





Gruszka modele choroby
Kumulacja deszczu
Intensywny deszcz zmyje pestycydy pozostające na liściach winorośli lub innych roślin. Od 1980 roku nastąpiła duża poprawa odporności na deszcz nowoczesnych fungicydów. Właściwie możemy oczekiwać, że najnowocześniejsze fungicydy wytrzymają do 30 mm deszczu, jeśli miały szansę wyschnąć na liściu. Jeśli deszcz zaczynał się zaraz po oprysku lub w trakcie oprysku, odporność na deszcz może być znacznie zmniejszona.
W przypadku starych receptur fungicydów kontaktowych musimy spodziewać się twardości deszczu poniżej 12 mm. Tak jak byliśmy do tego przyzwyczajeni w latach 70-tych. Aby zmoczyć liście w winnicy potrzeba około 2 mm deszczu. Dlatego w tym mdoulu kumulujemy tylko deszcze większe niż 2 mm w jednym okresie zwilżenia liści. Oznacza to, że może być w sumie 6 mm deszczu w ciągu jednego dnia, ale ten moduł nie gromadzi żadnego z nich, ponieważ liście wyschły ponownie zanim spadło 2 mm deszczu.
Deszcz kumuluje się przez 3, 5 i 7 dni. W dłuższym okresie wzrost roślin ma znacznie większe znaczenie dla działania fungicydów kontaktowych niż odporność związków na deszcz.
Zaraza ogniowa
Źródło: P.W. Steiner, T. van der Zwet, and A. R. Biggs
Zaraza ogniowa jest niszczącą chorobą bakteryjną jabłek i gruszek, która powoduje zamieranie kwiatów, pędów, gałęzi, a niekiedy całych drzew. Choroba ta występuje w prawie wszystkich umiarkowanych i ciepłych rejonach uprawy jabłek na całym świecie. Chociaż ogniska choroby są zazwyczaj bardzo nieregularne, powodując w niektórych sadach w niektórych latach poważne straty, a w innych niewielkie lub nie powodując żadnych istotnych szkód. To nieregularne występowanie przypisuje się różnicom w dostępności zimującego inokulum, specyficznym wymaganiom regulującym infekcję, wahaniom w specyficznych lokalnych warunkach pogodowych oraz stadium rozwoju dostępnych odmian. Potencjał niszczący i sporadyczny charakter zarazy ogniowej, wraz z faktem, że epidemia często rozwija się w kilku różnych fazach, sprawiają, że zwalczanie tej choroby jest trudne i kosztowne.
Symptomy
Na pniach i dużych konarach często wyraźnie widoczne są zimujące kanarki, w których znajduje się patogen zarazy ogniowej, w postaci lekko lub głęboko zagłębionych obszarów przebarwionej kory, które czasami są popękane na brzegach. Najwięcej jest jednak kanarków znacznie mniejszych i nie tak łatwo je odróżnić. Występują one na małych kończynach, gdzie w poprzednim roku doszło do infekcji kwiatów lub pędów, a także często wokół cięć wykonanych w celu usunięcia porażonych kończyn. Ponieważ wiele z tych kanarków powstaje w późniejszym okresie sezonu, nie są one często silnie zagłębione i rzadko widać na ich brzegach spękania kory. Ponadto są one często dość małe, sięgające mniej niż 2,5 cm, z czerwonawą lub fioletową korą, która może być pokryta drobnymi, czarnymi owocnikami grzyba (przede wszystkim Botryosphaeria obtusa, patogen czarnej zgnilizny jabłek).
Objawy zarazy kwiatowej pojawiają się najczęściej w ciągu jednego do dwóch tygodni od zakwitnięcia i zazwyczaj obejmują całe grono kwiatowe, które więdnie i zamiera, brązowiejąc w przypadku jabłoni i całkiem czerniejąc w przypadku gruszy. Gdy pogoda sprzyja rozwojowi patogenu, na kwiatach widoczne są kuleczki bakteryjnej maści. Pęd, na którym znajduje się grono kwiatowe, również zamiera, a infekcja może rozprzestrzenić się na część gałęzi podporowej i ją zabić. Końcówki młodych zainfekowanych pędów więdną, tworząc bardzo typowy objaw "pastucha". Starsze pędy, które zostały zainfekowane po wykształceniu około 20 liści, mogą nie wykazywać tego objawu zwijania się końcówki. W miarę rozprzestrzeniania się infekcji w dół osi pędu, na liściach pojawiają się najpierw ciemne smugi w środkowych żyłkach, a następnie więdną i brązowieją, pozostając przez cały sezon ściśle przylegającymi do pędu. Podobnie jak w przypadku infekcji kwiatów, patogen często poraża i zabija część kończyny podtrzymującej porażony pęd. Pierwszym objawem na pędach wodnych i pędach, które zostały zaatakowane systemowo przez pobliskie aktywne kanarki, jest pojawienie się żółtego do pomarańczowego przebarwienia na końcówce pędu przed zwiędnięciem. Ponadto ogonki liściowe i środkowe żyłki liści podstawowych na takich pędach zwykle stają się nekrotyczne przed tymi na wierzchołku pędu.
W zależności od odmiany i jej stadium rozwoju w momencie wystąpienia infekcji, zakażenie pojedynczego kwiatu lub pędu może spowodować śmierć całej kończyny, a w przypadku zaatakowania centralnej części drzewa lub pnia, znaczna część drzewa może zostać zabita w ciągu jednego sezonu. Ogólnie rzecz biorąc, wszelkiego rodzaju infekcje występujące pomiędzy opadaniem płatków a zawiązaniem pąków końcowych prowadzą zazwyczaj do największych strat w kończynach i drzewach. Ponadto drzewa silnie ustrukturyzowane mają tendencję do mniej poważnych strat w kończynach niż drzewa przyuczone do słabszych systemów w celu uzyskania wysokiej wydajności. W przypadku zakażenia wysoce podatnych podkładek jabłoni (M.26, M.9), znaczna część pnia zraza i głównych gałęzi powyżej zawiązka szczepu zwykle nie wykazuje objawów, podczas gdy wokół podkładki rozwija się wyraźny ciemnobrązowy rak. Gdy rak podkładki opasuje drzewo, górna część wykazuje objawy ogólnego spadku (słaby kolor liści, słaby wzrost) w połowie lub pod koniec sezonu. W niektórych przypadkach liście drzew dotkniętych zarazą podkładki nabierają wczesnojesiennego czerwonego koloru pod koniec sierpnia lub na początku września, co nie jest podobne do tego, co często wiąże się z chorobą zgnilizny kołnierzykowej powodowanej przez grzyby glebowe. Niektóre drzewa z infekcją podkładki mogą nie wykazywać objawów spadku aż do następnej wiosny, kiedy to można zaobserwować kłykcie rozszerzające się ku górze w dolnej części pnia.
Cykl chorobowy
Patogen bakteryjny powodujący zarazę ogniową zimuje prawie wyłącznie w kankach na kończynach zainfekowanych w poprzednim sezonie. Najwięcej sęków, a więc i najwięcej inokulum, występuje na kończynach o średnicy mniejszej niż 38 mm, zwłaszcza w okolicach cięć wykonanych w poprzednim roku w celu usunięcia porażonych kończyn. Wczesną wiosną, w odpowiedzi na cieplejsze temperatury i szybki rozwój pąkówBakterie na brzegach raka zaczynają się szybko namnażać i wytwarzają gęstą, żółtawą do białej maź, która jest wypracowywana na powierzchni kory do kilku tygodni przed okresem kwitnienia. Muł jest przyciągany przez wiele gatunków owadów (głównie muchy), które roznoszą bakterie po całym sadzie. Gdy kilka pierwszych otwartych kwiatów zostanie skolonizowanych przez bakterie, owady zapylające szybko przenoszą patogen na inne kwiaty, zapoczątkowując w ten sposób większą liczbę przypadków zarazy kwiatowej. Te skolonizowane kwiaty są narażone na infekcję w ciągu kilku minut po tym, jak każdy zdarzenie zwilżenia spowodowany przez deszcz lub silna rosa gdy średnie temperatury dzienne są równe lub wyższe niż 16 °C przy nienaruszonych płatkach kwiatów (po opadnięciu płatków odporne są zawiązki kwiatów i młode owoce). Po zakażeniu kwiatów można spodziewać się wczesnych objawów z nagromadzeniem at. co najmniej 57 stopniodni (DD) wyższa niż 13 °C co w zależności od temperatur dobowych może wymagać od 5 do 30 dni kalendarzowych.
Wraz z pojawieniem się symptomów zarazy kwiatowej, liczba i rozmieszczenie źródła inokulum w sadzie znacznie wzrasta. Inokulum z tych źródeł jest dalej rozprzestrzeniane przez wiatr, deszcz i wielu przypadkowych gości z owadami do młodych wierzchołków pędów, zwiększając prawdopodobieństwo pojawienia się zarazy pędów. Ostatnie badania przeprowadzone w Pensylwanii wskazują, że żerowanie mszyc nie przyczynia się do powstawania zarazy pędów. Potrzeba więcej badań, aby określić, czy koniki polne odgrywają rolę w występowaniu zarazy pędów. Większość infekcji wierzchołków pędów występuje pomiędzy okresem, w którym pędy mają około dziewięciu do dziesięciu liści, a zawiązaniem pąków końcowych, kiedy dostępne są źródła inokulum i wektory owadów, a temperatury dzienne wynoszą średnio 16 °C lub więcej.
W latach, w których infekcje kwiatów nie występują,. podstawowe źródła inokulum dla fazy zarazy pędu są następujące zimujące kanarki a w szczególności młode pędy wodne w pobliżu tych kanarków, które ulegają zakażeniu, gdy bakterie przemieszczają się do nich systemowo z brzegów kanarków. Takie ogólnoustrojowe infekcje pędów, nazywane zarazą kanarków, są najwyraźniej inicjowane około 111 DD powyżej 13 °C po green tipie, chociaż widoczne objawy mogą być widoczne dopiero po nagromadzeniu co najmniej 157 DD powyżej 13 °C po green tipie. W przypadku braku infekcji kwiatów, rozwój infekcji zarazy pędów jest często zlokalizowany wokół miejsc, w których zimują kanty.
Chociaż dojrzałe tkanki pędów i kończyn są generalnie odporne na zakażenie przez E. amylovoraUszkodzenia spowodowane przez grad, późne przymrozki o temperaturze -2 °C lub niższej oraz silne wiatry, które uszkadzają liście, mogą doprowadzić do sytuacji, w której dojrzałe tkanki naruszają normalne mechanizmy obronne i dochodzi do infekcji. Znane są przypadki występowania zarazy urazowej nawet na normalnie odpornych odmianach, takich jak 'Delicious'.
Zaraza podkładki, kolejna faza zarazy ogniowej, została ostatnio rozpoznana i jest związana przede wszystkim z bardzo podatnymi podkładkami M.26, M.9 i Mark. Na tych drzewach, już kilka infekcji kwiatów lub pędów na odmianie zrazowej może dostarczyć bakterii, które następnie przeniosą się do podkładki, gdzie często, ale nie zawsze, rozwija się rak i ostatecznie opasuje drzewo. Drzewa dotknięte zarazą podkładki wykazują zazwyczaj objawy spadku i wczesnej śmierci w połowie lub pod koniec sezonu, ale mogą nie być widoczne aż do następnej wiosny.
(c) Dr Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2007 r.
Wzorcowa zaraza gruszy
Symptomy
Model wymaga od użytkownika rozpoznania specyficzny i ciągle zmieniający się imprezy lokalne oraz aspekty ich sadu, które mogą zwiększyć lub zmniejszyć ryzyko wystąpienia zarazy ogniowej w stosunku do innych sadów w regionie. Model wymaga od użytkownika założenia, że ryzyko infekcji zarazą ogniową istnieje zawsze, gdy na drzewach obecne są kwiaty, zwłaszcza w okresie opadanie płatków i okres "po kwitnieniu", gdy na wielu odmianach jabłek i gruszek mogą pozostać rozproszone kwiaty. Użytkownik modelu proszony jest o dokładną ocenę sytuacji na swoim terenie i rozpoczęcie działań kontrolnych, jeśli kwiaty są obecne, poziom ryzyka jest "Wysoki" lub "Ekstremalny", a zwilżenie kwiatów prawdopodobnie wystąpi w ciągu najbliższych 24 godzin.
Struktura modelu
Temperatury i wilgotność: Kluczowym procesem zarazy ogniowej, który musi być modelowany, jest możliwość rozwoju bakterii na słupkach kwiatów jabłoni i gruszy. Wzrost ten jest zależny od temperatury, więc wiarygodne przewidywanie ryzyka infekcji wymaga zastosowania metody pomiarowej, która najdokładniej odzwierciedla wzrost kolonii Erwinia amylovora. Główna różnica zdań między modelami dotyczy tego, jak należy to zrobić.
Wyjście modelu zarazy ogniowej w FieldClimate
Model zarazy Cougar szacuje szybkość wzrostu bakterii z godzinami stopni na podstawie określonej krzywej wzrostu. Ta krzywa wzrostu jest oparta na tempie wzrostu E. amylovora bakterii w testach laboratoryjnych. Wartości godzinowe stopni są gromadzone w każdej godzinie dnia, w której temperatury są wyższe niż 15°C. Wartości godzinowe rosną wraz ze wzrostem temperatury od 15°C do 29°C, maleją przy wyższych temperaturach i osiągają zero dla każdej godziny z temperaturą powyżej 40°C.
Modelowa zaraza kwiatowa dla gruszek
- Kwiat musi być otwarty z nienaruszonymi słupkami i płatkami, słupki muszą być odsłonięte dla kolonizacji, kwiaty w opadnięciu płatków są odporne;
- nagromadzenie co najmniej 110 °C godzin > 18,3°C w ciągu ostatnich 44°C dni > 4,4°C określa potencjał infekcji epifitycznej dla najstarszego otwartego, a więc najbardziej skolonizowanego kwiatu w sadzie
- Zdarzenie zwilżające w postaci rosy lub 0,2 mm deszczu lub 2,5 mm deszczu w dniu poprzedzającym pozwala na przemieszczanie się bakterii ze skolonizowanych słupków do nektarthodes.
- średnia temperatura dobowa >= 15,6°C: Może to wpływać na szybkość migracji bakterii do nektarthodes, jak również na namnażanie się bakterii potrzebnych do ustanowienia infekcji.
Gdy wszystkie cztery z tych minimalnych wymagań są spełnione w przedstawionej kolejności, dochodzi do infekcji i można oczekiwać pojawienia się pierwszych wczesnych objawów zarazy kwiatowej wraz z nagromadzeniem dodatkowych 57 dni °C > 12,7 °C. Może to nastąpić od 5 do 30 dni po infekcji. Gdy warunki w sadzie są mniejsze od tych minimalnych wymagań, objawy występują w niewielkim stopniu lub wcale i nie rozwija się znacząca epidemia. (STEINER P.W. 1996)
Prezentacja graficzna Zaraza ogniowa na gruszkach
W FieldClimate dwa modele zarazy ogniowej są wyświetlane na jednym wykresie. Model Cougar Blight nosi nazwę Fire Blight DIV, a model Maryblight nosi nazwę Blossomblight. Aby zinterpretować wyniki Cougar Blight, wykres jest podświetlony w 5 różnych kolorach. Rozkład tych kolorów jest dokonywany na podstawie ustawień dotyczących pierwszej historii zarazy w sadzie. 5 kolorów wskazuje klasę ryzyka dla wartości DIV.
Możliwość wystąpienia infekcji kwiatów jest wskazywana przez słupek w zakresie 0 - 1 (warunki są spełnione lub nie) na tym samym wykresie. Ustawienia dotyczące historii sadu nie są zintegrowane z tym modelem. Za każdym razem, gdy słupek z infekcją kwiatów jest obliczany w FieldClimate.com jest to infekcja zarazą ogniową!
Praktyczne zastosowanie Zaraza ogniowa na gruszkach
Celem modeli zarazy ogniowej jest ocena prawdopodobieństwa infekcji przez Erwinia amyloflora w sadzie.
Na stronie Model Mary Blight który ocenia się dla zarazy kwiatowej bardzo dobrze wskazuje na sytuacje infekcji o dużym znaczeniu gospodarczym. Z tego powodu jest dość często wykorzystywany do wskazania zastosowania antybiotyków przeciwko temu patogenowi.
Zaraza pucharów daje informacje o ryzyku infekcji zarazą ogniową w związku z ogólnymi możliwościami rozprzestrzeniania się patogenu. Jego waga, wynikająca z historii sadu, jest bardzo pomocna w wskazaniu nam, jak uważnie musimy sprawdzać sady pod kątem objawów zarazy ogniowej, nawet w sytuacjach, w których Mary Blight nie będzie wskazywała na infekcję.
(c) Dr Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2008 r.
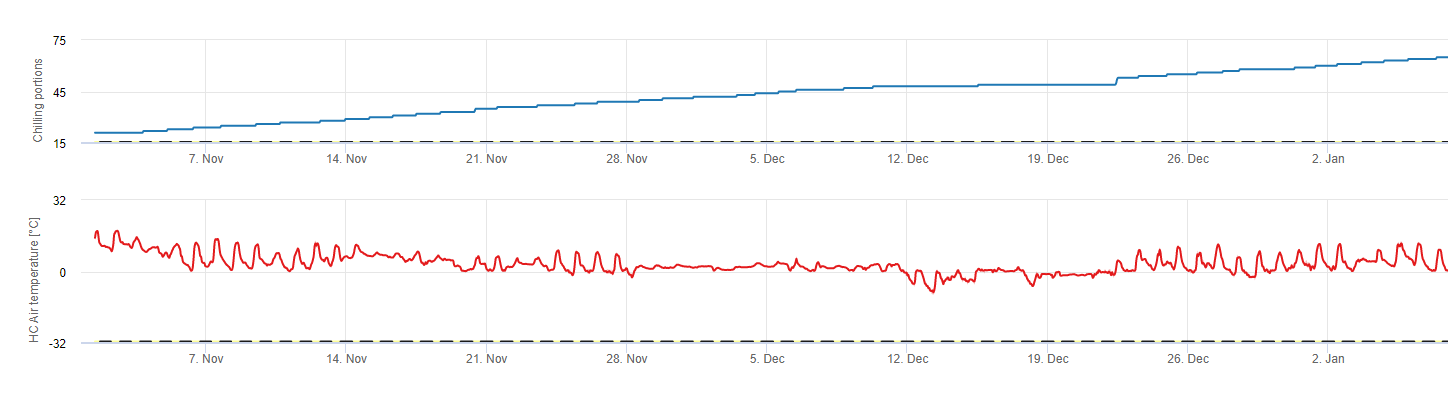
Chłodzenie porcji
Chłodzenie
Drzewa owocowe rozwijają swoje pąki wegetatywne i owocowe latem, a gdy zbliża się zima, rozwinięte już pąki przechodzą w stan uśpienia w odpowiedzi na krótszy dzień i niższe temperatury. Ten stan uśpienia chroni pąki przed nadchodzącą zimną pogodą. Po wejściu w stan spoczynku pąki są odporne na temperatury znacznie poniżej zera i nie rosną w odpowiedzi na ciepłe dni w środku zimy. Pąki pozostają w stanie uśpienia do czasu, gdy zgromadzą wystarczającą ilość jednostek chłodu (CU). Kiedy wystarczające schłodzenie gromadzi się, pąki są gotowe do wzrostu w odpowiedzi na ciepłe temperatury. Tak długo jak jest wystarczająca ilość CUs, pąki kwiatowe i liściowe rozwijają się normalnie. Jeśli pąki nie otrzymują wystarczających temperatur chłodzenia w okresie zimowym do całkowitego zwolnienia spoczynku, u drzew wystąpi jeden lub więcej objawów fizjologicznych związanych z niedostatecznym schłodzeniem: 1) opóźnione zawiązanie owoców, 2) zmniejszone zawiązki owoców i zwiększone guzowatość oraz, 3) obniżona jakość owoców.
Niewystarczające objawy wychłodzenia
Delayed Foliation:
Klasycznym objawem niedostatecznego schłodzenia jest opóźnione listowienie. Drzewo może mieć małą kępkę liści w pobliżu wierzchołków pędów i być pozbawione liści przez 12 do 20 cali poniżej wierzchołków. Dolne pąki w końcu pękają, ale pełne ulistnienie jest znacznie opóźnione, zawiązki owoców są zredukowane, a drzewo osłabione. Ponadto intensywne wysysanie liści z dolnych części drzewa powoduje problemy z zarządzaniem, a normalny rozwój przyszłorocznych pąków owocowych może zostać zaburzony.
Redukcja zawiązków owoców i guzków:
Kwitnienie, w odpowiedzi na niedostateczne schłodzenie, często przebiega według schematu obserwowanego w przypadku rozwoju liści. Kwitnienie jest opóźnione, przedłużone, a z powodu nieprawidłowości w rozwoju słupka i pyłku, zawiązywanie owoców jest ograniczone. U wielu odmian brzoskwini kwiaty opadają przed lub w okolicach rozłupania, ale u innych, takich jak 'Jersey Queen' i 'Harvester', tworzą się guzki. Guziki powstają z kwiatów, które pozornie zawiązały się, ale nie rozwinęły się w pełnowymiarowe owoce. W miarę dojrzewania owoce pozostają małe i zniekształcone. Jeśli przetniesz te owoce, nasiona są martwe. Ponieważ guzki nie są widoczne we wczesnej fazie sezonu, hodowcy nie mogą przerzedzić nieprawidłowych owoców, a rozwijające się guzki służą jako źródło pożywienia i miejsce zimowania dla owadów i chorób.
Obniżona jakość owoców:
Wpływ niedostatecznego schłodzenia na jakość owoców jest prawdopodobnie najmniej omówiony, ale wydaje się być bardzo powszechny, zwłaszcza w centralnym i południowym Teksasie. Wpływ na wzrost liści i zawiązki owoców jest dramatyczny, ale wpływ niedostatecznego chłodzenia na jakość owoców jest subtelny i może wystąpić, gdy inne objawy nie występują. Niedostateczne schłodzenie powoduje, że wiele odmian ma powiększony czubek i zmniejszoną jędrność. Ponadto zabarwienie podłoża owoców może być bardziej zielone niż zwykle, prawdopodobnie z powodu utraty jędrności przez owoce przed pełną zmianą koloru podłoża z zielonego na żółty. Zakres tych problemów jakościowych zależy od odmiany i stopnia niedoboru chłodu.
Modele
Istnieje wiele modeli używanych do obliczania chłodzenia, a każdy z nich definiuje, czym jest jednostka chłodzenia. Trzy najczęściej spotykane modele to model liczby godzin poniżej 45 stopni F (7°C), model liczby godzin pomiędzy 32 a 45 stopni F (2 i 7°C) oraz model Utah. Pierwsze dwa modele są proste i definiują jednostkę chłodzenia jako jedną godzinę poniżej lub pomiędzy określonymi temperaturami. Metoda Utah jest bardziej złożona, ponieważ wprowadza pojęcie względnej skuteczności chłodzenia i ujemnej akumulacji chłodu (lub negacji chłodzenia).
W FieldClimate używamy model obliczania porcji chłodu (CP). Akumulacje chłodu oblicza się jako porcje chłodu, stosując zakres temperatur od 2 do 7°C. Obliczenia proporcji schładzania kończą się po 96 godzinach równej lub większej, a następnie >15°C ' (utrzymuje się w przedziale od7 do 15°C)
Obliczenia oparte są na pracy Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peachs. Acta Hortic 276: 165-174.
Parch gruszy
Źródło: J.W. Travis, J.L. Rytter i K.S. Yoder
Wstęp
Parch gruszy jest ważną gospodarczo chorobą na całym świecie i może powodować poważne straty na podatnych odmianach. Choroba ta stanowi większy problem w krajach europejskich niż w Ameryce Północnej i jest szczególnie istotna w Japonii. Czasami nazywany czarną plamą, parch gruszy przypomina parcha jabłoni (Venturia inaequalis) pod niemal wszystkimi względami, a wywołuje go blisko spokrewniony grzyb, V. pirina. Odmiany gruszy różnią się podatnością na parcha; odmiany odporne w jednym regionie kraju mogą nie być odporne w innym regionie.
Symptomy
Objawy parcha gruszy są bardzo podobne do parcha jabłoni. Zmiany na liściach i ogonkach liściowych zaczynają się jako okrągłe, brązowe plamy, które w końcu stają się aksamitne w wyglądzie. Wewnątrz tych zmian wytwarzane są konidia. W późniejszym okresie sezonu na dolnej powierzchni liści można zaobserwować małe plamki. Są one zwykle wynikiem infekcji późną wiosną lub wczesnym latem. Infekcja liści gruszy nie jest tak powszechna jak parcha jabłoni na liściach jabłoni.
Cykl chorobowy
Zmiany parcha na owocach pojawiają się na końcu kielicha i ewentualnie na bokach owoców. W miarę powiększania się tych zmian stają się one ciemnobrązowe i tworzą duże czarne obszary. Zmiany na niedojrzałych owocach to małe, okrągłe, aksamitne plamy. W miarę dojrzewania owoców pojawiają się ciemniejsze, punktowe plamy. Zainfekowane owoce często mają nieregularny kształt. W przeciwieństwie do parcha jabłoni, w przypadku parcha gruszy częste są infekcje gałązek. Na początku sezonu wegetacyjnego zmiany na młodych pędach pojawiają się jako brązowe, aksamitne plamy. Później zmiany te przekształcają się w skorkowaciałe, przypominające raka obszary. Następnej wiosny w obrębie tych przezimowanych zmian powstają krostki. Krostki te wytwarzają zarodniki (konidia), które utrwalają rozprzestrzenianie się choroby. Grzyb zimuje w liściach na ziemi, a także w postaci grzybni na porażonych gałązkach. Zakażenie liści i owoców gruszy następuje w warunkach podobnych do tych, jakie są wymagane do zakażenia jabłek przez grzyb parcha jabłoni. Głównym źródłem pierwotnego inokulum są askospory. Do zakażenia dochodzi wiosną w fazie zielonego końca pąka kwiatowego. Ascospory znajdujące się w przezimowanych liściach uwalniają się w wyniku opadów deszczu i są przenoszone przez prądy powietrzne na młode liście i owoce. Ascospores kontynuują dojrzewanie w okresie od sześciu do ośmiu tygodni. Źródłem wtórnego inokulum są konidia, które powstają albo w pierwotnych zmianach zainicjowanych przez askospory, albo w krostach na porażonych gałązkach. W ciągu jednego sezonu wegetacyjnego może wystąpić wiele cykli wtórnych. Długość okresu zwilżenia i temperatura wymagana do infekcji zależą od liczby godzin ciągłego zwilżenia i temperatury podczas tego okresu zwilżenia== Wykres Millsa dla określenia okresów infekcji parcha jabłoni wraz z rejestratorem zwilżenia liści lub higrotermografem może dostarczyć informacji dla określenia okresów infekcji parcha gruszy. Zmiany parcha mogą rozwinąć się w ciągu zaledwie ośmiu dni od zakażenia na młodych liściach i w ciągu nawet dwóch miesięcy na liściach starszych. Owoce są również bardziej podatne, gdy są młode; jednakże dojrzałe owoce mogą zostać zainfekowane, jeśli okres zwilżenia jest wystarczająco długi.
Monitoring
W okresie spoczynku hodowcy nie muszą prowadzić monitoringu. Należy skonsultować się z regionalnymi pracownikami Cooperative Extension Service w celu określenia początku dojrzewania askospor. Znajomość sytuacji inokulum parcha w sąsiednich opuszczonych lub komercyjnych sadach może wpłynąć na decyzje dotyczące zwalczania parcha we wczesnym sezonie. W okresie poprzedzającym kwitnienie i trwającym do zawiązania owoców, zarówno w przypadku owoców świeżych, jak i przeznaczonych do przetwórstwa, należy określić okresy infekcji parcha gruszy, obserwując czas trwania zwilżenia liści i średnie temperatury w okresie zwilżenia.
Na stronie Venturia pirina Model infekcji zaprojektowany przez Spottsa, R. A., i Cervantesa, L. A. 1991
Zmienne wejściowe środowiska: Temperatura, czas trwania wilgotności.
Opis modelu: Spotts i Cervantes przedstawiają dane z eksperymentu przeprowadzonego w kontrolowanym środowisku z sadzonkami gruszy, jak również z eksperymentów prowadzonych w polu z wykorzystaniem worków na kończyny, dotyczących wpływu temperatury i czasu trwania wilgoci na infekcje konidialne sadzonek, liści i owoców gruszy. Autorzy nie oceniali warunków infekcji askosporami, ale sugerują, że powinny być one dość podobne do warunków infekcji konidialnej i dlatego ich model może być użyty do przewidywania pierwotnej infekcji przez askospory.
Próg działania: Twórcy modelu zaobserwowali, że minimalny okres wilgotności wymagany do zakażenia liści przez konidia mieścił się pomiędzy wartościami wymaganymi do "lekkiego" i "umiarkowanego" zakażenia jabłek przez V. inequalis. Ascospores according to the Mills table. Dlatego przy stosowaniu tabeli Millsa do infekcji askosporami lub konidiami parcha gruszy autorzy zalecają stosowanie godzin zwilżenia dla "lekkiej" infekcji, aby być bardziej konserwatywnym.
(c) Dr Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2009 r.
Fabraea leaf spot
Plamistość liści, powodowana przez grzyba Fabrea maculatum, jest szeroko rozpowszechnioną i niszczącą chorobą czerwonego czubka (Photinia fraseri), loquat (Eriobotrya japonica), głóg indyjski (Rhaphiolepis indica), niektóre odmiany gruszy (Pyrus sp.) i kilku innych członków rodziny różowatych. Choroba ta jest najbardziej szkodliwa dla roślin w krajobrazie i szkółkach w okresach chłodnej, wilgotnej pogody oraz w czasie aktywnego wzrostu.
Symptomy
Drobne, okrągłe, jasnoczerwone plamki zarówno na górnej, jak i dolnej powierzchni młodych, rozwijających się liści to pierwsze objawy Entomosporium plamistość liści. Na silnie chorych liściach liczne małe plamki mogą łączyć się w duże bordowe plamy. Plamy na dojrzałych liściach mają popielato-brązowe lub jasnoszare środki z charakterystyczną, głęboko czerwoną lub bordową obwódką. W centrum każdej plamki można często zauważyć małe czarne plamki, czyli ciała przetrwalnikowe grzyba. Plamy podobne do tych na liściach mogą pojawić się na ogonkach liściowych i delikatnych przyrostach łodyg podczas dłuższych okresów chłodnej, wilgotnej pogody.
Niski poziom plamistości liści powoduje zazwyczaj tylko kosmetyczne uszkodzenia, ale utrzymuje źródło zarodników dla przyszłych infekcji. Silne infekcje powodują jednak często wczesne i obfite opadanie liści. Silne opadanie liści poważnie obniża wartość krajobrazową czerwonej końcówki i może spowodować śmierć rośliny. Niektóre odmiany głogu indyjskiego są równie mocno porażane jak głóg czerwony.
Cykl chorobowy
Plamy na liściach i młodych pędach są ważne dla przetrwania Entomosporium grzyb plamistości liści. Opadłe, chore liście są mniej ważnym źródłem grzyba. Masy zarodników są uwalniane podczas okresów wilgotnej pogody z grzybowych struktur produkujących zarodniki w centrum plam od późnej zimy przez większą część roku, z wyjątkiem gorących okresów lata. Zarodniki te są rozprzestrzeniane na zdrowe liście przez połączenie rozpryskiwania wody i wiatru. Objawy nowej plamistości liści pojawiają się w ciągu 10-14 dni po wilgotnym okresie infekcji.Model infekcji: Czterokomórkowe konidia, o charakterystycznym owadzim wyglądzie, są rozprzestrzeniane głównie z zimującej ściółki liściowej, a częściowo z sęków gałązek, przez rozpryskiwanie wody z deszczu lub nawadniania górnego. Okres zwilżenia dla infekcji może wynosić od 8 do 12 godzin w temperaturze od 10°C do 25°C. Zmiany zaczynają pojawiać się po około 7 dniach od rozpoczęcia okresu infekcji. Choroba może się szybko rozwijać późnym latem, gdy wiatr i deszcz rozprowadzają konidia po całym drzewie. Podatność liści i owoców na infekcję nie zmniejsza się wraz z dojrzałością. Prawie wszystkie grusze pochodzenia europejskiego są podatne na tę plamistość liści. Model ten jest uruchamiany, gdy wilgotność liści zaczyna się od deszczu. Zatrzymuje się, gdy wilgotność liści jest zakłócona na dłużej niż 1 godzinę.
(c) Dr Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2008 r.
Brązowa plama
Brązową plamistość na gruszy wywołuje grzyb patogeniczny Stemphylium vesicarium, który powoduje również choroby na czosnku, porze, cebuli i szparagach. Na gruszy grzyb ten infekuje liście, owoce i w mniejszym stopniu gałązki. Powstające nekrozy i zgnilizny owoców są spowodowane penetracją przez grzyba komórek szparkowych i przetchlinek oraz wytwarzaniem związków chemicznych, które zmuszają gospodarza do zabijania komórek w zainfekowanym miejscu, powodując brązowe plamy. Brunatna plamistość powoduje poważne szkody zwłaszcza w południowej Europie. Jednak choroba ta została stwierdzona również w Dodatkowo, dominująca w Holandii i Belgii odmiana gruszy, jest bardzo podatna (Montesinos i in., 1995a).
Wpływ temperatury i czasu trwania wilgotności na zakażenie konidialne przez S. vesicarium na gruszy była już wcześniej badana (Montesinos i in., 1995b). Wyniki tych badań doprowadziły do opracowania systemu prognozowania brunatnej plamistości (Llorente i in., 2000). Warunki klimatyczne w Europie są zróżnicowane z południa na północ. W Europie Południowej możemy spodziewać się burz nawet w ciepłych okresach, natomiast w Europie Północnej kilkugodzinne chłodne opady mogą występować nawet w połowie lata. Jest oczywiste, że system prognozowania powinien być rewalidowany lub nawet przystosowany do stosowania w innych warunkach klimatycznych.
Okresy wilgotne prowadzące do Stemphylium versicarium infekcje na gruszce muszą być bardzo długie zgodnie z tym modelem. Model ten pochodzi z prac prowadzonych przez Llorente, I., Vilardell, P., Moragrega, C. i Montesinos, E. oraz z adaptacji do elektronicznych stacji meteorologicznych dokonanej przez A. Boshuizen, P.F. de Jong i B. Heijne z Holandii. Te wilgotne okresy mogą być zakłócone. Długość przerwy zależy od wilgotności względnej lub deficytu ciśnienia pary wodnej.
W obliczeniach modelu FieldClimate zaburzenie może trwać wiecznie, o ile wilgotność względna jest większa lub równa 75%. Przy wilgotności względnej pomiędzy 65% a 74% zaburzenie okresu wilgotnego może trwać 12 godzin. Przy wilgotności względnej pomiędzy 55% a 64% zaburzenie może trwać 9 godzin. Jeżeli wilgotność względna mieści się w przedziale od 45% do 54%, to zaburzenie może trwać 8 godzin. Przy wilgotności względnej pomiędzy 35% a 44% zakłócenie może trwać 6 godzin. Jeśli wilgotność względna jest poniżej 34%, zakłócenie może trwać 4 godziny.
Krzywe obrazujące przebieg infekcji lekkich, umiarkowanych i ciężkich mogą być wykorzystane jako progi działania dla sadów o różnej historii choroby. W sadach z ponad 1% występowania choroby na owocach należy przeprowadzić chemiczne zwalczanie choroby w przypadku wszystkich lekkich infekcji. W sadach, w których wystąpiło mniej niż 1% choroby na owocach chemiczne zwalczanie choroby powinno być przeprowadzone w przypadku wszystkich zakończonych umiarkowanych infekcji. Natomiast w sadach, w których do tej pory nie występowały Stemphylium metody kontroli należy rozpocząć, jeśli ciężkie infekcje osiągną 100%.
Warunki:
Temperatura: 8°C - 38°C
Wilgotność liści > 0 lub wilgotność względna >90%
Czynnik: 600; max: 60000 (infekcja 100%).
(c) Dr Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2009 r.
Zalecane wyposażenie
Sprawdź, jaki zestaw czujników jest potrzebny do monitorowania potencjalnych chorób tej uprawy.