupercă este universal prezentă în câmpurile în care au fost cultivate aceste culturi. De asemenea, poate fi purtată pe semințele de tomate și în tuberculii de cartofi. Sporii se formează pe resturile de plante infestate la suprafața solului sau pe leziunile active într-un interval de temperatură destul de larg, în special în condiții de alternanță de umiditate și uscăciune. Aceștia sunt ușor de transportat de curenții de aer, de solul purtat de vânt, de stropii de ploaie și de apa de irigare. Infectarea țesuturilor sensibile ale frunzelor sau tulpinilor apare pe vreme caldă și umedă, cu rouă sau ploaie abundentă. Bacteria timpurie se poate dezvolta destul de rapid la mijlocul sau la sfârșitul sezonului și este mai gravă atunci când plantele sunt stresate de o nutriție deficitară, de secetă sau de alți dăunători. Infecția tuberculilor de cartof se produce prin deschideri naturale de pe piele sau prin răni. Tuberculii pot intra în contact cu sporii în timpul recoltării, iar leziunile pot continua să se dezvolte în timpul depozitării.
TOMCAST (Jim Jasinski, coordonator TOMCAST pentru OHIO, INDIANA și MICHIGAN)
Fond
TOMCAST (TOMato disease foreCASTing) este un model computerizat bazat pe date de teren care încearcă să prezică dezvoltarea bolilor fungice, și anume a mucegaiului timpuriu, a pătării frunzelor cu Septoria și a antracnozei la tomate. Aparatele de înregistrare a datelor amplasate pe teren înregistrează date orare privind umiditatea și temperatura frunzelor. Aceste date au fost analizate pe o perioadă de 24 de ore și pot duce la formarea unei valori de gravitate a bolii (DSV); în esență, este vorba de un progres al dezvoltării bolii. Pe măsură ce DSV se acumulează, presiunea bolii continuă să crească asupra culturii. Atunci când numărul de DSV acumulate depășește intervalul de pulverizare, se recomandă aplicarea unui fungicid pentru a atenua presiunea bolii.
TOMCAST
Calendarul de aplicare a fungicidelor pentru combaterea arsurii timpurii, a pătării frunzelor cu Septoria și a antracnozei
Un sistem de prognoză a bolilor bazat pe condițiile meteorologice, numit TOMCAST, dezvoltat de Dr. Ron Pitblado de la Colegiul de Tehnologie Agricolă Ridgetown din Ontario, Canada, poate fi utilizat pentru a planifica aplicarea fungicidelor pentru trei boli fungice: arsura timpurie (cauzată de Alternaria solani), pătarea frunzelor cu Septoria (cauzată de Septoria lycopersici) și antracnoza fructelor (cauzată de Colletotrichum coccodes). Dacă în județul dvs. sau în județele adiacente este prezentă apariția focarului târziu sau dacă există condiții pentru deplasarea sporilor în zona dvs., utilizați sistemul Simcast de prognoză a focarului târziu pentru a planifica aplicarea fungicidelor.
NOTĂ DE PRECAUȚIE IMPORTANTĂ:
TOMCAST nu este util în fermele care au un istoric de boli bacteriene. Dacă aveți în mod obișnuit probleme cu pete bacteriene, pătarea sau caniculă la roșiile dumneavoastră, nu ar trebui să utilizați TOMCAST, deoarece intervalele de pulverizare recomandate nu vor fi suficiente pentru controlul bolilor bacteriene dacă amestecați cupru în rezervor cu aplicațiile de fungicide. Găsiți aici informații despre reducerea bolilor bacteriene: http://extension.psu.edu/plants/vegetable-fruit/news/2015/farming-like-you-expect-bacterial-diseases
TOMCAST utilizează datele privind umiditatea și temperatura frunzelor pentru a calcula valorile de severitate a bolii (DSV), după cum se arată în tabelul 1.
TOMCAST este derivat din modelul original F.A.S.T. (Forecasting Alternaria solani on Tomatoes) dezvoltat de Dr. Madden, Pennypacker și MacNab ? la Pennsylvania State University (PSU). Modelul F.A.S.T. de la PSU a fost modificat ulterior de Dr. Pitblado de la Colegiul Ridgetown din Ontario în ceea ce recunoaștem acum ca fiind modelul TOMCAST utilizat de Ohio State University Extension.
DSV sunt Valoarea de gravitate a bolii (DSV) este unitatea de măsură dată unui anumit grad de dezvoltare a bolii (arsură timpurie). Cu alte cuvinte, o DSV este o reprezentare numerică a rapidității sau a lentorii cu care se acumulează boala (arsura timpurie) într-un câmp de tomate. DSV este determinat de doi factori: umiditatea frunzelor și temperatura în timpul orelor de "umezeală a frunzelor". Pe măsură ce numărul de ore de umezeală a frunzelor și temperatura crește, DSV se acumulează într-un ritm mai rapid. Consultați graficul de mai jos privind valoarea de severitate a bolilor.
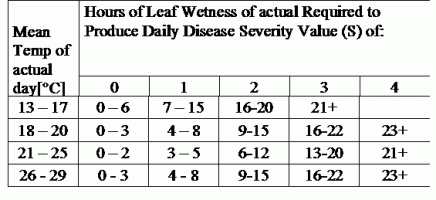
Dimpotrivă, atunci când sunt mai puține ore de umezeală a frunzelor și temperatura este mai scăzută, DSV se acumulează încet sau deloc. Atunci când numărul total de DSV acumulate depășește o limită prestabilită, numită interval de pulverizare sau prag, se recomandă o pulverizare cu fungicid pentru a proteja frunzele și fructele de dezvoltarea bolii.

Intervalul de stropire (care determină când trebuie să pulverizați) poate varia între 15-20 DSV. DSV-ul exact pe care ar trebui să îl folosească un cultivator este de obicei furnizat de către procesator și depinde de calitatea fructelor și de utilizarea finală a roșiilor. Urmarea unui interval de pulverizare de 15 DSV reprezintă o utilizare conservatoare a sistemului TOMCAST, ceea ce înseamnă că veți pulveriza mai des decât un cultivator care utilizează un interval de pulverizare de 19 DSV cu sistemul TOMCAST. Compromisul constă în numărul de stropiri aplicate în timpul sezonului și în potențialul de diferență în ceea ce privește calitatea fructelor.
UTILIZAREA TOMCAST
Roșiile cultivate pe o rază de 16 km de o stație de raportare ar trebui să beneficieze de funcția de gestionare a bolilor din TOMCAST pentru a ajuta la prognozarea atacului bacterian timpuriu, a septoriozei și a antracnozei. Dacă vă decideți să încercați TOMCAST în acest sezon, vă rugăm să țineți cont de trei concepte foarte importante.
Unu: Dacă este prima dată când utilizați acest sistem, se recomandă ca doar o parte din suprafața dumneavoastră să fie introdusă în program pentru a vedea cum se potrivește cu standardele dumneavoastră de calitate și cu stilul de operare.
Doi: Folosiți TOMCAST ca un ghid pentru a vă ajuta să planificați mai bine aplicarea fungicidelor, având în vedere că în unele sezoane este posibil să aplicați mai mult produs decât ar putea necesita un program stabilit.
Trei: Cu cât un câmp de tomate se află mai departe de un loc de raportare, cu atât crește probabilitatea de distorsiune în acumularea DSV, adică valoarea raportată poate fi cu câteva DSV mai mare sau mai mică decât cea înregistrată în locația câmpului. Acest lucru ar trebui luat în considerare atunci când aplicarea fungicidelor este probabil la câteva zile distanță. Ascultarea rapoartelor DSV de la stațiile din apropiere și triangularea la propria locație reprezintă cea mai bună modalitate de a estima aproximativ acumularea DSV.
PRIMA PULVERIZARE FOLOSIND TOMCAST
De-a lungul anilor au existat discuții cu privire la aplicarea primului spray atunci când se urmează TOMCAST. Regula enunțată în Ghidul de producție legumicolă din 1997 se concentrează în jurul datei de plantare.
La plantele de tomate care intră în câmp înainte de 20 mai, prima stropire ar trebui să se aplice atunci când DSV pentru zona respectivă depășește 25 sau atunci când se ajunge la data de 15 iunie. Data de siguranță este folosită numai dacă nu ați tratat după 20 mai și este un mijloc de a elimina inoculul inițial al bolii. După prima pulverizare, aceste roșii sunt tratate ulterior, atunci când se depășește intervalul de pulverizare ales (intervalul 15-20 DSV).
Roșiile plantate după 20 mai sunt tratate atunci când depășesc intervalul de pulverizare ales (intervalul 15-20 DSV) sau când nu au fost tratate până la data de 15 iunie, data limită de siguranță. Prin urmare, este esențial să se compare data plantării tomatelor cu data la care a început raportarea DSV în zona respectivă pentru a ghida procesul de decizie privind pulverizarea.
Există două boli majore de septorioză la grâu. Acestea sunt Septoria tritici blotch, provocată de ciuperca Septoria tritici (teleomorf: Mycophaerella graminicola) și Septoria nodorum pată, cauzată de ciuperca Septoria nodorum (teleomorfă: Leptosphaeria nodorum). Ambele boli cauzează pierderi serioase de randament care variază între 31 și 53% (Eyal, 1981; Babadoost și Herbert, 1984; Polley și Thomas, 1991). La nivel mondial, sunt afectate peste 50 de milioane de hectare de grâu, care cresc în principal în zonele cu precipitații abundente. În ultimii 25 de ani, aceste boli au fost în creștere și au devenit un factor limitativ major pentru producția de grâu în anumite zone. În cazul unor epidemii grave, miezul cultivarelor de grâu sensibile se zbârcește și nu este bun pentru măcinare. Epidemiile de Septoria tritici blotch și Septoria nodorum blotch la grâu sunt asociate cu condiții meteorologice favorabile (ploi frecvente și temperaturi moderate), practici culturale specifice, disponibilitatea inoculului și prezența unor soiuri de grâu sensibile (Eyal et al., 1987).
Septoria spp. Biologie
În urma lui Erick De Wolf, Septoria Tritici Blotch, Universitatea de Stat din Kansas, aprilie 2008 Septoria tritici cunoscută sub numele de "pată de frunze pătate", este cauzată de ciuperca Septoria tritici. Este răspândit în toate zonele de cultivare a grâului din lume și reprezintă o problemă gravă în multe regiuni. Septoria tritici este cel mai dăunătoare atunci când boala atacă frunzele superioare și căpățânile soiurilor sensibile la sfârșitul sezonului.
Simptomele
Septoria tritici Simptomele de pete apar pentru prima dată toamna. Simptomele inițiale sunt mici pete galbene pe frunze. Aceste leziuni devin adesea cafeniu deschis pe măsură ce îmbătrânesc, iar corpurile de fructificare ale ciupercii pot fi văzute încorporate în leziunile de pe coame. Leziunile au o formă neregulată și variază de la eliptică la lungă și îngustă (figura 1). Leziunile conțin pete mici, rotunde și negre, care reprezintă corpurile de fructificare ale ciupercii. Corpurile de fructificare negre arată ca niște boabe de piper negru și pot fi văzute, de obicei, fără ajutorul unei lupe. Boala începe pe frunzele inferioare și progresează treptat spre frunza steag. Tecile frunzelor sunt, de asemenea, susceptibile la atac. În anii umezi, ciuperca pătată a frunzelor se poate muta pe căpățâni și poate provoca leziuni brune pe glume și pe coame, cunoscute sub numele de pătarea glumelor. Aceste leziuni devin adesea cafeniu deschis pe măsură ce îmbătrânesc, iar corpurile de fructificare ale ciupercii sunt adesea vizibile încorporate în leziunile de pe coame.
Faza de pătare a glumei poate cauza pierderi semnificative de producție, dar relația dintre severitatea bolii și pierderea de producție nu este bine înțeleasă. Septoria tritici poate fi confundată cu alte boli ale frunzelor care au simptome foarte asemănătoare: pătarea bronzului și Stagonspora nodorum pată, de exemplu. Este obișnuit ca plantele să fie infectate de mai multe dintre aceste boli foliare și poate fi necesară o examinare de laborator pentru a diagnostica cu precizie care sunt cele mai răspândite boli. Examinarea de laborator este aproape întotdeauna necesară pentru a distinge cauza pătării glumei. Cunoașterea speciei nu este importantă pentru deciziile de pulverizare, deoarece toate cele trei boli răspund în mod similar la fungicide. Cu toate acestea, cunoașterea bolilor care sunt cele mai răspândite este o parte importantă a selecției soiurilor, deoarece diferite gene controlează rezistența la boli.
Cel mai sigur mod de a distinge petele de Septoria tritici de celelalte boli este prezența fructificațiilor negre ale ciupercii. Ciuperca care provoacă pătarea bronzată nu produce acest tip de structură reproductivă. Cu toate acestea, în condiții de umiditate, ciuperca care cauzează pata Stagonospora nodorum va produce corpuri fructifere de culoare maro deschis. În plus față de diferența de culoare, aceste structuri sunt, de asemenea, mai mici decât cele produse de Septoria tritici.
Ciclul de viață
Septoria tritici supraviețuiește pe parcursul verii pe reziduurile unei culturi anterioare de grâu și inițiază infecții în toamnă. Există unele dovezi că ciuperca poate supraviețui în asociere cu alte gazde ierboase și semințe de grâu. Aceste surse ale ciupercii sunt probabil cele mai importante atunci când nu există reziduuri de grâu. Indiferent de practicile de rotație sau de gestionare a reziduurilor, există, de obicei, suficient inocul pentru a iniția infecțiile de toamnă. Septoria tritici pătarea este favorizată de vremea rece și umedă. Intervalul optim de temperatură este cuprins între 16 și 21 °C; cu toate acestea, infecțiile pot apărea în timpul lunilor de iarnă la temperaturi de până la 5 °C. Infecția necesită cel puțin 6 ore de umezeală a frunzelor, iar pentru o infecție maximă sunt necesare până la 48 de ore de umezeală. Odată ce a avut loc infecția, ciuperca are nevoie de 21 până la 28 de zile pentru a dezvolta corpurile de fructificare negre caracteristice și pentru a produce o nouă generație de spori. Sporii produși în aceste corpuri de fructificare sunt exsudați în mase lipicioase și au nevoie de ploaie pentru a fi împrăștiați pe frunzele superioare și pe căpățâni.
Infecție prin Septoria tritici
Pycnidiospori de S. tritici germinează în apă liberă de la ambele capete ale sporului sau din celulele intercalare (Weber, 1922). Germinația sporilor nu începe decât după aproximativ 12 ore de la contactul cu frunza. Tuburile germinative se dezvoltă la întâmplare pe suprafața frunzei. Weber (1922) a observat doar penetrarea directă între celulele epidermice, dar alții au concluzionat că penetrarea prin stomatele deschise și închise este principalul mijloc de pătrundere în gazdă (Benedict, 1971; Cohen și Eyal, 1993; Hilu și Bever, 1957). Kema et al. (1996) au observat numai penetrarea prin stomate. Hifele care cresc prin stomate se restrâng până la un diametru de aproximativ 1 μm, apoi se lărgesc după ce ajung în cavitatea substomatală.
Hifele cresc paralel cu suprafața frunzei sub celulele epidermice, apoi prin mezofil până la celulele din partea inferioară a epidermei, dar nu în epidermă. Nu se formează haustorii, iar creșterea hifelor este limitată de celulele sclerenchymatice din jurul fasciculelor vasculare, cu excepția cazului în care hifele sunt foarte dense. Fasciculele vasculare nu sunt invadate. Hifele cresc intercelular de-a lungul pereților celulari prin mezofil, ramificându-se la un sept sau în mijlocul celulei. Nu apar simptome macroscopice timp de aproximativ 9 zile, cu excepția unei celule moarte ocazionale, dar celulele mezofile mor rapid după 11 zile. Picnidele se dezvoltă în camerele substomatice. Hifele rareori se dezvoltă în celulele gazdei (Hilu și Bever, 1957; Kema et al, 1996; Weber, 1922).
O infecție reușită are loc numai după cel puțin 20 de ore de umiditate ridicată. Doar câteva pete maronii s-au dezvoltat dacă frunzele au rămas umede timp de 5-10 ore după depunerea sporilor (Holmes și Colhoun, 1974) sau până la 24 de ore (Kema et al., 1996). Relațiile gazdă-parazit sunt aceleași pe grânele rezistente sau sensibile. Germinația sporilor pe suprafața frunzelor este aceeași, indiferent de sensibilitate. Numărul de penetrări reușite este aproximativ același, dar creșterea ifelor este mai rapidă la cultivarele sensibile, ceea ce duce la mai multe leziuni. Hifele se extind 44 Sesiunea 2 - B.M. Cunfer dincolo de zona necrotică la toate cultivarele. O toxină poate juca un rol în patogeneză (Cohen și Eyal, 1993; Hilu și Bever, 1957). În schimb, colonizarea a fost mult redusă pe o linie rezistentă (Kema et al., 1996).
Stagonospora (Septoria) și Septoria Pathogens of Cereals: Procesul de infectare
B.M. Cunfer, Departamentul de Fitopatologie, Universitatea din Georgia, Griffin, GA
Procesul de infectare a fost studiat cel mai intens pentru Stagonospora (Septoria) nodorum și Septoria tritici. Un studiu aprofundat privind Septoria passerinii este disponibil. Aproape toate informațiile raportate se referă la infecția prin picnidiospori. Cu toate acestea, procesul de infecție pentru alte forme de spori este destul de similar. Informațiile prezentate se referă în principal la infectarea frunzelor în condiții optime. Unele studii au fost realizate cu plante de răsaduri intacte, în timp ce altele au fost realizate cu frunze detașate. Infectarea coleoptilului și a răsadului de grâu de către S. nodorum a fost descrisă în detaliu de Baker (1971) și revizuită de Cunfer (1983). Deși nu s-au făcut comparații precise, se pare că procesul de infecție are multe asemănări în fiecare sistem gazdă-parazit și este tipic pentru mulți agenți patogeni necrotrofi. Informațiile privind factorii care influențează dezvoltarea simptomelor și expresia bolii sunt excluse, dar au fost analizate de alți autori (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Este inclus un rezumat al factorilor care influențează longevitatea sporilor pe suprafața frunzelor.
Rolul Cirrusului și supraviețuirea sporilor pe suprafața frunzei Cele mai detaliate informații privind funcția de înveliș al cirrusului care încorporează picnidiosporii exsudați din picnidium sunt pentru S. nodorum. Cirrul este un gel compus din compuși proteici și zaharoizi. Compoziția și funcția sa sunt similare cu cele ale altor ciuperci din familia Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths și Peverett, 1980). Rolurile primare ale componentelor cirrului sunt protecția picnidiosporilor împotriva desecării și prevenirea germinării premature.
Cirrusul protejează picnidiosporii, astfel încât unii rămân viabili cel puțin 28 de zile (Fournet, 1969). Atunci când cirrusul a fost diluat cu apă, dacă concentrația soluției de cirrus era >20%, mai puțin de 10% de picnidiospori au germinat. La o concentrație mai mică, componentele furnizează nutrienți care stimulează germinarea sporilor și alungirea tuburilor germinative. Lungimea tuburilor germinative a crescut până la o concentrație de 15% cirrus, apoi a scăzut moderat la concentrații mai mari (Harrower, 1976). Brennan et al. (1986) au raportat o germinație mai mare în fluidul cirrus diluat. Componentele cirrusului au redus germinația la 10-60% umiditate relativă. Odată ce sporii sunt dispersați, efectele stimulatoare ale fluidului cirrus sunt probabil neglijabile (Griffiths și Peverett, 1980).
La o umiditate relativă de 35-45%, sporii de S. tritici în cirri au rămas viabile cel puțin 60 de zile (Gough și Lee, 1985). Componentele cirrilor pot acționa ca un inhibitor al germinării sporilor sau potențialul osmotic ridicat al cirrilor poate împiedica germinarea. Pycnidiosporii de S. nodorum nu au supraviețuit timp de 24 de ore la o umiditate relativă mai mare de 80% la 20 C. Sporii au supraviețuit două săptămâni sau mai mult la o umiditate relativă <10% (Griffiths și Peverett, 1980). Atunci când lichidul de cirrus al S. nodorum a fost diluat cu apă, aproximativ două treimi din picnidiospori și-au pierdut viabilitatea în 8 ore, iar după 30 de ore la lumina zilei, doar 5% au germinat. Atunci când sporii au fost depozitați la întuneric, 40% au rămas viabili după 30 de ore (Brennan et al., 1986).
Conidii uscate de S. nodorum, la umbră și în lumina directă a soarelui, au supraviețuit în aer liber cel puțin 56 de ore (Fernandes și Hendrix, 1986a). Germinația picnidiosporilor de S. nodorum a fost inhibată de UV-B continuu (280-320 nm), în timp ce germinația de S. tritici nu a fost. Extinderea tubului germinativ în condiții de UV-B continuu a fost inhibată pentru ambele ciuperci, în comparație cu întunericul (Rasanayagam et al., 1995).
Infecția cu Septoria nodorum
Procesul de penetrare a gazdei și de dezvoltare a S. nodorum în interiorul frunzei a fost examinată în detaliu de mai mulți cercetători (Baker și Smith, 1978, Bird și Ride 1981, Karjalainen și Lounatmaa, 1986; Keon și Hargreaves, 1984; Straley, 1979; Weber, 1922). Pycnidiosporii au tendința de a se depune în depresiunile dintre două celule epidermice, iar multe încercări de penetrare a frunzelor încep acolo. Sporii germinează pe suprafața frunzei ca răspuns la umiditatea liberă (Fernandes și Hendrix, 1986b). Aceștia încep să germineze la 2-3 ore de la depunere, iar după 8 ore germinația poate ajunge la 90%. Pătrunderea în frunze începe la aproximativ 10 ore după depunerea sporilor (Bird și Ride, 1981; Brönnimann et al., 1972; Holmes și Colhoun, 1974).
La începutul germinației, tubul germinativ este înconjurat de un material amorf care se atașează de frunză. Tuburile germinative care cresc de la ambele capete ale unui spor și din celulele intercalare tind să crească de-a lungul depresiunilor dintre celule și sunt adesea orientate de-a lungul axei lungi a frunzei (O'Reilly și Downes, 1986). Hifele din spori care nu se află în depresiuni cresc la întâmplare, cu ramificări ocazionale (Straley, 1979). Se formează un appressorium cu un vârf de infecție care penetrează cuticula și pereții periclinali ai celulelor epidermice direct în lumenul celular, ceea ce duce la moartea rapidă a celulelor.
Multe penetrări sunt mai întâi subcuticulare sau creșterea laterală a unei hifă are loc în interiorul peretelui celular înainte de creșterea în citoplasmă (Bird și Ride, 1981; O'Reilly și Downes, 1986). De asemenea, are loc o pătrundere prin stomate deschise și închise, care poate fi mai rapidă decât penetrarea directă (Harrower, 1976; Jenkins, 1978; O'Reilly și Downes, 1986; Straley, 1979). Tuburile germinative se ramifică la nivelul stomatelor și al joncțiunilor celulelor epidermice. Penetrarea unui tub germinativ într-un stomat poate avea loc fără formarea unui appressorium. Uneori, penetrarea are loc prin tricomi (Straley, 1979). Aparent, majoritatea încercărilor de penetrare eșuează, formându-se papile dense în celulele de la locul încercării de penetrare (Karjalainen și Lounatmaa, 1986; Bird și Ride, 1981).
După penetrare, celulele epidermice mor rapid și se lignifică, iar hifele cresc în mezofil. Celulele mezofile se deformează, iar materialul lignificat se depune în afara unor celule, care apoi se prăbușesc. Lignificarea are loc înainte ca hifele să ajungă în celulă. Procesul este același la toate soiurile, dar se dezvoltă mai lent la soiurile rezistente. Hifele se dezvoltă intercelular între celulele epidermice, apoi în mezofil. Când mezofilul este penetrat, deteriorarea cloroplastelor începe în 6-9 zile (Karjalainen și Lounatmaa, 1986).
Cu toate acestea, rata de fotosinteză începe să scadă în decurs de o zi după infecție și înainte ca simptomele să fie vizibile (Krupinsky et al., 1973). Țesutul de sclerenchyma din jurul fasciculelor vasculare previne infectarea țesutului vascular. Fasciculele vasculare blochează răspândirea hifelor prin mezofil, cu excepția cazului în care țesutul sclerenchyma este tânăr și nu este complet format (Baker și Smith, 1978).
Stagonospora nodorum eliberează o gamă largă de enzime de degradare a peretelui celular, inclusiv amilază, pectină metil esterază, poligalacturonaze, xilanaze și cellaze in vitro și în timpul infecției frunzelor de grâu (Baker, 1969; Lehtinen, 1993; Magro, 1984). Informațiile legate de degradarea peretelui celular de către enzime sunt în concordanță cu observațiile histologice. aceste enzime pot acționa în combinație cu toxinele. Sensibilitatea enzimatică poate fi legată de rezistență și de rata de colonizare fungică (Magro, 1984). La fel ca mulți necrotrofi, agenții patogeni Septoria și Stagonospora produc compuși fitotoxici in vitro. Deteriorarea ș i moartea celulelor înainte de dezvoltarea hifelor în țesutul mezofil (Bird ș i Ride, 1981) sunt în concordanță cu producția de toxine. Cu toate acestea, nu s-a stabilit un rol definitiv al toxinelor în procesul de infecție și nici relația lor cu rezistența gazdei (Bethenod et al, 1982; Bousquet et al, 1980; Essad și Bousquet, 1981; King et al, 1983). Diferențele în ceea ce privește gama de gazde între tulpinile adaptate la grâu și orz de S. nodorum poate fi legată de producția de toxine (Bousquet și Kollmann, 1998). Inițierea germinării sporilor și procentul de spori germinați nu sunt influențate de sensibilitatea gazdei (Bird și Ride, 1981; Morgan 1974; Straley, 1979; Straley și Scharen, 1979; Baker și Smith, 1978).
Bird și Ride (1981) au raportat că extinderea tuburilor germinative pe suprafața frunzelor a fost mai lentă la cultivarele rezistente decât la cele sensibile. Acest mecanism, exprimat la cel puțin 48 de ore de la depunerea sporilor, indică o rezistență la alungirea tuburilor germinative înainte de penetrare. Au fost mai puține penetrări reușite la cultivarele rezistente, iar penetrarea a decurs mai lent la cultivarele rezistente (Baker și Smith, 1978; Bird și Ride, 1981). S-a propus ca lignificarea să limiteze infecția atât la cultivarele rezistente, cât și la cele sensibile, dar alți factori au încetinit dezvoltarea ciupercii la liniile rezistente. La liniile sensibile, hifele cu creștere mai rapidă pot scăpa de lignificarea celulelor gazdă.La patru zile după inocularea orzului cu un izolat de biotip de grâu de S. nodorum, hifele au crescut prin cuticulă și, uneori, în straturile exterioare de celuloză ale pereților celulari epidermici. Sub hifele de penetrare s-au depus papile groase, iar celulele nu au fost penetrate (Keon și Hargreaves, 1984).
Infecția cu Septoria passerinii: Green și Dickson (1957) prezintă o descriere detaliată a procesului de infectare a S. passerinii pe orz. Procesul de infectare este similar cu S. tritici. Ca și în cazul lui S. tritici, durata de timp necesară pentru penetrarea frunzelor este considerabil mai mare decât pentru S. nodorum. Tuburile germinative se ramifică și cresc pe suprafața frunzei la întâmplare, dar uneori de-a lungul depresiunilor dintre celulele epidermice. Pătrunderea în frunze se face aproape exclusiv prin stomate. Hifele germinative se umflă, iar dacă penetrarea nu reușește, hifele continuă să se alungească. La 48 de ore de la depunerea sporilor, nu se mai produce nicio penetrare. După 72 de ore, tuburile germinative se îngroașă peste stomate, se dezvoltă între celulele de gardă și pe fețele celulelor de acces și în cavitățile substomatice. Pătrunderea directă între celulele epidermice este observată doar rareori.
Germinația sporilor și penetrarea în gazdă sunt identice la cultivarele rezistente și la cele sensibile. Există o extindere mult mai mică a hifelor în interiorul frunzelor la cultivarele rezistente, iar papilele sunt observate pe mulți, dar nu pe toți pereții celulari. Hifele se extind sub epidermă de la un stoma la altul, dar nu pătrund între celulele epidermice. Mezofilmul este colonizat, dar nu se formează haustorii. După ce celulele mezofile devin necrotice, celulele epidermice se prăbușesc. Dezvoltarea miceliului în frunză este rarefiată și, de obicei, blocată de fasciculele vasculare. La frunzele mai tinere, dacă teaca vasculară este mai puțin dezvoltată, hifele trec între fascicul și epidermă. Picnidele se formează în cavități substomatice, mai ales pe suprafața superioară a frunzei (Green și Dickson, 1957).
Factori care afectează longevitatea sporilor pe suprafața frunzelor Dintre agenții patogeni Stagonospora și Septoria de la cereale, informații definitive privind procesul de infecție au fost raportate doar pentru S. nodorum, S. tritici, și S. passerinii. La fel ca mulți alți agenți patogeni necrotrofi, niciunul dintre cei doi grupuri de agenți patogeni nu provoacă reacția de hipersensibilitate. O diferență semnificativă în procesul de infecție între agenții patogeni Septoria și Stagonospora este că germinarea și penetrarea sporilor se desfășoară mult mai rapid în cazul S. nodorum decât pentru S. tritici și S. passerinii. Acest lucru are o influență semnificativă asupra epidemiologiei bolilor.
Agenții patogeni Septoria pătrund în plantă în primul rând prin stomate, în timp ce S. nodorum pătrunde atât direct, cât și prin stomate. S. nodorum penetrează și ucide rapid celulele epidermice, dar S. tritici și S. passerinii nu ucid celulele epidermice până când hifele nu se ramifică prin mezofilul frunzei și începe necroza rapidă. Studiile histologice privind creșterea fungică în urma pătrunderii în gazdă corespund datelor generate de studiile epidemiologice privind rezistența gazdei. Rezistența încetinește rata de colonizare a gazdei, dar nu are un efect apreciabil asupra procesului de dezvoltare a leziunilor.
Mecanismele care controlează răspunsul gazdei, fie că sunt legate de enzime și toxine sau de alți metaboliți eliberați de agenții patogeni în timpul infecției, sunt încă neclare. Există puține informații despre infecția prin ascospori. Procesul de infectare este probabil foarte asemănător cu cel al picnidiosporilor. Ascosporii de Phaeosphaeria nodorum germinează într-o gamă largă de temperaturi, iar tuburile lor germinative pătrund direct în frunze. Cu toate acestea, conform Rapilly et al. (1973), ascosporii, spre deosebire de picnidiospori, nu germinează în apă liberă.
Model de infecție cu Septoria spp.
Infecțiile cu septorioză sunt posibile la temperaturi scăzute, în timp ce temperaturile sub 7°C pot să nu ducă la infecții în decurs de 2 zile. Temperatura optimă a bolii este atinsă în zona de 16-21°C. Infecțiile sunt posibile într-o perioadă de umiditate relativă ridicată sau de umezeală a frunzelor de 14 ore sau mai mult. Pentru a îndeplini aceste condiții, am decis să separăm modelele pentru infecții slabe, moderate și severe. Infecțiile slabe pot fi date în cazul în care este posibil ca agentul patogen să infecteze țesutul gazdei. Acest lucru înseamnă că infecțiile slabe pot avea loc dacă temperaturile sunt minime, iar perioadele de umezeală a frunzelor au o durată critică. O infecție moderată va avea loc în condițiile în care majoritatea testelor de infecție conduc la rezultate rezonabile, iar infecțiile severe au loc în condițiile în care agentul patogen are condiții optime de infectare.

Începe infecția după o ploaie de 0,5 mm. Am decis să nu folosim un model pentru formarea picnidiei. Condiția necesară pentru formarea picnidiei este presupusă a fi o perioadă cu o umiditate relativă mai mare de 85%. Se estimează că durata de viață a picnidiei este de 24 de ore. În toate climatele în care Septoria tritici are șansa de a se infecta, vom găsi 2 ore care îndeplinesc aceste condiții aproape în fiecare zi în jurul răsăritului soarelui.
Evaluarea severității infecției: Pentru a putea evalua presiunea de infecție cu Septoria tritici între stadiul 10 (prima frunză de pe coleoptil) și stadiul 32 (nodul doi la cel puțin 2 cm deasupra nodului 1) și între 32 și 51 (începutul epilării), trebuie să evaluăm severitatea infecțiilor în funcție de condițiile climatice. Această evaluare se face pe o scară de la 1 la 5. Gradul de gravitate 1 este dat dacă sunt îndeplinite condițiile pentru o infecție slabă și a plouat mai puțin de 5 mm, în caz contrar, gradul de gravitate corespunzător va fi 2. Se acordă un grad de severitate 3 dacă este îndeplinită o infecție moderată și a plouat mai puțin de 5 mm. Dacă a plouat mai mult de 5 mm în timpul unei infecții moderate sau mai puțin de 5 mm în timpul unei infecții severe, se acordă un grad de gravitate 4.
O infecție severă, cu mai mult de 5 mm de ploaie, corespunde unei valori de gravitate de 5.
Septoria tritici evaluarea presiunii bolii: Clima este doar unul dintre factorii care influențează presiunea bolilor în câmp. Ceilalți doi factori sunt istoricul câmpului și susceptibilitatea soiului cultivat. Dacă putem acumula valorile severității bolii din stadiul 10 până în stadiul 32 la valoarea 4, ne putem aștepta la o presiune slabă a bolii din cauza climei. Dacă această valoare ajunge la 6, ne putem aștepta la o presiune moderată a bolii, iar dacă ajunge la 10, ne putem aștepta la o presiune ridicată a bolii din partea climei. Cunoașterea sensibilității soiului și a istoricului câmpului ne va determina să pulverizăm sau nu la o presiune slabă sau moderată a bolii în această situație. Dacă valoarea acumulată este de 10, poate duce oricum la o pulverizare în stadiul 32. Decizia de a face o pulverizare într-un stadiu ulterior depinde mai mult de climatul de primăvară. Dacă reușim să acumulăm valorile de severitate din stadiul 10 până la o valoare de 6, ne putem aștepta la o presiune slabă a bolii. Dacă această valoare ajunge la 10, ne putem aștepta la o presiune moderată a bolii, iar dacă această valoare ajunge la 15, ne putem aștepta la o presiune ridicată a bolii, în funcție de situația climatică.
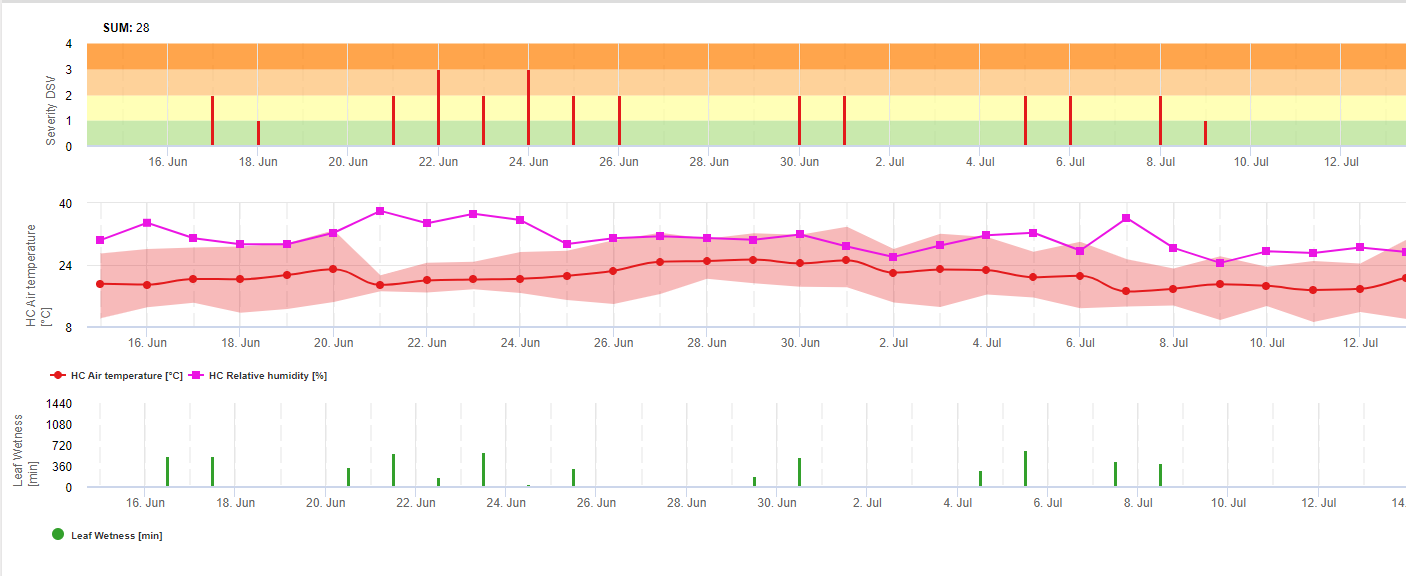
În FieldClimate arătăm Septoria tritici Severitatea împreună cu cele trei grade de infecție diferite într-un singur grafic (a se vedea mai sus). Din cauza precipitațiilor și a perioadelor lungi de umezeală a frunzelor, condițiile pentru o infecție severă cu S. tritici au fost fullfield pe 14 și 16 mai. Nivelurile de gravitate ating cea mai mare valoare de 5 pe 14 mai, ceea ce înseamnă că există acum un risc ridicat de infectare.
Biologia infecției Stagonospora nodorum diferă într-o oarecare măsură de cea a S. tritici, dar această diferență nu este suficient de mare pentru un model separat. Prin urmare, sugerăm să se utilizeze acest model pentru întregul complex de boli ale Stagnospora și Septoria la cereale, inclusiv S. passerinii. S. tritici și S. passerinii au tendința de a avea nevoie de perioade mai lungi de umezire a frunzelor decât S. nodorum. În zonele cu o presiune ridicată de S. nodorum infecțiile clasificate ca fiind slabe, cu o valoare de gravitate 2, trebuie tratate mai serios decât în alte domenii.
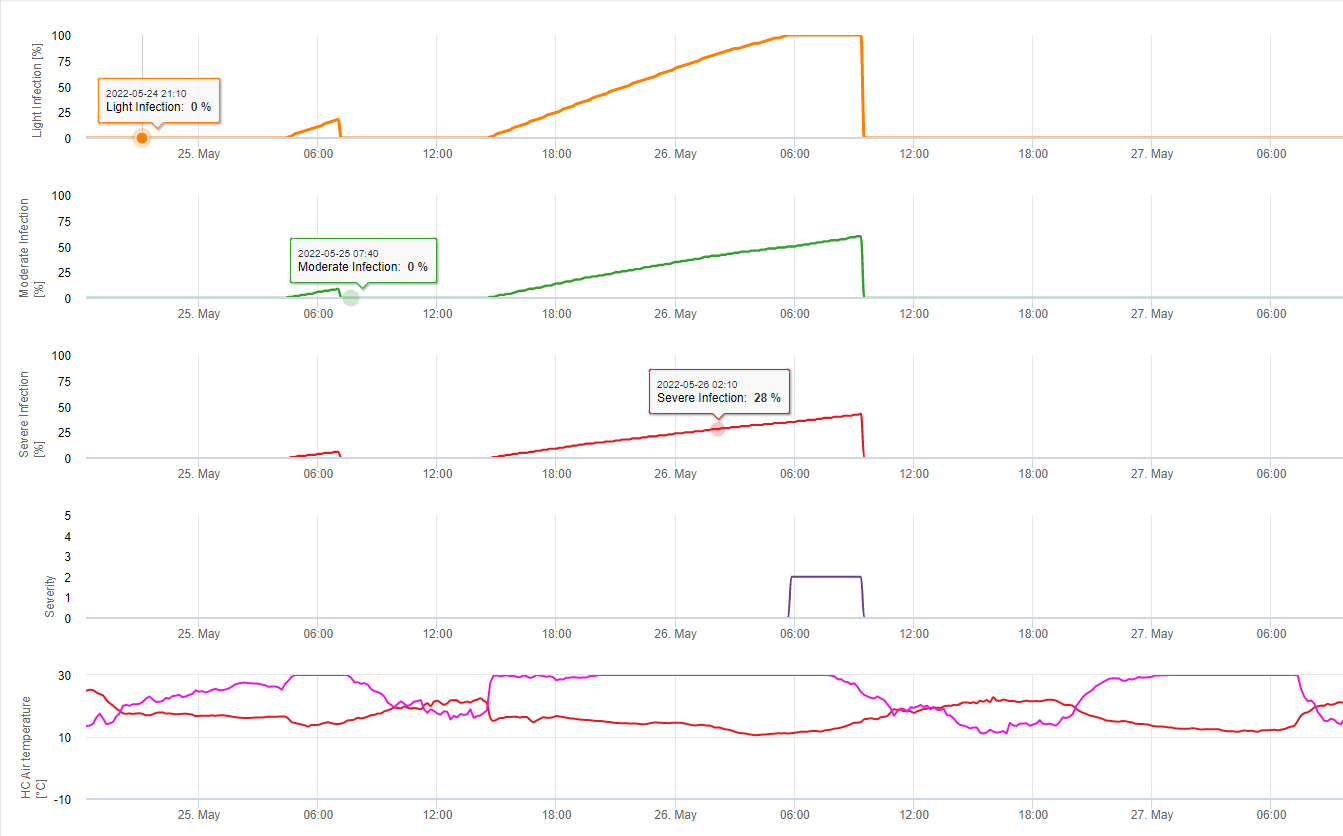
Pentru Septoria nodorum un model de risc este prezentat în FieldClimate (a se vedea mai sus). Un risc ridicat a fost determinat la 17 iunie și 7 iulie (100%). În funcție de stadiul plantelor sensibile la infecție, trebuie luate în considerare măsurile de protecție a plantelor dacă riscul ajunge la 80% (a se vedea, de asemenea, prognoza meteo, protecția plantelor). În cazul în care riscul a fost de 100% și s-a constatat deja o infecție, trebuie luate măsuri de protecție sistemică a plantelor (aplicare curativă) pentru a proteja planta.
Mai multe specii de ciuperci fitopatogene din genul Colletotrichum provoacă antracnoza la ardei și la multe alte legume și fructe. Până la sfârșitul anilor 1990, antracnoza ardeilor și a roșiilor era asociată doar cu fructele coapte sau în curs de coacere. De atunci, s-a stabilit o formă mai agresivă a bolii. Această formă atacă ardeii în orice stadiu de dezvoltare a fructelor și poate amenința profitabilitatea culturilor de ardei în zonele în care se instalează. Această boală poate afecta, de asemenea, roșiile, căpșunile și, eventual, alte culturi de fructe și legume.
Simptomele
Leziunile circulare sau unghiulare se dezvoltă pe fructele imature de orice mărime. Adesea se formează leziuni multiple pe fiecare fruct în parte. În cazul în care boala este severă, leziunile se pot unifica. Adesea, pe suprafața leziunilor se formează mase de spori de ciupercă de culoare roz sau portocalie în inele concentrice. În cazul leziunilor mai vechi, se pot observa structuri negre numite acervuli. Cu o lentilă de mână, acestea arată ca niște puncte negre mici; la microscop, ele arată ca niște smocuri de fire de păr negre mici. Agentul patogen formează sporii rapid și abundent și se poate răspândi rapid în toată cultura de ardei, ducând la pierderi de producție de până la 100%. Leziunile pot apărea, de asemenea, pe tulpini și frunze sub forma unor pete maro de formă neregulată, cu margini maro închis.
Agenți patogeni
Această formă de antracnoză a piperului este cauzată de ciuperca Colletotrichum acutatum. Agentul patogen supraviețuiește pe resturile vegetale provenite din culturile infectate și pe alte specii de plante sensibile. Ciuperca nu se transmite în sol pentru perioade lungi de timp în absența resturilor vegetale infestate. Ciuperca poate fi, de asemenea, introdusă într-o cultură prin semințe infestate. În timpul perioade calde și umede, sporii sunt stropiți de ploaie sau de apa de irigare de la fructe bolnave la fructe sănătoase. Fructele bolnave acționează ca o sursă de inoculum, permițând ca boala să se răspândească de la o plantă la alta în cadrul câmpului. Ciuperca supraviețuiește în și pe semințe. Antracnoza este introdusă în câmp pe transplanturi infectate sau poate supraviețui între sezoane în resturile de plante sau pe plantele gazdă din buruieni. Gazdele alternative includ buruienile și alte plante din familia Solanaceae (tomate, cartofi, vinete), deși infecțiile acestor gazde sunt extrem de rare în Florida. Fructele sunt infectate atunci când sporii ciupercii sau resturile infestate sunt stropiți de ploaie pe plantele de ardei. Noi spori sunt produși în țesutul infectat și apoi sunt împrăștiați pe alte fructe. Lucrătorii pot, de asemenea, să mute sporii cu ajutorul echipamentelor sau în timpul manipulării plantelor infectate. Infecția are loc, de obicei, pe vreme caldă și umedă. Temperaturile din jurul 80° F (27° C) sunt temperaturile optime. pentru dezvoltarea bolii, deși infecția apare atât la temperaturi mai ridicate, cât și la temperaturi mai scăzute. Pierderi grave apar în timpul ploilor, deoarece sporii sunt spălați sau împrăștiați pe alte fructe, ceea ce duce la mai multe infecții. Boala este mai probabil să se dezvolte pe fructele mature care sunt prezente o perioadă lungă de timp pe plantă, deși poate apărea atât pe fructe imature, cât și pe cele mature. Antracnoza poate infecta la temperaturi cuprinse între 15 °C și 30 °C. Dar o umezeala frunzelor lungi este necesară pentru a satisface nevoile unei infecții. La temperatura optimă de la 20°C la 25°C încă 12 ore de umezeală a frunzelor sunt necesare. Temperaturile mai ridicate sau mai scăzute vor necesita perioade și mai lungi de umezeală a frunzelor (nu este nevoie de o funcție liniară/un tablou pentru calcul). FieldClimate calculează posibilele evenimente de infecție pe baza umezelii frunzelor și a temperaturilor din timpul acestui eveniment.
Biologie a Plasmopara viticola (Miliardul)
P. viticola este un parazit obligat. Ceea ce înseamnă că are nevoie de organe de viță de vie verzi și proaspete pentru a se dezvolta. În timpul perioadei fără vegetație, persistă formând corpuri de fructificare, așa numitele oospori. Oosporii oomicetelor pot supraviețui perioade foarte lungi în sol. Prin urmare, putem găsi putregaiul în locuri unde infecțiile nu sunt posibile în fiecare an. Primăvara, când solul superior este suficient de umed și cald, oosporii formează așa-numitele macrosporangii care pot elibera până la 200 de zoospori în apa liberă. Zoosporii se deplasează până la frunze și la ciorchini prin vânt în picături de apă. Au doi flageli și se deplasează în pelicula de apă de pe partea inferioară a frunzelor, a ciorchinilor și a boabelor tinere pentru a găsi un stom pentru a pătrunde în țesutul plantei. Ele intră și germinează în stoma, în care își transferă toată plasma în mai puțin de o oră. În cadrul studiilor microscopice, găsirea stomei, încifrarea și germinarea în stomă s-au încheiat în 90 de minute.
P. viticola se dezvoltă în spațiul intercelular și se hrănește prin intermediul haustoriilor care pătrund în celulele epidermice și parenchimatoase. În funcție de temperatură și de umiditatea relativă, dezvoltă o creștere intercelulară suficientă cu suficiente haustorii pentru a forma un corp substomaterial care umple întreaga zonă substomaterială și care ridică țesutul epidermic din țesutul parenchimatos. Acest lucru duce la apariția simptomului vizibil al petei de ulei.
Oomicetele sunt sporularea în absența luminii, atunci când umiditatea relativă este foarte mare. În P. viticola nu există sporulație dacă temperaturile sunt sub 12°C și umiditatea relativă este sub 95%. Sporangioforii sunt formați de veziculele substomatelor și vor ieși din stoma. Sporangii proaspăt formați sunt lipicioși și pot fi îndepărtați din sporangiafori doar cu ajutorul apei. În timpul scăderii umidității relative, sporangii devin mai greu de îndepărtat și pot fi îndepărtați și de vânt.
Sporangia va elibera până la 20 de zoospori în apa liberă. Acești zoospori trebuie să fie distribuiți de vânt în picături de apă pentru a ajunge și pe frunzele proaspete, sau sporangii pot fi distribuiți de ploaie sau de vânt. Procesul de infectare a infecției primare și secundare este același.
Datorită importanței mari a stadiului sexual pentru hibernarea agentului patogen, putem presupune că tipurile de împerechere se potrivesc în toate podgoriile în care apare putregaiul viței de vie. Formarea zoosporelor are loc pe frunzele mai bătrâne în timpul verii și la începutul toamnei.
Infecția primară în detaliu
Infecțiile care provin din oospori se numesc infecții primare. Acest termen este înșelător, deoarece la începutul verii pot apărea mai multe infecții primare. În cadrul evenimentelor epidemiologice, infecția primară nu joacă un rol important dacă există suficiente pete de ulei în podgorii și dacă potențialul de infecție al sporilor de vară (sporangii) depășește oosporii.
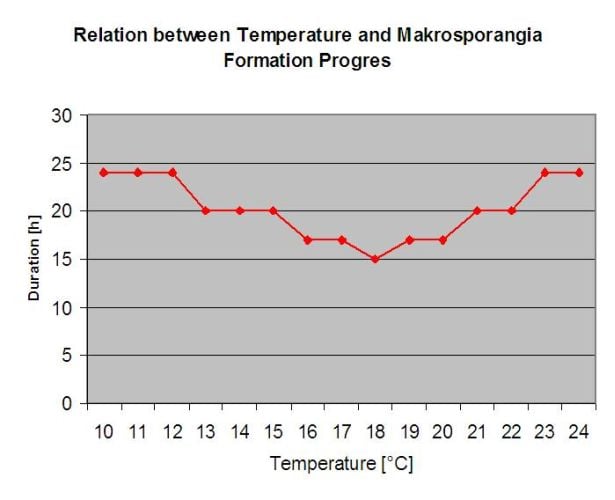
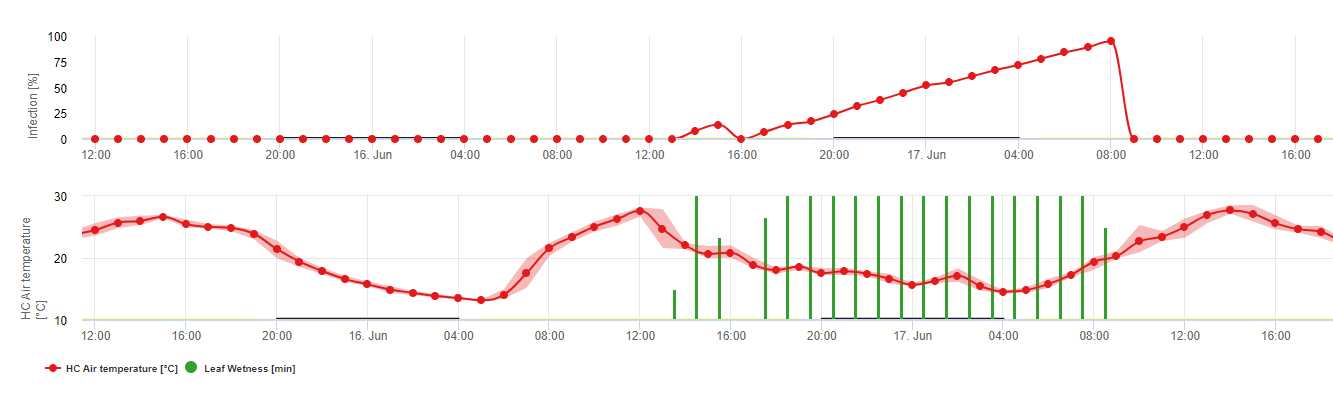
Oosporii care iernează se dezvoltă în așa-numitele macrosporangii atunci când există suficientă umiditate relativă timp de aproximativ 24 de ore. Macrosporangia își eliberează zoosporii în apa liberă și în ploile puternice (ca în cazul unei furtuni). Sporii sunt transportați pe frunzele viței de vie și/sau pe lăstarii verzi. Prin urmare, infecțiile primare, au nevoie de ploaie mai lungă perioade sau mai multe furtuni succesive. Prima ploaie face ca frunzele căzute să fie saturate cu apă, iar ploile puternice din zilele următoare fac ca macrosporangii să își elibereze zoosporii, care ajung pe frunzele sau lăstarii viței de vie. Pentru ca infecția să reușească, este necesară o perioadă suficient de lungă de umezeală a frunzelor, pentru a permite zoosporilor să ajungă la stoma frunzelor sau lăstarilor și să îi infecteze (ilustrația de mai sus prezintă ciclul de dezvoltare a putregaiului pufos al viței de vie (Plasmopara viticola).
Modelul pentru infecția primară a putregaiului verifică mai întâi dacă vremea este potrivită pentru dezvoltarea macrosporangiei. Acesta este cazul atâta timp cât frunzele sunt umede sau dacă umiditatea relativă după ploaie nu scade sub 70%. În funcție de temperatură, macrosporangii se pot dezvolta în decurs de 16 până la 24 de ore. În cazul în care există macrosporangii mature, acest lucru va fi indicat grafic în afișarea infecției primare de Downy Mildew. În cazul în care sunt prezente macrosporangii, o ploaie puternică poate răspândi zoosporii acestora. O ploaie continuă de 5 mm este interpretată ca o ploaie puternică și zoosporii sunt răspândiți, fiind posibilă o infecție primară dacă umezeala frunzelor a durat suficient de mult timp.
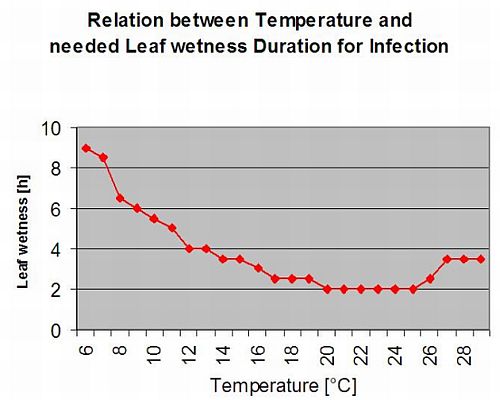
Infecții secundare
Infecții secundare ale Plasmopara viticola sunt posibile numai dacă în podgoria dumneavoastră sau în apropiere există pete de ulei deja mature. Petele de ulei sunt mature atunci când sunt capabile să sporuleze (să producă sporangi). Sporangiile sunt produse numai de noapte. Lumina soarelui inhibă sporularea. Sporangii se produc dacă este mai cald de 12°C și umiditate relativă este foarte mare. Rata de producere a sporangiei crește odată cu temperatura până la 24°C. Temperatura optimă pentru sporularea la soiurile de struguri europeni (Vitis vinifera) este de aproximativ 24°C. Dacă temperaturile depășesc 29°C, nu poate avea loc sporularea. În modelul nostru, verificăm dacă umiditate mai mare de 95% are loc în timpul nopții. În cazul în care această condiție durează pentru o temperatură orară cumulată de peste 50°C, sporulația s-a terminat și se formează noi sporangi de Plasmopara există în podgorie. De exemplu, orele de 50°C corespund la 4 ore cu 13°C sau la 3 ore cu 17°C.
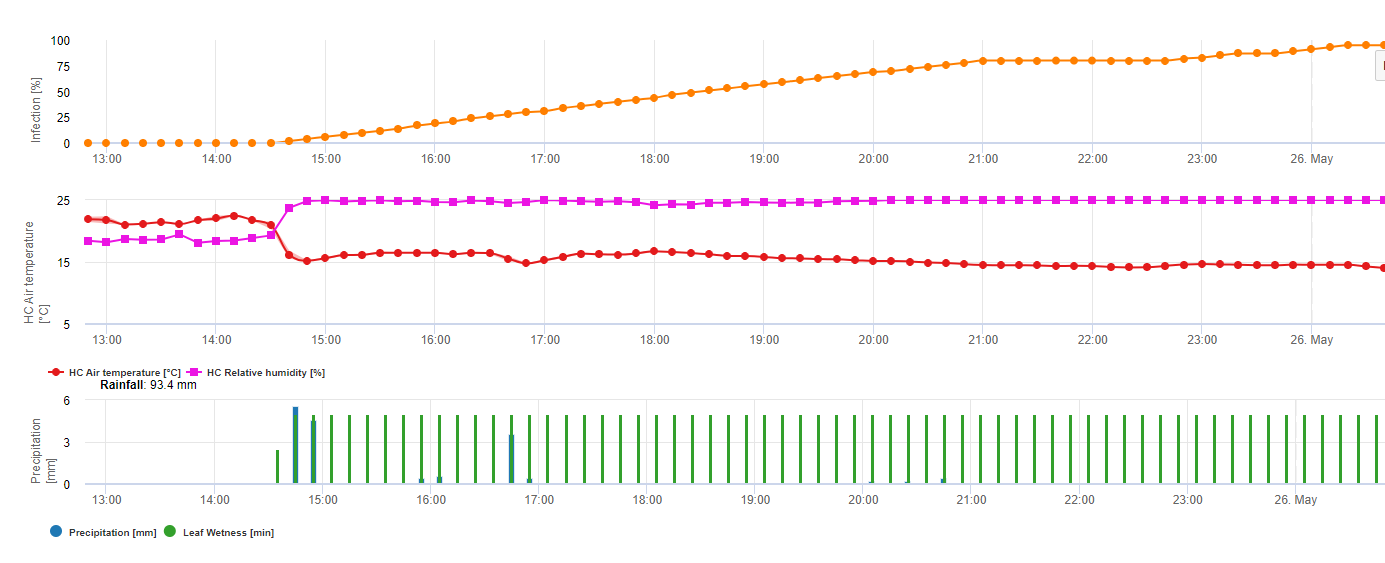
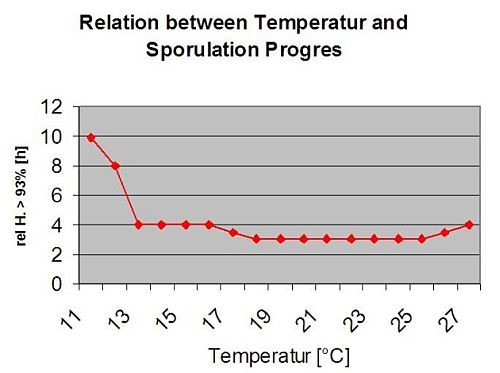
Graficul de mai jos arată relația dintre temperatură și condițiile de umezeală (umiditate relativă, umezeala frunzelor, precipitații) care sunt folosite pentru a modela evenimentele de infecție în fieldclimate.com.
Oribusul tomatelor poate fi cauzat de trei agenți patogeni la nivel mondial.
Leveillula taurica (Oidiopsis taurica) este un agent patogen al unei game largi de specii gazdă din climatele calde aride și semiaride din Asia, Marea Mediterană, Africa și, mai recent, din sud-vestul Statelor Unite.
Erysiphe orontii (E. cichoracearum și E. polyphaga) este o altă specie comună pentru multe plante gazdă din regiunile temperate și tropicale.
Și ca o a treia specie Odium lycopersicum.
Factori de dezvoltare a bolii
- niveluri de umiditate relativă > 50% (RH optimă > 90%)
- nu este necesară prezența apei libere pe suprafața frunzelor
- interval de temperatură: 10-35 °C (cel mai bine sub 30 °C)
Oribusul este o boală determinată de inoculum. Prin urmare, doar perioadele de risc ar putea fi determinate, factorul principal pentru daune, de exemplu, focarul este inoculul inițial (care este activ într-o gamă largă de temperaturi). Așadar, pentru strategiile de control: combinați modelarea perioadei de risc cu monitorizarea inoculului fungic (boala) în câmp!
Tomate de câmp deschis
Rugina sparanghelului Puccinia asparagi este favorizată de un climat umed și cald. Cantități mici de ploaie și vânt vor distribui urediasporii, în timp ce ploile abundente le pot spăla până la sol. Din acest motiv, o perioadă de umezeală a frunzelor cu mai mult de 10 mm de ploaie nu este luată în considerare pentru o perioadă de infecție. Perioadele de umezeală a frunzelor cauzate de ploi ușoare și de rouă de 3 până la 8 ore, cu temperaturi de la 25°C până la 10°C, pot constitui perioade de infecție pentru urediaspori în timpul verii și la sfârșitul verii.
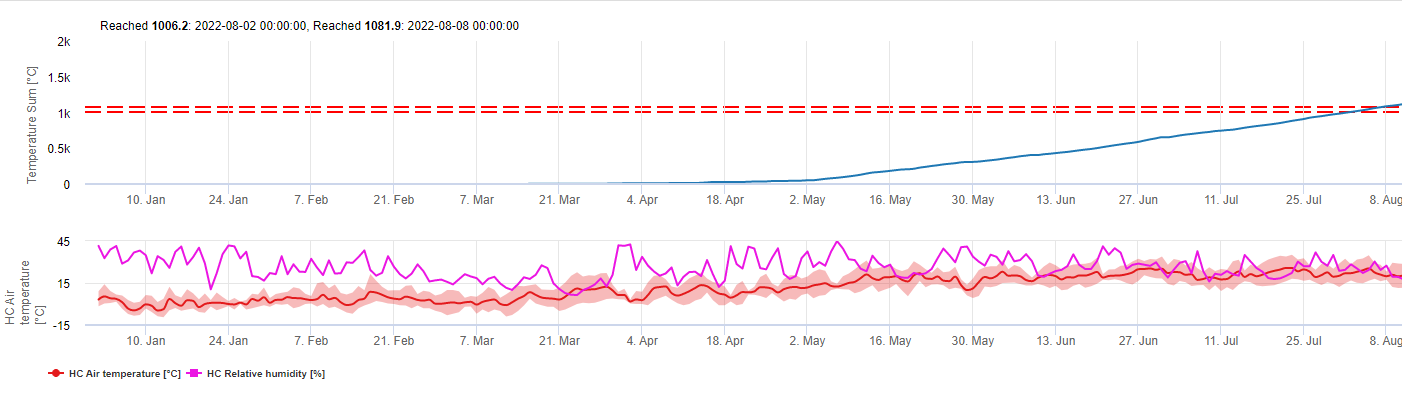
Acest model estimează data primei apariții a C. beticola pe baza temperaturilor medii zilnice cumulate de la 1 ianuarie la peste 5°C. Se ia pragul de 1006,2°C în perioadele cu umiditate relativă mai mare de 60%, iar dacă nu am avea umiditate relativă este valabil un prag de 1081,9°C.
Acest model a fost dezvoltat pentru Italia și este utilizat în Italia și Germania.
Pentru anul 2010, în Styria, CercoPrim indică data de 19 iunie pentru prima pulverizare. Aceasta este aproximativ aceeași dată pe care modelul de risc Pessl Instruments sau modelul DIV ar fi indicat-o pentru varietățile sensibile la germinare.
Agentul patogen
Boala petelor purpurii pe sulițele de sparanghel și ferigă este cauzată de ciuperca Stemphylium vesicarium. Ciuperca supraviețuiește iarna sub formă de spori sexuați (ascospori) într-un sac (ascus) produs în structuri de iernare (pseudotecii) care apar ca mici puncte negre pe resturile de sparanghel din sezonul precedent. Site-ul ascosporii sunt eliberați din ascus prin ploaie și pot fi purtate de vânt până la plantele de sparanghel abia răsărite, unde provoacă infecția primară a sezonului de creștere. Aceste noi infecții dau naștere la spori (conidii) produși printr-un proces asexuat, care, la rândul lor, pot provoca infecții secundare, proces care se repetă atât timp cât temperaturile și precipitațiile sunt favorabile.
Boala
Apariția petelor purpurii ca o problemă semnificativă în producția de sparanghel din Michigan se poate datora adoptării unui sistem de cultură "no- till", prin care feriga uscată din sezonul precedent este tăiată în aprilie și lăsată la suprafața solului. Acest reziduu persistă pe toată durata sezonului de recoltare (jumătatea lunii iunie) și este vizibil pe toată perioada de creștere a ferigii (de la sfârșitul lunii iunie până în septembrie), fiind sursa ascosporilor care declanșează infecțiile la începutul sezonului de creștere. Simptomele bolii sunt: Boala apare sub forma unor pete numeroase, ușor înfundate, violacee, cu centrul maro, care apar pe sulițele și ferigile recoltate. Leziunile de pe sulițe se găsesc adesea pe partea dinspre vânt, deoarece nisipul care suflă provoacă răni care favorizează infecția. În anii de epidemie, petele pot apărea pe 60-90% din sulițe și pot duce la respingerea recoltei, în special în cazul vânzărilor pe piața proaspătă. Petele apar, de asemenea, pe ferigile sparanghelului, afectând tulpina principală, ramurile secundare și acele (cladofile). O infecție severă a ferigii poate duce la defolierea prematură a plantei. Creșterea severității bolii cu pete purpurii este asociată cu perioade prelungite de precipitații, ceață sau rouă.
Credit text: Mary K. Hausbeck, profesor și specialist în extensie, Universitatea de Stat din Michigan, Departamentul de Fitopatologie, E. Lansing.
TomCast pentru sparanghel
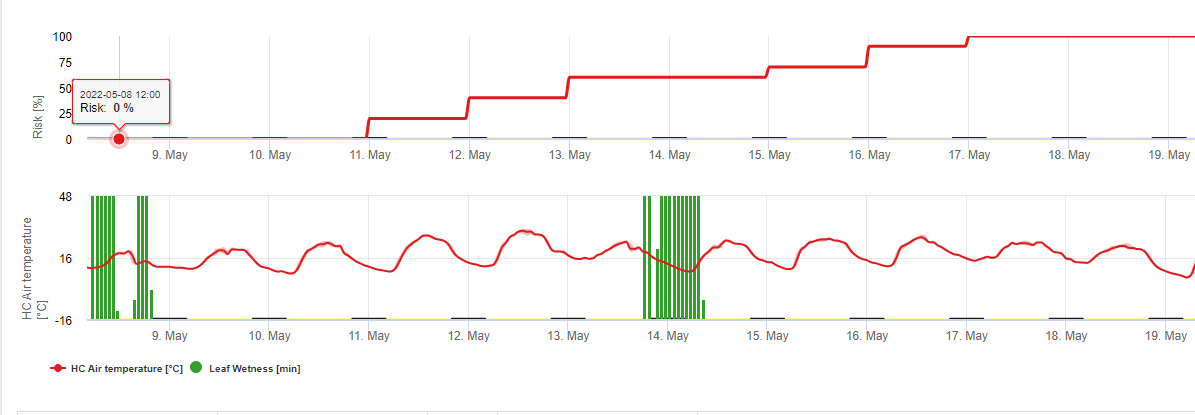
Context: TOMCAST (TOMato disease foreCASTing) este un model computerizat bazat pe date de teren care încearcă să prezică dezvoltarea bolilor fungice, și anume Bacteria timpurie, pătarea frunzelor cu Septoria și antracnoza pe roșii. Aparatele de înregistrare a datelor amplasate pe teren înregistrează din oră în oră datele privind umiditatea frunzelor și temperatura. Aceste date au fost analizate pe o perioadă de 24 de ore și pot duce la formarea unei valori de gravitate a bolii (DSV); în esență, o creștere a dezvoltării bolii. Pe măsură ce DSV se acumulează, presiunea bolii continuă să crească asupra culturii. Atunci când numărul de DSV acumulate depășește intervalul de pulverizare, se recomandă aplicarea unui fungicid pentru a atenua presiunea bolii.
TOMCAST este derivat din modelul original F.A.S.T. (Forecasting Alternaria solani on Tomatoes) dezvoltat de Dr. Madden, Pennypacker și MacNab de la Pennsylvania State University (PSU). Modelul F.A.S.T. de la PSU a fost modificat ulterior de Dr. Pitblado de la Colegiul Ridgetown din Ontario în ceea ce acum recunoaștem ca fiind modelul TOMCAST utilizat de Ohio State University Extension.
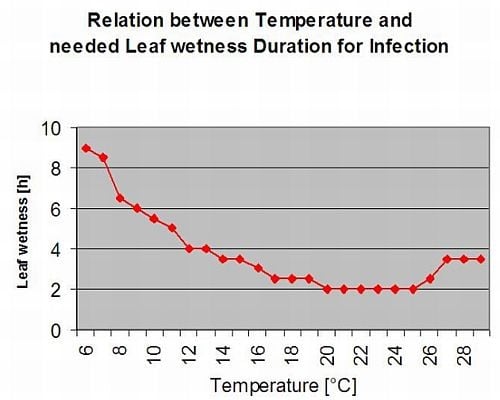
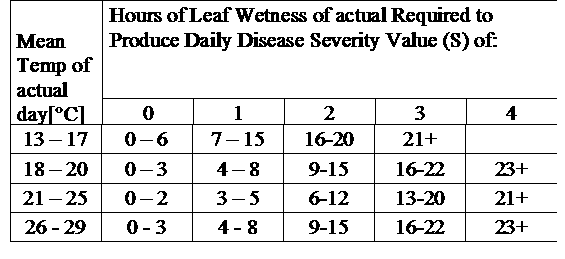
DSV sunt: O valoare de gravitate a bolii (DSV) este unitatea de măsură dată unui anumit grad de dezvoltare a bolii (arsură timpurie).
Cu alte cuvinte, un DSV este o reprezentare numerică a rapidității sau încetinelii cu care se acumulează boala (arsura timpurie) într-un câmp de tomate. DSV este determinat de doi factori: umezeala frunzelor și temperatura în timpul orelor de "umezeală a frunzelor". Pe măsură ce numărul de ore de umezeală a frunzelor și temperatura crește, DSV se acumulează într-un ritm mai rapid. Consultați graficul de mai jos privind valoarea de severitate a bolilor.
Dimpotrivă, atunci când sunt mai puține ore de umezeală a frunzelor și temperatura este mai scăzută, DSV se acumulează încet sau deloc. Atunci când numărul total de DSV acumulate depășește o limită prestabilită, numită interval de pulverizare sau prag, se recomandă o pulverizare cu fungicid pentru a proteja frunzele și fructele de dezvoltarea bolii.
Intervalul de pulverizare (care determină când trebuie să pulverizați) poate varia între 15-20 DSV. DSV-ul exact pe care ar trebui să-l folosească un cultivator este de obicei furnizat de către procesator și depinde de calitatea fructelor și de utilizarea finală a roșiilor. Urmarea unui interval de pulverizare de 15 DSV reprezintă o utilizare conservatoare a sistemului TOMCAST, ceea ce înseamnă că veți pulveriza mai des decât un cultivator care utilizează un interval de pulverizare de 19 DSV cu sistemul TOMCAST. Compromisul constă în numărul de stropiri aplicate în timpul sezonului și în potențialul de diferență în ceea ce privește calitatea fructelor.
 TomCast cu sparanghel
TomCast cu sparanghel
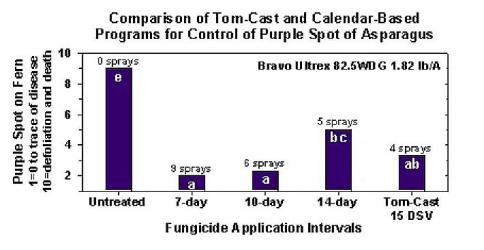
Cercetările au stabilit că sistemul de prognoză a bolilor Tom-Cast este o alternativă promițătoare la pulverizarea calendaristică a ferigii în câmpurile comerciale de sparanghel. Tom-Cast îi avertizează pe cultivatori atunci când condițiile de mediu sunt favorabile pentru dezvoltarea bolii de pătarea purpurie (perioade prelungite de rouă sau de ploaie însoțite de temperaturi ridicate). Fungicidele eficiente aplicate în conformitate cu previziunile Tom-Cast permit cultivatorilor să gestioneze boala petelor purpurii la sparanghel, economisind bani și protejând în același timp mediul înconjurător.
Credit text: Jim Jasinski, coordonator TOMCAST pentru OHIO, INDIANA și MICHIGAN
Model de infecție cu pată purpurie
The Stemphylium versicarium Modelul de infecție se bazează pe lucrările lui SUHERI și PRICE în cazul cepei și pe cele ale lui LLorent, VILARDELL, BUGIANI, GHERARDI și MONTESINO în cazul perelor.
Se calculează curbele de infecție pentru infecții ușoare, moderate și severe.
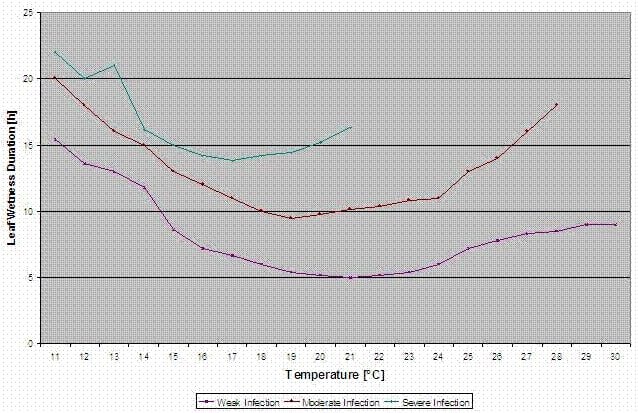
Cu această separare în situații favorabile, moderat favorabile și foarte favorabile, este la latitudinea cultivatorului să decidă cât de mare va fi presiunea bolii într-un anumit câmp și dacă trebuie să acopere o anumită infecție.
Condiții:
Temperatura: 11-30°C
Umiditatea frunzelor > 0 (început pentru infecție) sau umiditate relativă > 90% (doar dacă a existat deja o infecție înainte de calcul și încă se menține).
infecție ușoară (11-30°C), infecție moderată (13-30°C), infecție severă (16-30°C), o matrice cu durata umezelii frunzelor (vezi mai sus).
Resetare: dacă umiditatea relativă este mai mică decât 70%.