Rdza soi, a konkretnie azjatycka rdza soi jest poważną chorobą soi wywoływaną przez patogen grzybowy Phakopsora pachyrhizi. Rozmieszczenie choroby: Azja i Australia, a ostatnio z Afryki, Ameryki Północnej i Południowej. Choroba została po raz pierwszy zaobserwowana w Japonii w 1902 roku. Patogen został rozprowadzony w całej Azji i Australii przez Amerykę. Rdza soi nie może przezimować w rejonach, gdzie panują mrozy, ale może szybko rozprzestrzeniać się przez wiatr na tak duże odległości, jej rozwój może być tak wybuchowy i może powodować tak szybką utratę liści, że obecnie jest to jedna z najbardziej obawiających się chorób w rejonach uprawy soi na świecie.
Objawy i oznaki
Pierwsze objawy rdzy soi powodowanej przez Phakopsora pachyrhizi zaczynają się jako bardzo małe brązowe lub ceglastoczerwone plamy na liściach. W warunkach polowych plamy te pojawiają się zwykle w dolnych partiach liści w okresie kwitnienia lub po jego zakończeniu, choć w pewnych okolicznościach może dojść do zakażenia siewek. Często pierwsze zmiany pojawiają się u podstawy liścia w pobliżu ogonka i żyłek liściowych. Ta część liścia prawdopodobnie dłużej zachowuje rosę, co stwarza warunki bardziej sprzyjające infekcji. Zmiany pozostają małe (2-5 mm średnicy), ale w miarę postępu choroby ich liczba rośnie. W tych zmianach, głównie na dolnej powierzchni liścia, tworzą się krostki, zwane urediniami, które mogą wytwarzać wiele urediniospor.
Podniesione krostki mogą być widoczne nieuzbrojonym okiem, zwłaszcza podczas sporulacji. Mimo że zmiany są małe, każda zmiana ma często kilka krost (uredinia) . Zmiany mogą być całkowicie pokryte urediniosporami, gdy krosty są aktywne. Urediniospory rdzy sojowej są bladożółto-brązowe do bezbarwnych, z echinulacyjnym (krótkie kolce) ornamentem powierzchniowym. Takie ubarwienie różni się od wielu innych patogenów rdzy, których zarodniki są często czerwonobrunatne (zabarwione rdzą). Kiełkowanie P. pachyrhizi urediniospory powstają przez równikowy (centralny) por, wytwarzając rurkę zarodkową zakończoną appressorium, którą grzyb wykorzystuje do wniknięcia do gospodarza bezpośrednio lub przez stomię.
W miarę jak na liściu tworzy się coraz więcej zmian, zaatakowane miejsce zaczyna żółknąć, a w końcu liść opada z rośliny. Rdza soi zwykle zaczyna się w dolnej części łanu, ale szybko postępuje w górę rośliny, aż wszystkie liście są w pewnym stopniu zaatakowane. Silnie chore rośliny mogą zostać całkowicie zdeformowane. Utrata efektywnej tkanki liściowej skutkuje obniżeniem plonów zarówno z powodu mniejszej liczby, jak i mniejszych nasion. Straty w plonie sięgają od 30 do 80%, ale wielkość strat zależy od tego, kiedy choroba się zaczyna i jak szybko postępuje. Oprócz liści, rdza soi może pojawiać się także na ogonkach liściowych, łodygach, a nawet liścieniach, ale większość zmian rdzy występuje na liściach.
Zmiany mogą być brązowe lub czerwono-brązowe. Zmiany opalenizny mają wiele krost, które produkują liczne urediniospory. Zmiany czerwono-brązowe, uważane za reakcję umiarkowanej odporności, mają tylko kilka krost, które produkują tylko kilka urediniosporów. Jak zostanie to omówione w części dotyczącej zwalczania choroby, ten typ zmian zależy od szczepu patogenu i może pojawić się na tym samym liściu ze zmianami opalenizny lub zmiany opalenizny mogą z wiekiem zmienić się w czerwono-brązowe. Objawy i oznaki na innych żywicielach, takich jak kudzu, są podobne, chociaż wielkość zmian może się różnić. Z wiekiem krosty mogą stać się czarne. Jest to spowodowane tworzeniem się warstwy teliospor w krostach, co powoduje przekształcenie krost z uredinii w telia.
Teliospory pełnią dwie funkcje: przetrwanie grzyba przy braku żywego gospodarza (overseasoning) oraz rozmnażanie płciowe. Grube ściany teliospor chronią grzyba przed środowiskiem i atakiem innych organizmów. U rdzy teliospory kiełkują tworząc basidium i cztery basidiospoory, podczas których dochodzi do rekombinacji płciowej. Kiełkowanie P. pachyrhizi teliospory obserwowano jedynie w laboratorium i nie wydaje się, aby w istotny sposób przyczyniały się do utrwalania tej choroby w terenie.
Biologia patogenów
Istnieją dwa blisko spokrewnione grzyby, które powodują rdzę na soi: Phakopsora pachyrhiziokreślany czasami jako azjatycki lub australijski patogen rdzy sojowej, który obecnie występuje również na półkuli zachodniej, oraz P. meibomiaeTak zwany patogen rdzy soi z Nowego Świata, który występuje tylko na półkuli zachodniej. Z wyjątkiem kilku drobnych cech, te dwa grzyby wydają się morfologicznie identyczne, ale P. pachyrhizi jest znacznie bardziej agresywny na soi niż P. meibomiae. Do tej pory, P. meibomiae nie udokumentowano, aby powodował znaczne straty w plonach w Ameryce Środkowej i Południowej. Te dwa gatunki można rozróżnić dzięki zastosowaniu protokołów analizy DNA.
Podobnie jak inne rdze, patogeny rdzy sojowej są obligatoryjnymi pasożytami, które wymagają żywego gospodarza do wzrostu i rozmnażania. W warunkach naturalnych mogą przetrwać z dala od żywiciela jako urediniospory tylko przez kilka dni. Oba patogeny rdzy soi, według naszej najlepszej wiedzy, wytwarzają tylko dwa rodzaje zarodników: urediniospory i teliospory (Rysunek 15). Stanowi to kontrast w stosunku do innych rdzy, które mogą mieć do pięciu stadiów rozwojowych zarodników (na przykład rdza źdźbłowa pszenicy). W przypadku rdzy sojowej, podobnie jak w przypadku większości rdzy, stadium urediniowe jest stadium powtarzającym się. Oznacza to, że urediniospory mogą infekować tego samego gospodarza, na którym zostały wytworzone (soję) w tym samym sezonie. Epidemia może szybko rozwinąć się z zaledwie kilku krost, ponieważ krostki wytwarzające zarodniki powstają już po 7-10 dniach od zakażenia, a każda krostka może wytworzyć setki urediniospor. W starych zmianach chorobowych wytwarzane są teliospory, ale w naturze nie wydają się one kiełkować, nie jest też znany żaden żywiciel zastępczy, ani aecia czy spermogonia.
Bez kiełkowania teliospor nie może nastąpić rozmnażanie płciowe. Brak rozmnażania płciowego powinien ograniczać zmienność grzyba rdzy, ale mimo to istnieje znaczna zmienność w P. pachyrhizi w odniesieniu do wirulencji. Ogranicza to stosowanie pojedynczych genów odporności w soi, ponieważ w krótkim czasie pojawiają się nowe izolaty patogenu, które pokonują gen odporności. Nie wiadomo, skąd bierze się ta zmienność w P. pachyrhizi. Rdza pasiasta pszenicy, Puccinia striiformis, ma podobny cykl życia jak P. pachyrhizi bez funkcjonującego stadium telialnego, a więc bez rozmnażania płciowego, ale ma wiele ras. Być może każdy gen odporności jest tak specyficzny, że pojedyncza mutacja we właściwym genie grzyba pozwala mu być zjadliwym na gospodarzach z nowym genem odporności.
Epidemiologia
Epidemia rdzy soi rozpoczyna się wraz z pojawieniem się inokulum przenoszonego drogą powietrzną (urediniospory). Patogen ten jest wyjątkowy wśród rdzy, ponieważ posiada wielu żywicieli alternatywnych, którzy mogą służyć jako źródła inokulum. Żywiciele alternatywni to inne rośliny, które mogą zostać zainfekowane przez ten sam patogen, ale nie są wymagane do zakończenia cyklu życiowego patogenu. Żywicieli alternatywnych nie należy mylić z żywicielami zastępczymi, czyli roślinami innymi niż żywiciel główny, które są potrzebne do zakończenia cyklu życiowego patogenu. Na obszarach wolnych od mrozu, takich jak Ameryka Południowa, Ameryka Środkowa, basen Morza Karaibskiego, południowy Teksas i Floryda, źródłem inokulum mogą być znajdujące się w pobliżu samosiewy soi, kudzu lub inny żywiciel alternatywny. W rejonach, w których występują mrozy, takich jak środkowo-zachodnie Stany Zjednoczone, inokulum musi być nawiewane ze źródeł zimujących, które mogą być oddalone o setki kilometrów.
Ponowne wprowadzenie patogenów obligatoryjnych do odległego regionu ma miejsce w przypadku kilku innych chorób, takich jak rdza źdźbłowa pszenicy i mączniak rzekomy, np. sina pleśń na tytoniu. Ponieważ zarodniki P. pachyrhizi są wrażliwe na promieniowanie ultrafioletowe, przemieszczanie się zarodników rdzy na duże odległości odbywa się prawdopodobnie w systemach burzowych, gdzie chmury chronią zarodniki przed słońcem. Gdy żywe zarodniki znajdą się na powierzchni liścia odpowiedniego gospodarza, infekcja i późniejszy rozwój epidemii zależą od warunków środowiskowych. Ogólnie rzecz biorąc, do zakażenia dochodzi, gdy liście są wilgotne, a temperatura wynosi od 8°C do 28°C, przy czym optymalna temperatura wynosi od 16°C do 28°C. W temperaturze 25°C do zakażenia dochodzi już po 6 godzinach od zwilżenia liści, ale optymalne jest 12 godzin. Po infekcji, w ciągu 7 lub 8 dni mogą pojawić się zmiany i krostki z urediniosporami i rozpoczyna się kolejny cykl infekcyjny.
Ten krótki cykl życiowy oznacza, że w odpowiednich warunkach epidemia rdzy soi może szybko narastać od prawie niewykrywalnego poziomu do bardzo wysokiego. Epidemia rdzy soi może przejść od poziomu poniżej wykrywalnego do defoliacji w ciągu miesiąca. Epidemia może wydawać się postępować nawet szybciej, ponieważ wczesne infekcje występują w dolnej części łanu i trudno je znaleźć. Poza środowiskiem, wiek roślin wpływa na epidemię rdzy soi. Zazwyczaj zmiany rdzy nie są znajdowane na soi do czasu kwitnienia, chyba że na początku sezonu występuje wysoki poziom inokulum. Może to być spowodowane większą podatnością roślin na rdzę, gdy gospodarz wchodzi w fazę reprodukcji, może to być spowodowane tym, że w niższych partiach łanu zarodniki są bardziej chronione przed promieniowaniem UV, lub może to być spowodowane tym, że warunki w łanie stają się bardziej wilgotne w miarę zamykania się łanu. W każdym razie zmiany chorobowe mogą powstawać w każdej fazie wzrostu, ale większe nasilenie chorób występuje dopiero po kwitnieniu.
W FieldClimate opisano i obliczono dwa modele dla rdzy soi:
Wzór 1: Głównymi czynnikami do obliczenia infekcji i nasilenia rdzy soi są wilgotność liści i temperatura (uwaga, infekcja musi wynosić 100%, aby można było obliczyć klasy nasilenia (od 1-3)) Liście muszą być wilgotne, a temperatura musi wynosić od 8 do 28°C - zaczyna się obliczanie infekcji (suma 7200 minut wynosi 100%). Jeśli infekcja wynosi 100% i w zależności od temperatury i czasu trwania zwilżenia liści, zaczynają być obliczane klasy nasilenia od 1- 3.
Model 2: Tutaj do obliczeń rdzy soi włącza się znacznie więcej czynników: Temperaturę, Wilgotność liści, Humidtiy względne, Opady atmosferyczne i promieniowanie słoneczne (hieny grzybów są w stanie wejść do aparatów szparkowych w ciągu dnia).
Gdy infekcja rdzy soi osiągnie 100% zaczyna się obliczać jej nasilenie. Jeśli obie wartości (infekcja i nasilenie) wynoszą 100%, przewiduje się wystąpienie (powstanie?) lokalnego inokulum.
Literatura:
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L., and Royer, M. H. 1989. Effects of duration, frequency, and temperature of leaf wetness periods on soybean rust. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte, and A.P. Dias. Poznanie ryzyka wystąpienia rdzy soi poprzez porównanie z Brazylią. Wydział Patologii Roślin, Uniwersytet Stanowy Iowa
- Yang, X. B. 1995. Ocena i zarządzanie ryzykiem wystąpienia rdzy soi. Proceeding of the soybean rust workshop, 9-11 August 1995. J. B. Sinclair i G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL
Zgnilizna twardzikowa lub biała pleśń soi to choroba wywoływana przez grzyb Sclerotinia sclerotiorum. Choroba ta może powodować znaczne obniżenie plonu nasion, a nawet całkowicie zniszczyć uprawę, gdy soja jest sadzona w porażonej glebie i występuje gęsty baldachim roślin z przedłużającymi się okresami wilgotnej pogody. Straty w plonie występują zazwyczaj, gdy występowanie choroby wynosi 15 % lub więcej. W większości lat w Północnej Dakocie zgnilizna twardzikowa łodyg stanowi niewielki lub umiarkowany problem dla hodowców, a choroba jest rzadko obserwowana w latach suszy. Mokra pogoda jest głównym czynnikiem rozwoju choroby. Stanowi ona uporczywy problem w przypadku nawadnianych upraw soi. Oprócz zmniejszenia plonu nasion, choroba powoduje również obniżenie jakości nasion i zanieczyszczenie nasion czarnymi sklerotami grzyba. Zanieczyszczenie nasion może być poważnym problemem w przypadku nasion eksportowanych, ponieważ może spowodować odrzucenie partii nasion w zagranicznych portach wejścia. Ponadto sklerocja zawrócona do gleby może mieć wpływ na inne uprawy w płodozmianie. Zgnilizna twardzikowa łodyg może być kontrolowana przez hodowców poprzez zrozumienie patogenu i cyklu chorobowego.
Symptomy
Objawy są zwykle obserwowane dopiero po zamknięciu się łanu między rzędami, co powoduje powstanie wilgotnego mikroklimatu. Zwykle pierwszymi obserwowanymi objawami są więdnięcie i usychanie liści, a następnie zamieranie roślin. Dokładna inspekcja pod baldachimem roślin ujawnia cottony, białe grzybnie (nici grzyba) na łodygach, liściach lub strąkach. Zmiany rozwijają się na głównych łodygach i bocznych gałęziach. Ostatecznie, zmiany opasują łodygi i części roślin powyżej umierają. Łodygi wydają się bielone, a czasem poszarpane od zaawansowanej zgnilizny. Z białej grzybni rosnącej na tkankach roślinnych powstają duże, czarne skleroty o różnych kształtach i rozmiarach. Sklerocja tworzą się również w rdzeniu łodygi i mają charakterystyczny cylindryczny kształt. Nasiona w chorych strąkach są zwykle wyschnięte i mogą być porażone przez grzyba lub zastąpione przez czarne sklerocja. Nasiona są zwykle zanieczyszczone sklerotiami podczas zbioru porażonych roślin.
Biologia patogenów
Grzyb ten ma rozległy zakres żywicieli obejmujący ponad 370 gatunków roślin i powoduje choroby na wielu różnych uprawach, takich jak słonecznik, sucha fasola, canola (rzepak), ziemniak, lucerna, gryka, łubin, gorczyca, topinambur, krokosz, soczewica, len, groch polny i wiele warzyw. W Północnej Dakocie rzadko zdarza się, aby patogen ten powodował poważne szkody w niektórych z tych upraw, takich jak len i ziemniak. Występuje również wiele pospolitych chwastów liściastych, takich jak bieluń dziędzierzawa, barwinek, oset kanadyjski i dzika gorczyca. Grzyb powodujący białą pleśń na soi jest tym samym, który powoduje białą pleśń lub chorobę Sclerotinia słonecznika, suchej fasoli, rzepaku i innych upraw. Sclerotinia sclerotiorum zimuje głównie jako sklerocja w glebie. Sklerocja kiełkują tworząc małe, brązowe grzyby zwane apotecjami (o średnicy od jednej ósmej do jednej czwartej cala). Wytwarzają one zarodniki zwane askosporami, które zapoczątkowują chorobę na soi i innych podatnych uprawach.
Cykl chorobowy
Wilgoć i kwitnienie są czynnikami krytycznymi dla rozwoju choroby. Choroby zwykle nie pojawiają się przed zamknięciem łanu, ponieważ gęsty łan sprzyja chłodnym temperaturom i wilgotnemu mikroklimatowi wokół łodyg oraz utrzymuje wysoką wilgotność gleby po deszczu lub nawadnianiu. Inicjacja choroby jest również ściśle związana z kwitnieniem. Po siedmiu do 14 dniach wysokiej wilgotności gleby, skleroty w górnej części kilku cali gleby kiełkują tworząc grzybopodobne apotecje. Pojedyncze sklerotium może wytworzyć kilka apotecji. Apotecja siłą wyrzuca swoje askospory do powietrza, gdzie są one przenoszone przez prądy powietrzne na rośliny soi. Najważniejszym źródłem askospor są apotecja wytwarzane na danym polu, ale askospory mogą być nawiewane z sąsiednich lub pobliskich pól. Jedno apothecium może wytwarzać ogromne ilości askospor w ciągu kilku dni. Askospory przez krótki czas utrzymują się na tkankach roślinnych, ale nie zimują. Do kiełkowania i wzrostu askospory wymagają warstwy wody i bazy pokarmowej, takiej jak obumarłe lub starzejące się tkanki kwiatów, zanim zainfekują roślinę. Tkanka kwiatowa jest najważniejszą bazą pokarmową do zainicjowania infekcji. Często infekcje rozpoczynają się w osiach łodyg, gdzie opadły i utkwiły opadłe tkanki kwiatowe. Do infekcji może dojść również poprzez rany spowodowane gradem lub innymi urazami. Warstwa wody na powierzchni rośliny sprzyja rozwojowi zmian chorobowych i zwiększa rozmiar uszkodzeń tkanek. Początkowy rozwój choroby wymaga zazwyczaj ponad 40 godzin ciągłego zwilżenia powierzchni rośliny, ale gdy choroba już się rozwinie, krótsze okresy zwilżenia umożliwiają rozwój zmian. Jest to powód, dla którego choroba jest związana z długimi okresami pochmurnej, wilgotnej i deszczowej pogody. Gdy powierzchnie roślin wysychają, postęp choroby jest spowolniony. Chłodne temperatury pomiędzy 59 a 75 stopni Fahrenheita sprzyjają rozwojowi choroby. Im większa gęstość łanu roślin, tym korzystniejsze warunki środowiskowe dla choroby. Dlatego też soja zasiana w całości i uprawy złożone są bardziej podatne na rozwój choroby. W miarę wzrostu grzybni w i na tkankach roślinnych powstają sklerocja. Te sklerocja nie będą kiełkować, aby utworzyć więcej apotecji w trakcie sezonu, ale raczej zostaną zwrócone do gleby podczas zbiorów i zabiegów uprawowych i przezimują, aby stać się inokulum (źródłem infekcyjnego grzyba) dla przyszłych podatnych upraw. Sklerocja są strukturami bardzo odpornymi i przetrwają w glebie przez długi czas.
Źródło: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
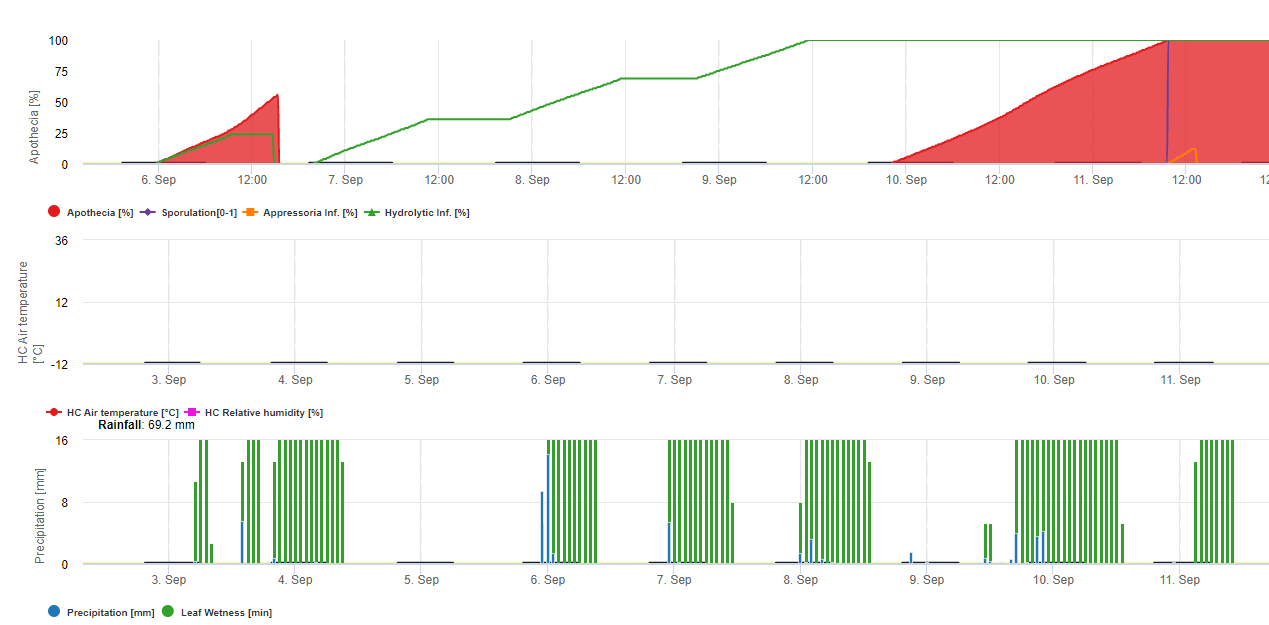
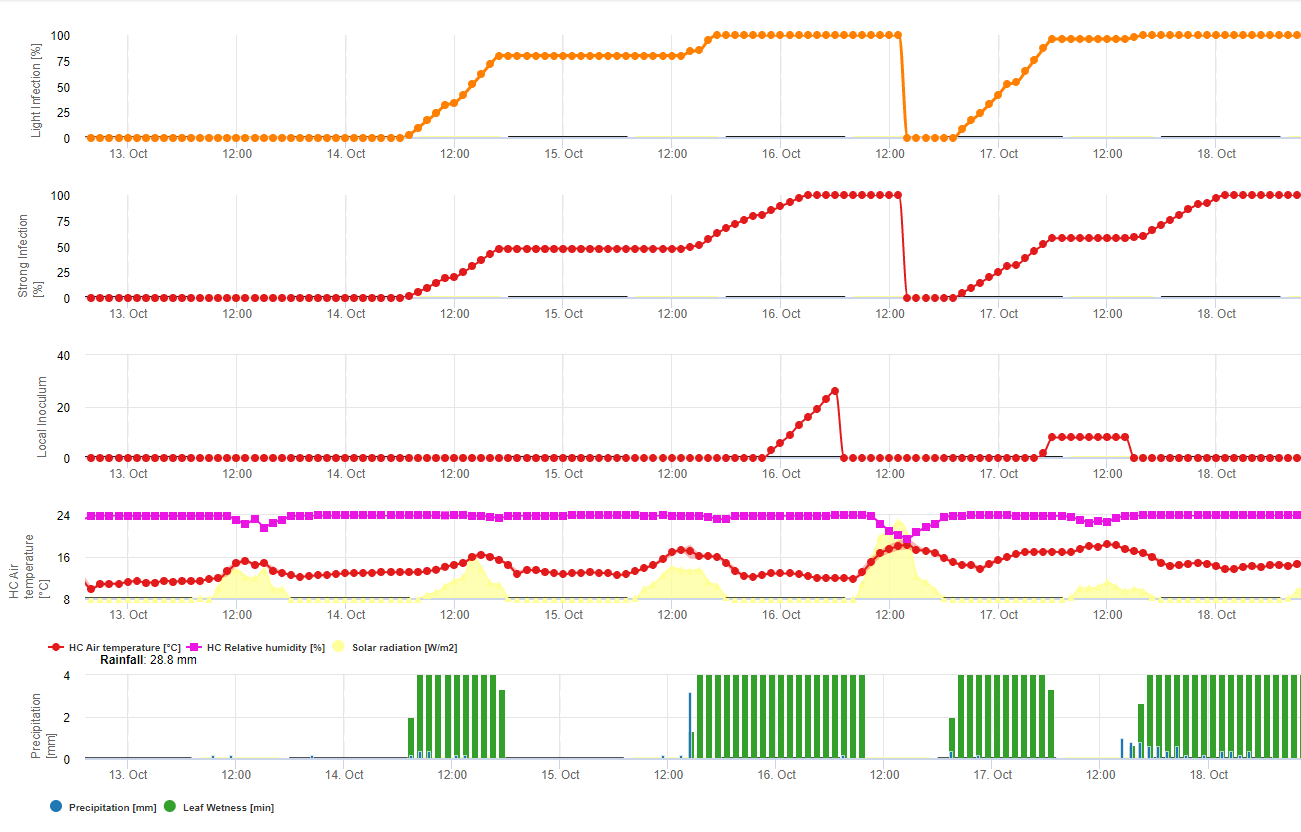
W FieldClimate model dla Sclerotinia jest obliczany w zależności od okresu deszczowego, wilgotności względnej i temperatury oraz wilgotności liści. Podczas długich okresów wilgotnych zalecana jest infekcja poprzez utworzenie apresorium przez patogen grzybowy. Innym sposobem infekcji jest tzw. infekcja hydrolityczna. Sposób ten polega na uwalnianiu enzymów hydrolitycznych, które kolejno degradują kutikulę roślinną, blaszki środkowe oraz pierwotne i wtórne ściany komórkowe i całą roślinę. Model opisany szczegółowo na tej stronie.