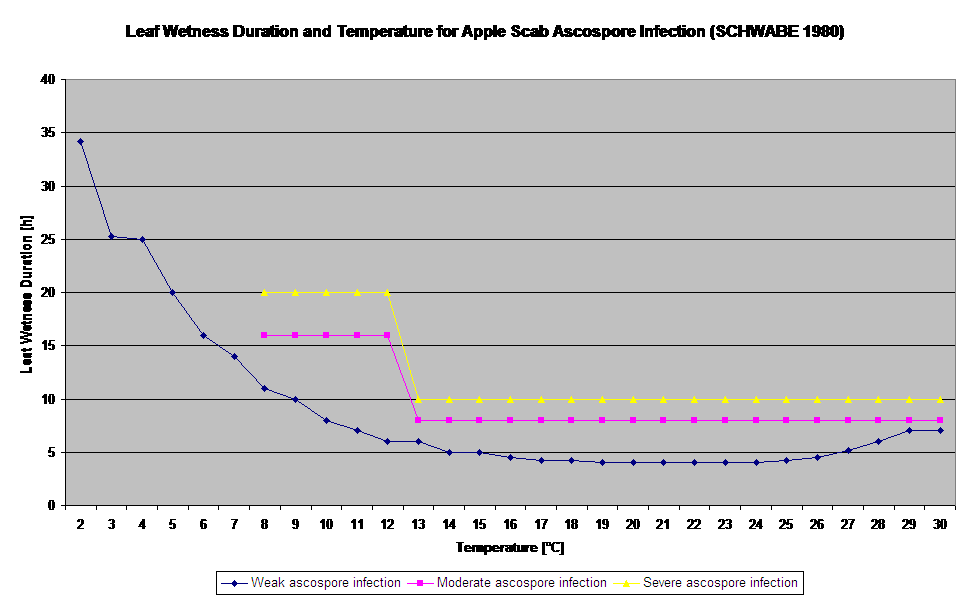
A germinação e penetração dos ascósporos de Venturia inaequalis em folhas ou frutos dependem dos períodos de humidade das folhas (folhas molhadas ou humidade rel. >80% (mantém o nível de infecção até abaixo de 80% r.h.). e temperatura do ar (acima de 2°C). Temperaturas mais altas levam, em condições húmidas, a uma penetração mais rápida, enquanto que a temperaturas mais baixas são necessários períodos mais longos de humidade das folhas. Esta relação foi publicada pela primeira vez por MILLS e LAPLANTE (1945). O nosso cálculo é realizado com base nas publicações de SCHWABE (1980). SCHWABE descreveu a gravidade das infecções de sarna, dependendo da temperatura. Estas observações estão em congruência com as nossas próprias observações.
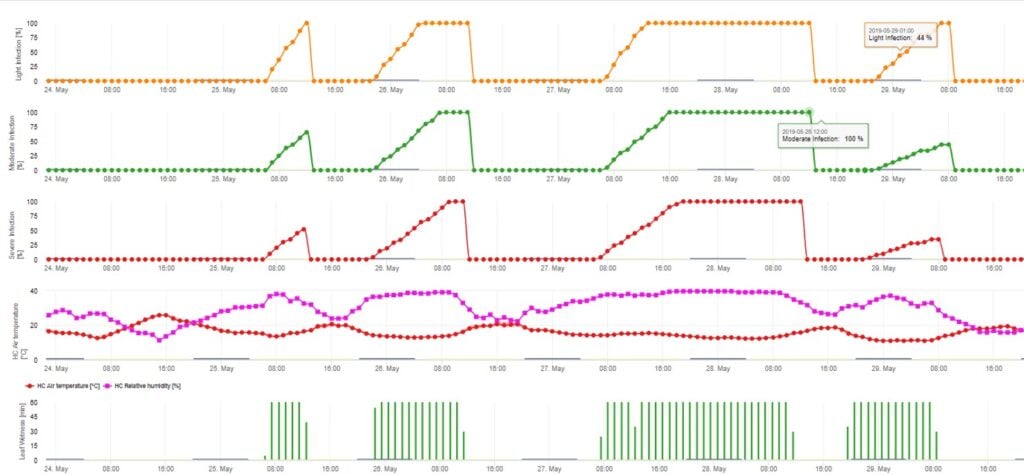
O Modelo de Infecção Ascospore calcula o progresso da infecção, dependendo da humidade das folhas e da temperatura, para infecções fracas, moderadas e graves. Dividir a previsão em três "classes de gravidade" é muito tradicional para modelos de sarna de maçã. O progresso da infecção é apresentado como um gráfico para valores horários. Uma infecção já ocorreu assim que a curva de progresso para uma infecção fraca atinge 100%. Uma infecção moderada ou grave é completa se as curvas de progresso para estas classes de gravidade estiverem a atingir (100%). Muitas vezes, na prática ou na realidade, uma infecção atinge 70% ou 80% no início da manhã e na dependência do tempo real, poderá saber se esta infecção terminará ou não. A apresentação dos valores máximos diários da curva do processo de infecção dá uma visão geral sobre as datas de infecção esperadas na estação.
Utilização principal do modelo
Para o mercado é absolutamente importante ter frutos de maçã sem doenças para obter um bom preço. Portanto, os cultivadores tendem a não ter qualquer tolerância à sarna nos seus pomares. O modelo deve ser implementado num programa de aplicação curativa (aplicação de sprays contendo compostos de ciprodinil ou pirimetanil no pré-bloom) quando o modelo apontava uma infecção (não importa se fraca ou grave). No período de floração e pós-bloom precoce, as infecções graves são mais frequentes e todos os períodos húmidos foram cobertos por sprays de protecção. Além disso, o tecido vegetal cresce muito rapidamente, pelo que uma protecção pode funcionar apenas durante 4 dias. Neste caso, uma pulverização curativa após um período de infecção incompleto pode também ser necessária. Mas neste caso, é relevante se tiver sido uma infecção fraca ou grave e também se houve um número baixo ou alto de ascósporos prontos para a infecção.
Literatura
- Schwabe WFS, 1980. Requisitos de humidade e temperatura para infecção das folhas de maçã por Venturia inaequalis na África do Sul. Phytophylactica, 69-0.
- Schwabe WFS, 1980. O tempo favorece a infecção da sarna da maçã na África do Sul. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Biologia, epidemiologia e gestão da sarna da maçã. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Infecções secundárias por conidia
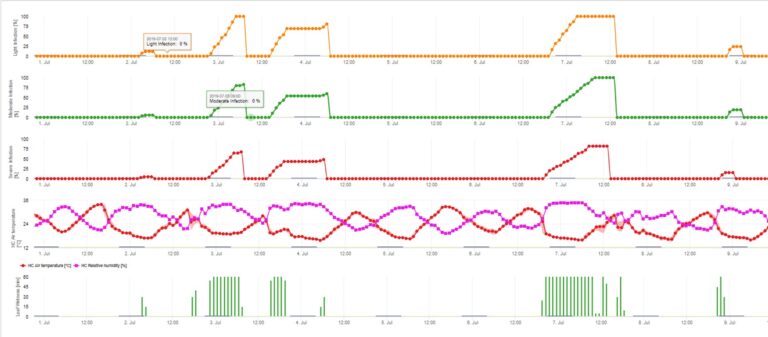
As infecções da sarna da maçã por conidia (assexual) dependem dos mesmos factores que as infecções causadas por ascósporos. A diferença é que as conidias podem ter alta em condições claras e escuras e as infecções conidiais ocorrem sobretudo durante a época de Verão.
Dependendo dos regimes de temperatura do ar, as conidias de Venturia inaequalis necessitam de períodos mais longos ou mais curtos de humidade foliar para germinação e penetração nas folhas ou frutos da macieira. Esta relação foi publicada pela primeira vez por MILLS e LAPLANTE (1945). Os nossos cálculos são efectuados com base nas publicações de SCHWABE (1980). Onde MILLOS e LAPLANTE (1945) assumiram apenas que as conidias necessitam de um certo período de tempo de humidade das folhas; SCHWABE (1980) também incluiu a importância da temperatura no processo de infecção.
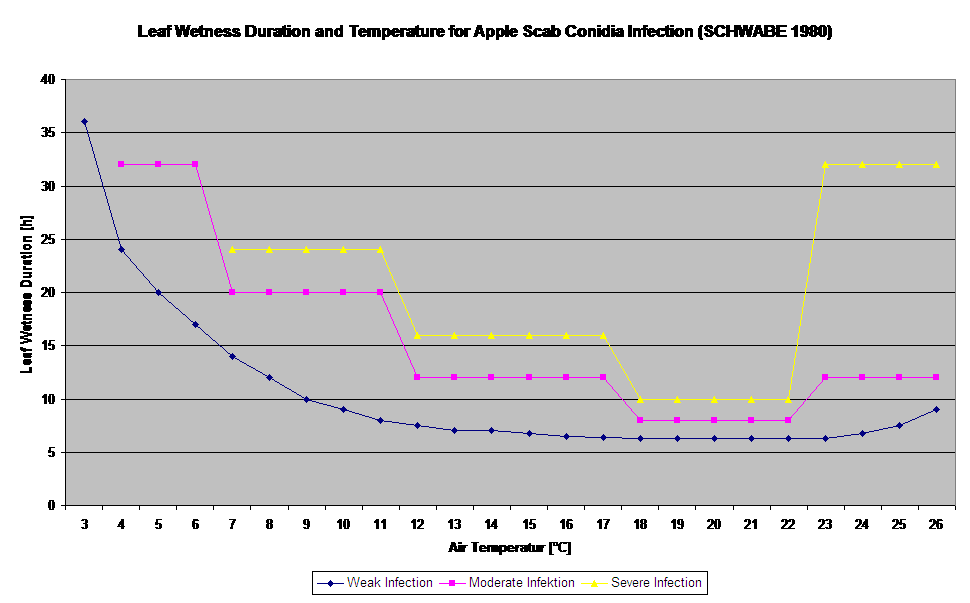
O Modelo de Infecção Conidia calcula o progresso da infecção dependendo da humidade das folhas e da temperatura para infecções fracas, moderadas e graves. Dividir a previsão em "três classes de gravidade" é muito tradicional para modelos de sarna de maçã. O progresso da infecção é apresentado como um gráfico para valores horários. Uma infecção já ocorreu assim que a curva de progresso para uma infecção fraca atinge 100%. Uma infecção moderada ou grave é completa se as curvas de progresso para estas classes de gravidade atingirem 100%. Muitas vezes, na prática ou na realidade, uma infecção atinge 70% ou 80% no início da manhã e em combinação com o tempo real poderá saber se esta infecção terminará ou não. A apresentação dos valores máximos diários da curva de progresso da infecção dá uma visão geral sobre as datas de infecção esperadas na estação.
Literatura:
- Infecção por Venturia inaequalis, indicando as datas de infecção por ascósporos e conídios (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Biologia, epidemiologia e gestão da sarna da maçã. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Apresentação gráfica do modelo
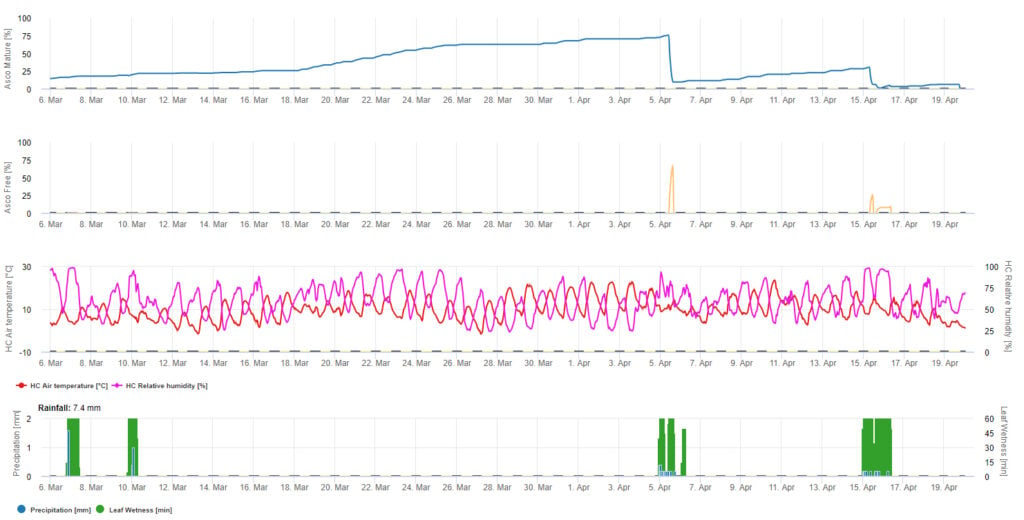
Para a apresentação gráfica da maturação Ascospore, são utilizados valores diários do modelo de formação e descarga. Mostra que os períodos de humidade das folhas levam à libertação de ascósporos e curvas para os ascósporos maduros e libertados. A gravidade da libertação e os números de ascósporos maduros não podem ser avaliados em valores absolutos. Os valores entre 0 e 100% indicam a quantidade relativa de ascósporos maduros e libertados em dependência das condições climáticas.
1. Maturação, formação e descarga de ascósporos:
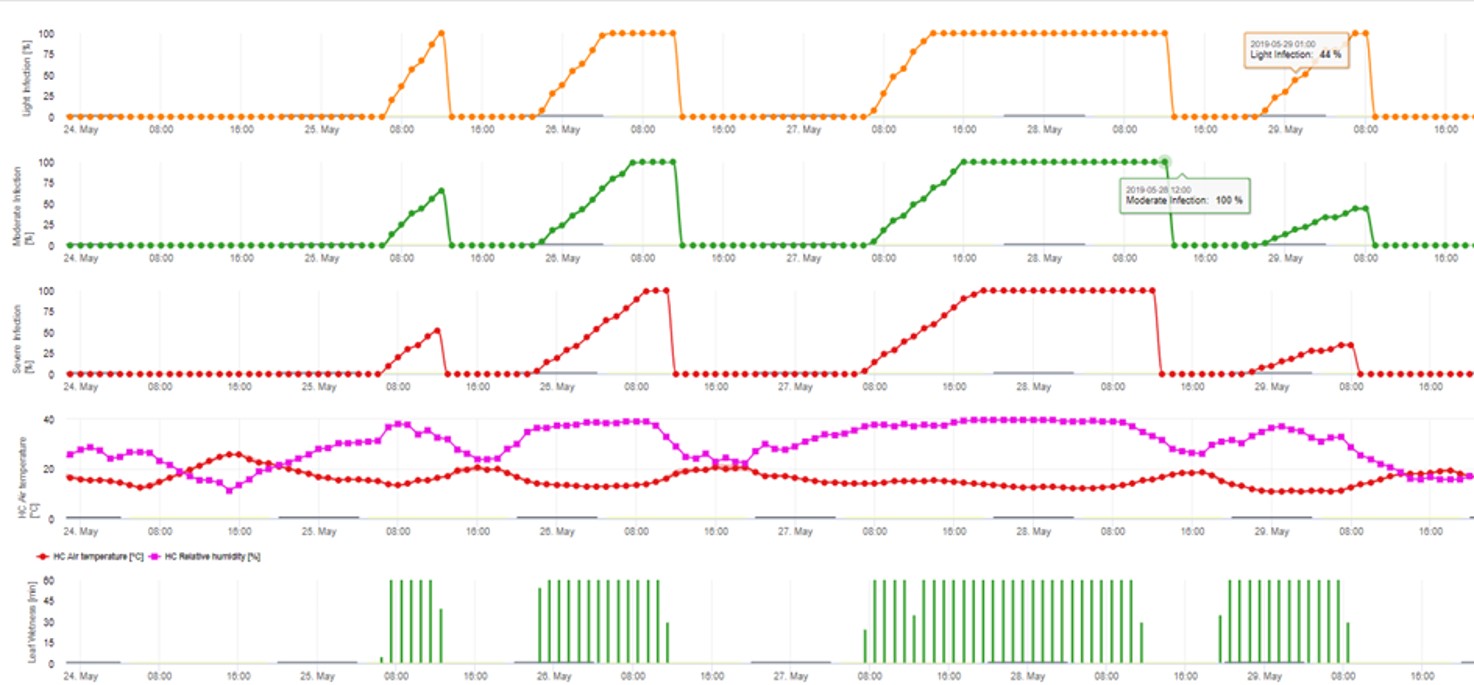
2. Infecção por Ascósporos mostrando em três classes de gravidade diferentes (fraca: laranja, moderada: verde, grave: vermelha). Sempre que a infecção pelo 100% foi atingida, foram determinadas as condições ideais para que o ascósporo infectasse o tecido vegetal. Assim, as medidas de protecção das plantas (antes da profilaxia da infecção, após o curativo da infecção 100%) têm de ser tidas em conta:
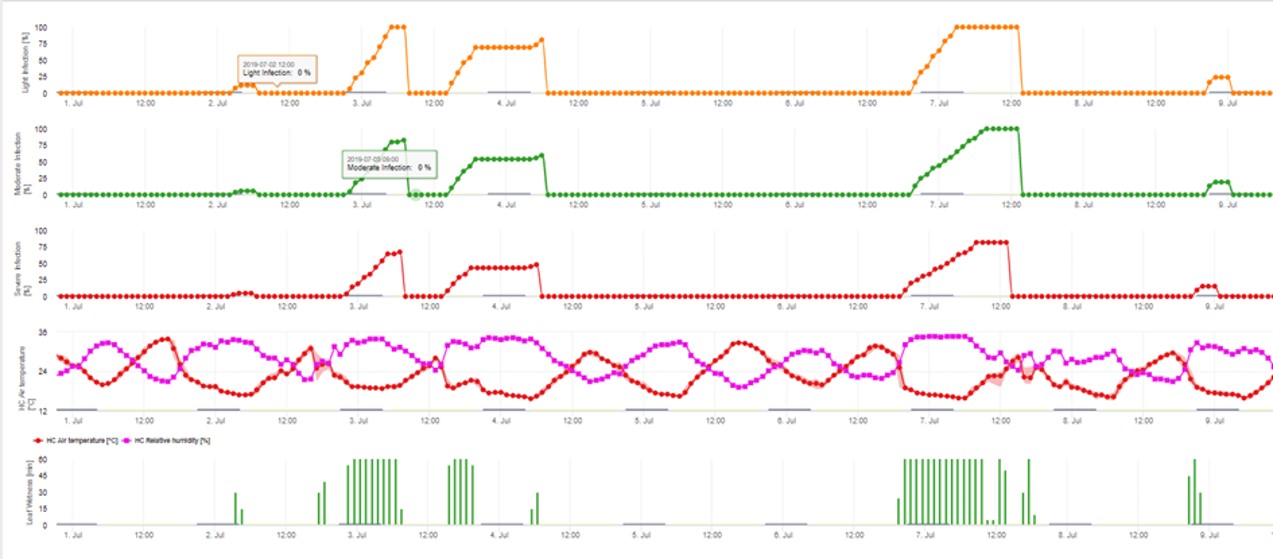
3. Conidia Infecção mostrando em três classes de gravidade diferentes (fraca: laranja, moderada: verde, grave: vermelha). Sempre que a infecção pelo 100% foi atingida, foram determinadas as condições ideais para que o ascósporo infectasse o tecido vegetal. Assim, as medidas de protecção das plantas (antes da profilaxia da infecção, após o curativo da infecção 100%) têm de ser tidas em conta:
As maçãs cultivadas comercialmente têm de estar livres de sarna. As maçãs com sarna só serão vendidas para processamento. Portanto, o objectivo de todas as actividades de protecção de plantas, tanto no cultivo convencional como no biológico, é ter frutos sem sarna. Os modelos, que mostram a descarga da sarna da maçã e a infecção por ascósporos/conídios são ferramentas muito importantes para atingir este objectivo.
Dois tipos básicos de fungicidas contra a sarna da maçã são utilizados em sistemas de cultivo convencionais: a) produtos preventivos como Captan, Mancozebe, Dithianone e Strobilurins ou b) produtos curativos como Cyprodinil (Chorus) ou Pyrimethanil (Scala) ou para aplicação nos últimos períodos mais quentes da estação, os fungicidas DMI. Na verdade, a maioria dos cultivadores segue uma estratégia preventiva. No entanto, uma estratégia preventiva prática não é capaz de proteger completamente as macieiras porque a macieira cresce e desenvolve flores, frutos e folhas. Portanto, o spray preventivo protege apenas durante um período de 4 a 7 dias em dependência do crescimento real da árvore. Estes intervalos de pulverização tão estreitos não são controláveis; por conseguinte, os cultivadores integrarão a sua experiência sobre o clima local, previsão do tempo e modelos de sarna da macieira na sua gestão de pulverização. Eles programarão as pulverizações preventivas com base na sua experiência e previsão do tempo. Os modelos de infecção da sarna da maçã irão mostrar-lhes a data exacta da infecção (fraca, moderada e grave), bem como o modelo de descarga ascósporo/conídio e, com a sua experiência, são capazes de estimar a importância de uma infecção. Isto dá a possibilidade de agir com um produto curativo se uma infecção da sarna da maçã foi demasiado tempo após o último spray preventivo.
Na produção biológica de maçã, o enxofre-calcário apontado como sendo o agente de controlo mais eficaz contra a sarna. O controlo óptimo pode ser alcançado se for pulverizado pouco antes da infecção ou no início da infecção. Isto tem de ser planeado com base na previsão do tempo. Um dia, falharemos este período óptimo e teremos de pulverizar nas folhas molhadas de uma infecção quase completa da sarna. Isto ainda dará uma boa eficácia. Os modelos da sarna da maçã estão a ajudar a decidir se é necessário um spray de emergência para uma infecção quase completa.
Sintomas de Fogo
Os cancros de Inverno, abrigando o agente patogénico do fogo, são muitas vezes claramente visíveis nos troncos e membros grandes como ligeiramente a áreas profundamente deprimidas de casca descolorada, que por vezes estão rachadas nas margens. O maior número de cancros, no entanto, é muito mais pequeno e não se distingue tão facilmente. Estes ocorrem em membros pequenos onde ocorreram infecções por florescimento ou rebentos no ano anterior e muitas vezes em torno de cortes feitos para remover membros com bolhas. Uma vez que muitos destes cancros são estabelecidos mais tarde na estação, não são frequentemente fortemente deprimidos e raramente apresentam fissuras na casca nas suas margens. Além disso, são frequentemente bastante pequenos, estendendo-se menos de 2,5 cm, com casca avermelhada a roxa que pode estar coberta com minúsculos corpos de frutificação de fungos pretos (mais notavelmente Botryosphaeria obtusao patogénico da podridão negra da maçã).
Os sintomas da floração aparecem mais frequentemente dentro de uma a duas semanas após a floração e geralmente envolvem todo o cacho de flores, que murcha e morre, ficando castanho na maçã e bastante preto na pêra. Quando o tempo é favorável ao desenvolvimento de agentes patogénicos, podem ser vistos glóbulos de exsudado bacteriano nas flores. O esporão com o aglomerado da flor também morre e a infecção pode alastrar e matar porções do membro de suporte. As pontas dos rebentos jovens infectados murcham, formando um sintoma muito característico de "tintureira do pastor". Os rebentos mais velhos que ficam infectados depois de terem sido desenvolvidos cerca de 20 folhas podem não mostrar este sintoma de ondulação na ponta. À medida que a infecção se espalha pelo eixo do rebento, as folhas mostram primeiro estrias escuras no meio das veias, depois murcham e tornam-se castanhas, permanecendo firmemente presas ao rebento durante toda a estação. Tal como nas infecções das flores, o patogéneo invade e mata frequentemente uma porção do membro que suporta o rebento infectado. O primeiro sintoma nos rebentos de água e rebentos que são invadidos sistemicamente a partir de cancros activos próximos é o desenvolvimento de uma descoloração amarela a laranja da ponta do rebento antes de ocorrer a murchidão. Além disso, os petíolos e veios médios das folhas basais desses rebentos tornam-se normalmente necróticos antes dos que se encontram na ponta do rebento.
Dependendo da cultivar e do seu estado de desenvolvimento no momento em que a infecção ocorre, uma única flor ou rebento pode resultar na morte de um membro inteiro, e onde o líder central ou tronco da árvore é invadido, uma grande parte da árvore pode ser morta em apenas uma estação. Em geral, as infecções de qualquer tipo que ocorrem entre a queda das pétalas e o conjunto de botões terminais levam geralmente à maior perda do membro e da árvore. Além disso, as árvores fortemente estruturadas tendem a sofrer perdas menos graves de membros do que as que são treinadas para sistemas mais fracos para alta produtividade. Quando os porta-enxertos de maçã altamente susceptíveis (M.26, M.9) ficam infectados, grande parte do tronco do enxerto e os membros principais acima da união do enxerto permanecem muito tipicamente menos sintomáticos, enquanto que se desenvolve um cancro castanho escuro distinto à volta do porta-enxerto. À medida que este porta-enxerto cilha a árvore, a porção superior mostra sintomas de declínio geral (má cor da folhagem, crescimento fraco) a meio ou fim da estação. Em alguns casos, a folhagem das árvores afectadas pelo míldio do porta-enxerto desenvolve-se de cor vermelha no final de Agosto a início de Setembro, não muito diferente da frequentemente associada à doença da podridão do colar causado por um fungo do solo. Algumas árvores com infecções de porta-enxertos podem não apresentar sintomas de declínio até à primavera seguinte, altura em que se podem ver cancros que se estendem para cima até ao tronco inferior.
Ciclo da doença
O agente patogénico bacteriano causador de queimaduras quase exclusivamente em cancros de membros infectados na estação anterior. O maior número de cancros e, portanto, os mais importantes na contribuição do inóculo, ocorre em membros de diâmetro inferior a 38 mm, especialmente em torno de cortes feitos no ano anterior para remover os membros com ferrugem. Durante o início da Primavera, em resposta às temperaturas mais quentes e ao rápido desenvolvimento dos botões, as bactérias nas margens do cancro começam a multiplicar-se rapidamente e produzem uma espessa gota amarelada a branca que é elaborada na superfície da casca até várias semanas antes do período de floração. Muitas espécies de insectos (predominantemente moscas) são atraídas pela exsudação, e subsequentemente dispersam as bactérias por todo o pomar. Quando as primeiras flores abertas são colonizadas pelas bactérias, os insectos polinizadores deslocam rapidamente o patogéneo para outras flores, iniciando mais floração. Estas flores colonizadas estão sujeitas a infecção dentro de minutos após qualquer evento de humedecimento causado pela chuva ou orvalho pesado quando as temperaturas médias diárias são iguais ou superiores a 16 °C, enquanto as pétalas das flores estão intactas (os recipientes das flores e os frutos jovens são resistentes após a queda das pétalas). Uma vez ocorridas as infecções das flores, podem esperar-se sintomas precoces com a acumulação de dias de pelo menos 57 graus (DD) superiores a 13 °C que, dependendo das temperaturas diárias, podem requerer 5 a 30 dias de calendário.
Com o aparecimento de sintomas de floração, o número e distribuição de fontes de inóculos no pomar aumenta muito. O inóculo proveniente destas fontes é ainda mais disseminado pelo vento, chuva, e muitos visitantes casuais de insectos a jovens pontas de rebentos, aumentando a probabilidade de um surto de flagelo de rebentos. Pesquisas recentes conduzidas na Pensilvânia indicam que a alimentação de pulgões não contribui para o abate de rebentos de pulgões. É necessária mais investigação para determinar se a cigarrinha desempenha ou não um papel na incidência do flagelo do rebento. A maioria das infecções de ponta de rebento ocorre entre o momento em que os rebentos têm cerca de nove a dez folhas e o conjunto de botões terminais, quando estão disponíveis fontes de inóculos e vectores de insectos, e temperaturas diárias médias de 16 °C ou mais.
Em anos em que não ocorrem infecções de floração, as fontes primárias de inóculo para a fase de floração são os cancros de Inverno e, em particular, os jovens rebentos de água perto destes cancros, que se infectam à medida que as bactérias se deslocam sistemicamente a partir das margens do cancro. Tais infecções sistémicas de rebentos, chamadas canker blight, são aparentemente iniciadas cerca de 111 DD superiores a 13 °C após a ponta verde, embora os sintomas visíveis possam não ser aparentes até à acumulação de pelo menos 157 DD superiores a 13 °C após a ponta verde. Na ausência de infecções de inflorescência, o desenvolvimento de infecções por cancros de rebentos é frequentemente localizado em redor de áreas com cancros de Inverno.
Floração de modelo
O modelo exige que o utilizador reconheça eventos e aspectos locais específicos e em constante mudança do seu pomar que podem aumentar ou diminuir o risco de incêndio em relação a outros pomares da região. O modelo exige que o utilizador assuma que existe um risco de infecção por fogo sempre que as flores estejam presentes nas árvores, especialmente durante a queda das pétalas e o período pós floração, quando as flores dispersas podem permanecer em muitas variedades de maçã e pêra. Pede-se ao utilizador do modelo que avalie cuidadosamente a situação no seu local específico e que inicie medidas de controlo se as flores estiverem presentes, se os níveis de risco forem "Elevados" ou "Extremos", e se for provável que a floração ocorra em algum momento durante as próximas 24 horas.
Estrutura do modelo: Temperaturas e Humidade: O processo chave de Fogo Blight que deve ser modelado é o potencial de crescimento bacteriano nos estigmas das flores de maçã e pêra. Este crescimento depende da temperatura, pelo que a previsão fiável do risco de infecção requer a utilização de um método de medição que reflicta com maior precisão o crescimento de Erwinia amylovora colónias. A principal discordância entre os modelistas é como isto deve ser feito.
O modelo Cougar blight estima a taxa de crescimento bacteriano com graus-horas com base numa curva específica de taxa de crescimento. Esta curva de crescimento é baseada na taxa de crescimento de E. amylovora bactérias em testes de laboratório. Os valores do grau hora são acumulados a cada hora do dia em que as temperaturas são superiores a 15 °C. Os valores horários aumentam à medida que as temperaturas aumentam de 15°C para 29°C, diminuem a temperaturas mais elevadas e chegam a zero para qualquer hora com temperaturas superiores a 40°C.
No FieldClimate.com ambos os modelos são exibidos na mesma figura. O modelo Fireblight DIV é baseado em Courgarblight e o modelo Blossomblight é baseado em Maryblight.
Puma modelo blight
A incidência de pragas de flores é um dos aspectos mais esporádicos das epidemias de pragas de fogo. O modelo foi construído no pressuposto de que existe uma abundância de inóculos e que, para que um evento de infecção por blossom blight ocorra, quatro condições estritas devem ser cumpridas em sequência. Estas condições são:
1) as flores devem ser abertas com estigmas e pétalas intactas, os estigmas têm de ser expostos para colonização, as flores em queda de pétalas são resistentes;
2) acumulação de pelo menos 110 °C horas > 18,3 °C nos últimos 44 °C dias > 4,4 °C define o potencial de infecção epífita para a flor mais antiga aberta e, portanto, mais colonizada do pomar
3) um evento de molhagem que ocorre como orvalho ou 0,2 mm de chuva ou 2,5 mm de chuva no dia anterior permite o movimento de bactérias dos estigmas colonizados para os néctarthodes
4) uma temperatura média diária de >= 15,6°C: Isto pode influenciar o ritmo a que as bactérias migram para os néctarthodes, bem como a multiplicação das bactérias necessárias para estabelecer infecções.
Apresentação gráfica dos modelos
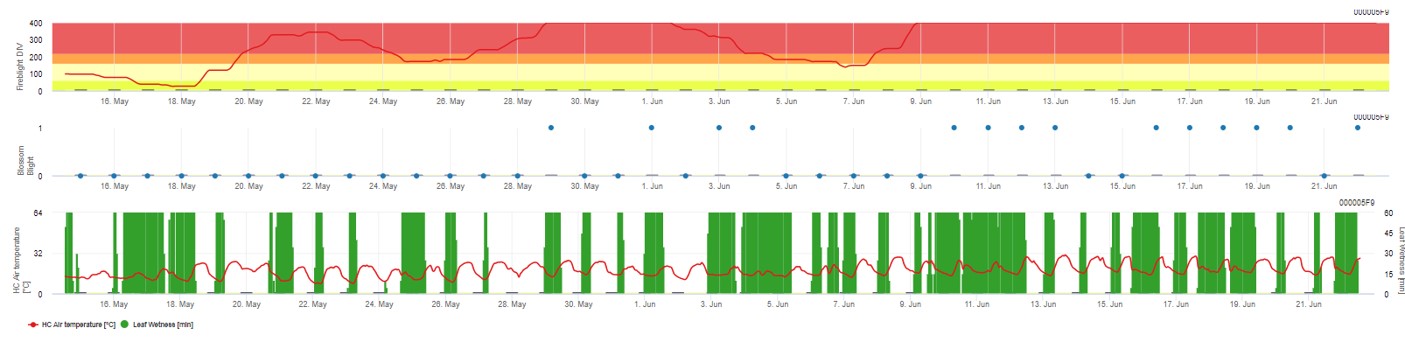
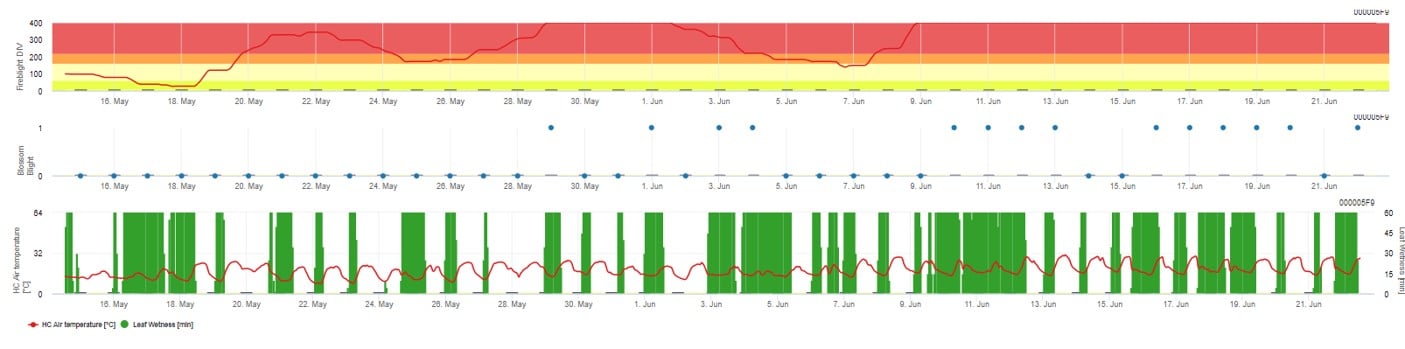
Ambos os modelos são exibidos na mesma página. A apresentação gráfica dos modelos Fire Blight é melhor para utilizar com valores agregados na base diária. O gráfico mostra os valores climáticos relacionados com esta doença. O resultado do Puma Blight chamado Fire Blight DIV e o resultado do Bloco de Fogo Blight. Para interpretar os resultados do Puma-Borrão, o gráfico é subposto por 5 cores diferentes. A distribuição destas cores é feita com base nas definições da primeira história da praga (ver botão no lado superior direito). As 5 cores indicam a classe de risco para os valores DIV.
Interpretação de valores (na dependência da ocorrência do flagelo do fogo nos últimos anos, criação básica):
Cankers activos presentes nas proximidades
Definir os parâmetros para a história da queimadura de fogo com o fundo azul "Settings" no lado superior direito:
Mostrando os resultados do modelo Fire Blight por 1) Modell Cougar Blight (chamado Fireblight DIV baseado na história e cores de fundo) e 2) Modell Blossom Blight
| Potencial para a presença de Pathogen | Baixo | Moderado | Alto | Extremo |
| Potencial para a presença de Pathogen | Baixo | Moderado | Alto | Extremo |
| Não houve fogo de artifício nos últimos dois anos | 0-350 | 350-500 | 500-800 | 800+ |
| Incêndio na área local em duas estações | 0-300 | 300-500 | 500-750 | 750+ |
| Incêndio na área local em duas estações | 0-250 | 250-450 | 450-700 | 700+ |
| Incêndio no pomar no ano passado | 0-200 | 200-350 | 350-500 | 500+ |
Limiar de acção:
O controlo é aconselhado se for detectado um período de infecção de alto ou extremo risco. Os factores que determinam os danos são: número de flores, idade da árvore, vigor e variedade,.... também.
A oportunidade de uma infecção por Blossom Blight é exibida por uma barra de 0 a 1 (para que todas as condições sejam ou não satisfeitas).
O objectivo dos modelos de Fogo Blight é avaliar a probabilidade de uma infecção por Erwinia amyloflora no pomar.
O modelo de Mary Blight que está a avaliar para o blossom blight indica muito bem situações de infecção de alto impacto económico. Por esta razão, é bastante utilizado para indicar o uso de antibióticos contra este agente patogénico. O puma está a dar informação sobre o risco que as infecções com o flagelo do fogo representam para as possibilidades globais de propagação do agente patogénico. A sua ponderação feita pela história de um pomar é muito útil para nos indicar quão cuidadosamente temos de verificar os pomares quanto aos sintomas do flagelo do fogo, mesmo em situações em que Mary Blight não indicará uma infecção.
Biologia da traça do Codling
O C. Pomonella passa o Inverno como uma larva madura fiada num casulo que se encontra em áreas protegidas na árvore (fendas ou fendas da casca), que é o local preferido, ou na base da árvore em ninhada moída. Nos pomares modernos de maçã de alta densidade há muito poucos locais de casulo nas árvores devido à casca lisa da árvore. Forma-se pupae no casulo girado pela larva madura.
A fase adulta emerge da caixa da pupa e após um curto período para endurecer o exoesqueleto voa para a árvore. A maior parte da literatura indica que a actividade da traça ocorre no terço superior da copa da árvore. Os adultos são activos ao anoitecer durante algumas horas com um limiar médio de actividade entre 13°C a 15°C. O acasalamento ocorre normalmente no hospedeiro e a postura dos ovos ocorre imediatamente a seguir. É possível, em noites quentes, que uma traça fêmea emerja, acasalar e pôr ovos num período de dois dias. A C. Pomonella A traça pode viver durante um período de 20 dias em condições ideais, no entanto, nos campos é provável que sobrevivam durante muito menos tempo, especialmente no Verão quente. Em condições de laboratório, uma traça fêmea põe até 100 ovos, mas a fecundidade real no campo é provavelmente menor e a maioria é posta nos primeiros dias após o aparecimento e acasalamento.
Os ovos são colocados individualmente na superfície superior da folha, nos galhos ou nos frutos. Na primeira geração, os ovos são postos apenas nas folhas até cerca de duas semanas depois de os frutos se terem depositado, quando a penugem dos frutos se desfaz. Na segunda geração, cerca de 65% dos ovos são postos nas folhas, mas estes são normalmente muito próximos de um fruto. Os ovos são brancos cremosos quando são postos pela primeira vez, têm um anel vermelho a meio do desenvolvimento e quando estão perto da eclosão pode ser vista a cápsula preta da cabeça.
Uma larva recém-eclodida vagueia até localizar um fruto. Ela mastiga através da pele e alimenta-se da polpa da maçã sob a pele durante alguns dias. No processo de mastigar através da pele e alimentar-se sob as larvas empurra a frass para fora da área de entrada. Existe frequentemente um anel vermelho à volta da entrada, especialmente nos frutos jovens. Depois de as larvas se molestarem até à fase seguinte, fura em direcção ao núcleo, onde se alimentará de sementes até atingir a maturidade. Quando quase madura, a larva fará um túnel desde o núcleo até ao exterior da maçã e tapará o buraco com frass (excremento) e seda. Quando pronta para rodar um casulo, a larva deixa o fruto, maravilhando-se através da árvore ou caindo ao chão num fio de seda, e encontra um local protegido.
Modelo para traça de Codling
O nosso modelo baseia-se na acumulação das temperaturas efectivas entre >= 10°C e <=32°C. No hemisfério norte, esta acumulação começa com o primeiro de Janeiro. No hemisfério Sul, começa no dia 1 de Julho. Acumulamos cada medida no seu equivalente para o dia (ver gráfico abaixo). Os seguintes limiares são utilizados para o desenvolvimento dos insectos:
*Geração Completa= 650
| *Colocação de ovos na Escotilha de Ovos | = 88 |
| *Primeira fase das larvas | = 60 |
| *Segundo Estágio de Larvas | = 65 |
| *Terceira fase de larvas | = 55 |
| *Para a fase das larvas | = 45 |
| *Cinco estádio Larvas mais Pupae | = 335 |
O gráfico produzido pelo modelo mostra a primeira aparência e duração do instar larvar 1 a 5 e a presença de adultos em 5 níveis.
| *Nível 1: | Actividade de voo, colocação não de ovos |
| *Nível 2: | Actividade de voo com possível postura de ovos. |
| *Nível 3: | Actividade de voo com alguma postura de ovos. |
| *Nível 4: | Actividade de voo com postura razoável a forte de ovos. |
| *Nível 5: | Actividade de voo com forte postura de ovos. |
Apresentação das fases adultas da Traça Codificadora em 5 níveis diferentes (primeiro gráfico) assim como os graus-dia acumulados (linha vermelha, segundo gráfico) para o desenvolvimento adulto e as fases larvares mostradas a azul (terceiro gráfico).
Várias espécies de afídeos estão presentes em pomares de maçãs. O pulgão de maçã verde, o pulgão de maçã aveia ou pulgão de erva de maçã e o pulgão de maçã rosada. Nos jardins privados, sobretudo, os pulgões de maçã rosada têm alguma importância. Este pulgão leva a fortes alterações hormonais nas pontas dos rebentos com infestações. As folhas tornam-se enroladas e o grito torna-se curto. Mais tarde, as folhas tornam-se amarelas e morrem. Este pulgão está a afectar sobretudo os rebentos jovens. O pulgão de maçã rosada pode produzir danos graves já numa pequena abundância. Enquanto que o pulgão de maçã verde e o pulgão de erva de maçã podem ocorrer em maior abundância sem levar a danos económicos. O pulgão de erva de maçã tem de ser classificado como não tendo qualquer importância económica.
Pulgão de maçã rosada
Esta é uma antiga espécie europeia introduzida nos Estados Unidos por volta de 1870. Tornou-se uma grande praga de maçãs perto do final do século XIX. Para prosperar, esta espécie deve ter uma abundância da sua planta hospedeira de verão, a plátano de folhas estreitas. Existe um paralelismo notável entre a introdução e a propagação do pulgão de maçã rosada e a plátano de folhas estreitas. Este pulgão ocorre em todas as zonas frutícolas do mundo.
Fases da vida
Ovo: O ovo é oval e ligeiramente achatado no lado ao lado da casca. O comprimento varia de 0,49 a 0,56 mm. Quando colocado pela primeira vez, é amarelo vivo e coberto com uma substância glutinosa que endurece com a idade. A cor muda gradualmente para amarelo esverdeado e finalmente para um jacto preto brilhante. O tempo necessário para esta mudança de cor varia, em condições normais de exterior, de cerca de nove dias a mais de duas semanas.
Ninfa: Os indivíduos que eclodem dos ovos são todos fêmeas sem asas vivíparas. Existem cinco instars. O último é a mãe de caule madura, que pouco depois do quarto instar, começa a produzir jovens vivas partenogeneticamente. Ela produz uma média de cinco a seis crias por dia. As ninfas da segunda geração, todas elas fêmeas, atingem a maturidade em duas a três semanas. A grande maioria das ninfas começa a reproduzir-se na maçã, embora algumas possam desenvolver asas e migrar para a plátano.
A terceira geração é produzido em Junho e início de Julho. A maioria desta geração desenvolve asas e migra para o plátano de folhas estreitas. Em algumas estações, as fêmeas sem asas da terceira geração produzem uma quarta geração de maçã. Nos últimos anos, tem sido observado em algumas áreas que populações prejudiciais de pulgão de maçã rosada têm persistido em pomares até ao meio do Verão ou mais tarde. Não se sabe se esta mudança se deve ao processo de selecção, a mudanças de hábito ou à falta de necessidade de um hospedeiro alternativo.
Adulto: O adulto varia consideravelmente nas marcações de cor. A cor geral é castanho rosado, com um molde rosado devido a uma cobertura pulverulenta. Alguns dos adultos mais velhos são roxos, enquanto que os adultos mais jovens são decididamente rosados-avermelhados.
Gama de hospedeiros
A maçã é o hospedeiro preferido, mas o pulgão também se alimenta de pêra e espinheiro. Cortland, Golden Delicious, Rhode Island Greening e Ida Red são todas variedades de maçã particularmente susceptíveis. O pulgão encontra-se em todas as áreas de fruticultura dos Estados Unidos e Canadá.
Lesão ou Dano: A alimentação com pulgão de maçã rosada provoca frequentemente o enrolamento das folhas de maçã, começando na queda das pétalas. Estas folhas podem mais tarde ficar vermelho vivo. A alimentação das folhas em redor dos cachos de frutos resulta frequentemente em cacho, atrofiamento e malformação dos frutos em desenvolvimento. Estas anomalias tornam-se piores à medida que os frutos se desenvolvem e podem eventualmente tornar os frutos insaldáveis.
Grandes populações de pulgões podem produzir grandes quantidades de melada como resíduo da seiva sobre a qual se alimentam. O mel de melada excretado na fruta servirá de solo de crescimento para fungos de bolor fuliginoso, o que afectará o acabamento da maçã. As toxinas na saliva dos afídeos também servem como uma "gota stop", impedindo a abcisão dos frutos (separação natural da árvore) na colheita normal.
História de vida: O pulgão passa o Inverno na fase dos ovos. A eclosão ocorre no início da Primavera, cerca de uma semana a 10 dias mais tarde do que o pulgão de grão de maçã e aproximadamente ao mesmo tempo que o pulgão de maçã. Os ovos eclodem quando os botões começam a abrir na Primavera, ao longo de um período de duas semanas. Assim que eclodem, os jovens procuram os botões de abertura da maçã; parecem preferir os botões de fruta. Alimentam-se do exterior do botão da folha e dos cachos de botões de frutos até as folhas começarem a desabrochar. Depois trabalham no interior dos cachos e começam a sugar a seiva dos caules e dos frutos recém-formados.
A sua alimentação provoca o enrolamento das folhas, protegendo os afídeos de pulverizações e alguns inimigos. O encaracolamento severo da folhagem provocado por esta espécie é provavelmente a característica mais característica do seu trabalho. Uma única mãe de caule localizada na parte inferior de uma folha perto da broca média fará com que a folha se dobre firmemente. São necessárias apenas algumas mães de caule para causar um encaracolamento severo de todas as folhas em torno de um botão floral aberto, proporcionando a protecção ideal aos afídeos em rápido desenvolvimento. As mães de caule atingem a maturidade quando as macieiras estão a entrar em floração.
As mães-tronco maduras são muito inactivas. Estabelecem-se e alimentam-se e produzem crias a um ritmo rápido. Quando distribuídas, retiram rapidamente os seus bicos dos tecidos vegetais e procuram outro local onde possam continuar. As mães de caule amadurecem cerca de duas semanas após a eclosão. A duração do tempo depende em grande parte das condições meteorológicas. A produção de crias começa normalmente dois ou três dias após a última muda e continua sem interrupção durante mais de um mês.
A produção total por uma única fêmea é de cerca de 185. Normalmente, o período de reprodução estende-se de cerca do início de Maio a Junho. Normalmente, o período máximo de actividade reprodutiva é por volta da última semana de Maio e da primeira semana de Junho, quando os frutos jovens começam a fixar-se e a iniciar o crescimento activo. O pulgão de maçã rosada raramente é encontrado atacando os rebentos jovens e de crescimento rápido. Restringe-se à folhagem, aos caules das flores e aos frutos jovens.
Uma das características desta espécie é a congregação das crias sobre a mãe. Cada mãe de caule individual ou grupo de mães terá reunido sobre ela centenas de crias. As folhas infestadas poderão em breve ser cobertas - nalguns casos, por mais de uma camada de afídeos. Este hábito de congregação mata em breve as folhas infestadas e provoca a migração forçada dos afídeos. As crias movem-se activamente e apressadamente, aparentemente ansiosas por localizar um local de alimentação adequado. São frequentemente encontrados durante este período reunidos nos frutos em formação ou atacando a nova e suculenta folhagem em desdobramento.
A segunda geração precisou de quatro a 40 dias para atingir a maturidade e produzir jovens. A maioria da segunda geração é de fêmeas sem asas. A produção média total de cada indivíduo na segunda geração é de cerca de 119. Os hábitos e actividades da terceira geração não diferem dos da segunda. Os afídeos reúnem-se em imenso número na parte inferior da folhagem, causando um encaracolamento severo. Também atacam o ambiente e desenvolvem frutos, produzindo lesões características.
A maioria desta terceira geração adquire asas e migra para a planície de folhas estreitas, a planta hospedeira de Verão. Uma quarta geração pode ser produzida. Todas estas produzem asas e migram para a banana-plátano de folhas estreitas. Após a última muda, os adultos com asas são muito tenros e inactivos. Permanecem segregados nas folhas enroladas durante dois ou três dias antes de se aventurarem no seu voo migratório. Pouco antes de voarem, tornam-se muito activos e nervosos, correndo ou movendo as suas asas para cima e para baixo em antecipação ao seu voo.
Pulgão de maçã verde
O pulgão de maçã verde, Aphis pomi (DeGeer), está amplamente distribuído em todas as áreas de cultivo de maçã. No hemisfério norte, aparece pela primeira vez em pomares de maçãs em finais de Maio a princípios de Junho. O insecto suga a seiva das folhas nos rebentos de água e no crescimento suculento terminal. Os pulgões de maçã verde são geralmente encontrados perto das veias principais na superfície inferior da folha. O pulgão da maçã verde também ataca pêra, espinheiro, marmelo, caranguejo e espiraea.
Descrição do pulgão de maçã verde
Os ovos do pulgão de maçã verde são ovais e negros brilhantes. As ninfas e os adultos são de cor verde escuro ou verde claro. O pulgão de maçã verde tem cabeça verde, abdómen verde uniforme ou verde amarelado, pernas pretas e longos cornicles pretos - os "tubos da cauda" (na extremidade posterior do insecto). As fêmeas adultas são de verde escuro a preto e com asas. Durante o Verão estas fêmeas, ou alates, dão à luz jovens vivas partenogenicamente.
Biologia do pulgão de maçã verde
O pulgão de maçã verde sobrepõe-se como um ovo em ventosas, na base de gomos e em cicatrizes foliares de rebentos terminais. Os ovos eclodem normalmente quando os botões rebentam e as primeiras folhas se desdobram. As ninfas começam a alimentar-se imediatamente sobre as folhas em desenvolvimento. Estão inicialmente presentes no rebento terminal e mais tarde deslocam-se para folhas mais velhas de aglomerado. Os afídeos fêmeas adultas produzem fêmeas sem asas em duas semanas, sendo cada fêmea capaz de produzir 50-100 crias vivas. Os afídeos jovens desenvolvem-se em sete a dez dias. As populações de afídeos verdes de maçã costumam desenvolver-se lentamente no início da Primavera (floração, pétalas, queda), e mais rapidamente à medida que as temperaturas médias diárias aumentam. Pulgões alados, ou alates, e pulgões sem asas são produzidos durante o Verão. Os afídeos de maçã verde são mais numerosos durante Julho e início de Agosto. Há mais ninfas que adultos alazados nas fases iniciais de uma infestação. A percentagem de alates aumenta à medida que a população cresce. As colónias de afídeos de maçã verde podem duplicar numa semana, se as condições forem favoráveis. Se os afídeos não tiverem um novo crescimento suculento e tiverem de se alimentar de folhas mais velhas, então o número de jovens produzidos pode descer até 50%. Se as temperaturas diárias forem de 30-32°C e superiores, as fêmeas não se reproduzem bem. Quando as temperaturas são altas durante vários dias, os afídeos morrem. Várias gerações do pulgão de maçã verde desenvolvem-se durante o verão, e as formas aladas dispersam-se pelo pomar. Por vezes o pulgão de maçã verde e o pulgão de maçã rosada, Dysaphis plantaginea Pars., têm colónias na mesma folha. O pulgão de maçã rosada é rosa ou púrpura. No Outono, machos e fêmeas sem asas desenvolvem-se e acasalam, e as fêmeas põem ovos para o Inverno. O maior número de ovos é de 15-20cm das pontas dos galhos. Os ovos raramente são colocados nos grandes membros do andaime ou nos troncos das macieiras.
Danos
O pulgão de maçã verde suga a seiva da folha. Infestações pesadas reduzem o vigor e o crescimento de rebentos. Isto é particularmente preocupante em viveiros e pomares jovens e não portadores. O insecto pode reduzir o tamanho dos botões e o comprimento dos entrenós, e causar o enrolamento das folhas. A alimentação com pulgões pode também estimular o crescimento de ramos laterais que podem afectar a forma das árvores. O enrolamento das folhas e os terminais enfraquecidos são susceptíveis a lesões no Inverno. O mel produzido pelos afídeos pode pingar nos frutos, permitindo o crescimento de fungos fuliginosos. Os fungos mancham os frutos e baixam o valor de mercado. Quando as infestações são pesadas, o pulgão de maçã verde pode alimentar-se de maçãs imaturas e causar carepa.
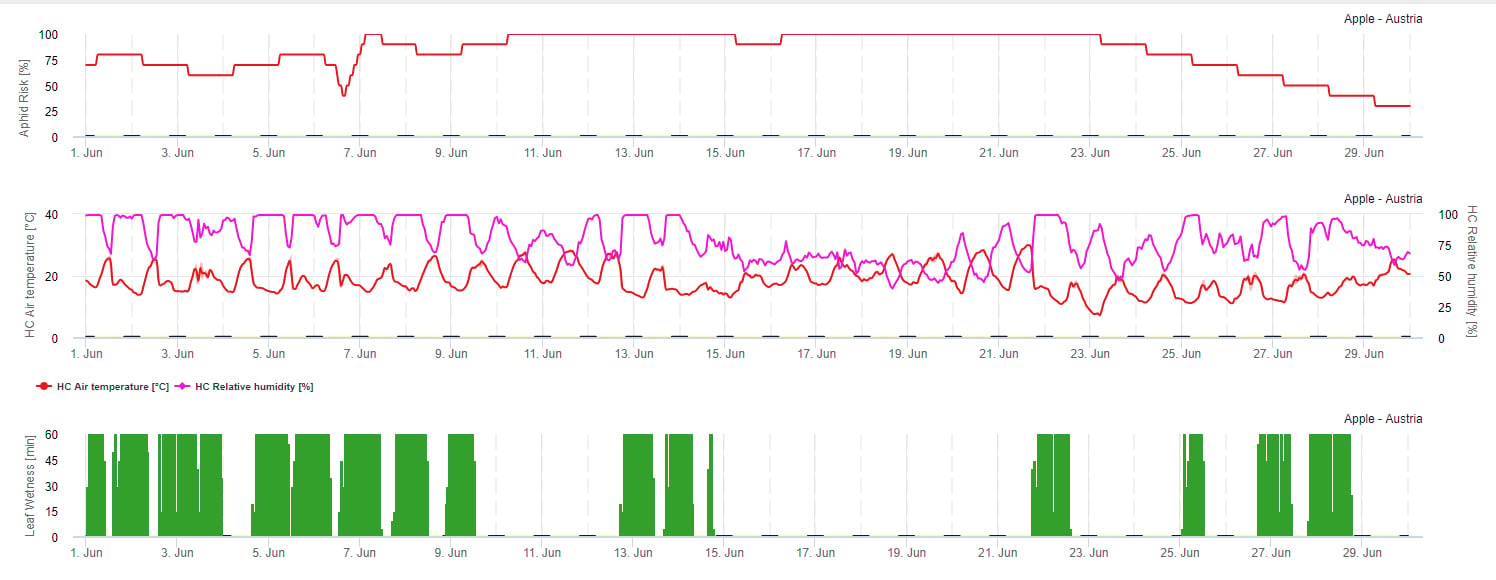
Modelo de risco de propagação
A propagação do afídeo é principalmente influenciada pela temperatura. Como mais horas se aproximam da temperatura óptima (20-32°C) e existe uma fonte alimentar disponível, uma vez que mais elevada será a taxa de propagação. Enquanto que os ovos e os instares jovens são susceptíveis contra o ar seco. A baixa humidade relativa pode danificar também as populações de pulgões (abaixo de 30% r.h.). Para além dos insecticidas, os afídeos são controlados por predadores e fungos parasitas. Os fungos parasíticos dependem da humidade. A maioria desta espécie como Beauvaria bassiana são favorecidos pela humidade livre.
Aumentamos o risco quando a temperatura óptima está entre 20 e 32°C e a humidade relativa entre 30 e 95% h. e diminuímos o risco quando está demasiado húmida (humidade foliar, chuva, ou humidade relativa acima de 95%), demasiado fria (temperaturas inferiores a 20°C) ou demasiado quente (temperaturas acima de 32°C), noites húmidas (humidade foliar em horas nocturnas).