Biologia Plasmopara viticola (Downy Mildew)
P. viticola jest obowiązkowym pasożytem. Co oznacza, że do rozwoju potrzebne są zielone, świeże organy winorośli. W okresie wolnym od wegetacji utrzymuje się tworząc owocniki, tzw. oospory. Oospory oomycetes mogą przetrwać bardzo długi okres w glebie. Dlatego też możemy spotkać mączniaka w miejscach, gdzie infekcje nie są możliwe w każdym roku. Wiosną, gdy wierzchnia warstwa gleby jest wystarczająco wilgotna i ciepła, oospory utworzą tzw. makrosporangia, które mogą uwolnić do wolnej wody do 200 zoospor. Zoospory przenoszą się na liście i skupiska przez wiatr w kropelkach wody. Posiadają one dwie flagi i poruszają się w warstwie wody na spodniej stronie liści lub na klastrach i młodych jagodach, aby znaleźć stomię, przez którą dostaną się do tkanki roślinnej. Wchodzą i kiełkują do stomii, w której przenoszą całą swoją plazmę w ciągu mniej niż jednej godziny. W badaniach mikroskopowych znalezienie stomii, encystacja i kiełkowanie do stomii kończyło się w ciągu 90 minut.
P. viticola rośnie w przestrzeni międzykomórkowej i odżywia się haustoriami wnikającymi w komórki epidermy i miąższu. W zależności od temperatury i wilgotności względnej rozwija się wystarczająco dużo wzrostu międzykomórkowego z wystarczającą ilością haustorii, aby utworzyć ciało podkomórkowe, które wypełnia cały obszar podkomórkowy i które podnosi tkankę naskórkową z tkanki miąższowej. Prowadzi to do widocznego objawu w postaci plamy olejowej.
Oomycetes to. sporulujące przy braku światła, gdy wilgotność względna jest bardzo wysoka. Na stronie P. viticola nie dochodzi do sporulacji, jeśli temperatura jest poniżej 12°C, a wilgotność względna poniżej 95%. Sporangiaphores są tworzone przez pęcherzyki substomatelowe i będą one wychodziły ze stomii. Świeżo uformowane sporangia są lepkie i można je usunąć ze sporangiów tylko za pomocą wody. Podczas spadku wilgotności względnej powietrza sporangia stają się lepkie i mogą być usunięte przez wiatr.
Sporangia uwalniają do 20 zarodników do wolnej wody. Te zarodniki muszą być rozprowadzone przez wiatr w kropelkach wody, aby dotarły do świeżych liści, lub sporangia mogą być rozprowadzone przez deszcz lub sam wiatr. Proces infekcji pierwotnej i wtórnej jest taki sam.
Ze względu na duże znaczenie fazy płciowej dla hibernacji patogenu możemy przyjąć typy kojarzenia pasujące do wszystkich winnic, w których występuje mączniak rzekomy winorośli. Tworzenie zoospor ma miejsce na starszych liściach w okresie letnim i wczesną jesienią.
Zakażenie pierwotne w szczegółach
Infekcje pochodzące z oospor nazywane są infekcjami pierwotnymi. Termin ten jest mylący, ponieważ na początku lata może wystąpić kilka infekcji pierwotnych. W zdarzeniach epidemiologicznych infekcja pierwotna nie odgrywa istotnej roli, jeśli w winnicach jest wystarczająca ilość plam olejowych, a potencjał infekcyjny zarodników letnich (sporangiów) przewyższa oospory.
Zimujące oospory rozwijają się do tzw. makrosporangiów, gdy tylko przez około 24 godziny jest wystarczająca wilgotność względna. Makrosporangia uwalniają swoje zarodniki do wolnej wody i silnego deszczu (np. podczas burzy). Zarodniki przenoszone są na liście i/lub zielone pędy winorośli. Infekcje pierwotne, a więc, potrzebują dłuższego deszczu okresy lub kilka następujących po sobie burz. Pierwszy deszcz powoduje nasycenie opadłych liści wodą, a silne opady w kolejnych dniach powodują uwalnianie przez makrosporangia swoich zoospor, które docierają do liści lub pędów winorośli. Do skutecznej infekcji niezbędny jest odpowiednio długi okres zwilżenia liści, aby zoospory mogły dotrzeć do stomii liści lub pędów i je zainfekować (Ilustracja powyżej przedstawia cykl rozwojowy mączniaka rzekomego winorośli (Plasmopara viticola).
Model infekcji pierwotnej mączniaka sprawdza najpierw, czy pogoda jest odpowiednia do rozwoju makrosporangiów. Tak jest, jeśli liście są wilgotne lub wilgotność względna po deszczu nie spada poniżej 70%. W zależności od temperatury, makrosporangia mogą się rozwinąć w ciągu 16-24 godzin. Jeśli dojrzałe makrosporangia są dostępne, zostanie to pokazane graficznie na wyświetlaczu infekcji pierwotnej Downy Mildew. Jeśli makrosporangia są obecne, silny opad deszczu może spowodować rozprzestrzenienie się ich zarodników. Ciągły deszcz o wielkości 5 mm jest interpretowany jako silny deszcz i zarodniki są rozprzestrzeniane, a infekcja pierwotna jest możliwa, jeśli zwilżenie liści trwa wystarczająco długo.
 Zakażenia wtórne
Zakażenia wtórne
 Zakażenia wtórne
Zakażenia wtórneWtórne zakażenia Plasmopara viticola są możliwe tylko wtedy, gdy w Twojej winnicy lub w jej pobliżu istnieją już dojrzałe plamy olejowe. Plamy olejowe są dojrzałe, gdy są w stanie sporulować (wytwarzać sporangia). Sporangia są wytwarzane tylko przez noc. Światło słoneczne hamuje sporulację. Sporangia powstają, jeśli jest cieplejsze niż 12°C i wilgotność względna jest bardzo wysoka. Tempo wytwarzania sporangiów wzrasta wraz z temperaturą do 24°C. Optymalna temperatura dla sporulacji na europejskich odmianach winogron (Vitis vinifera) wynosi około 24°C. Jeśli temperatura przekracza 29°C, nie może dojść do sporulacji. W naszym modelu sprawdzamy, czy wilgotność powyżej 95% występuje w nocy. Jeśli ten stan utrzymuje się przez skumulowaną temperaturę godzinową powyżej 50°C, sporulacja zostaje zakończona i pojawiają się nowe sporangia Plasmopara istnieją w winnicy. Na przykład godziny z 50°C odpowiadają 4 godzinom z 13°C lub 3 godzinom z 17°C.
Sporangia z Plasmopara viticola mają ograniczoną żywotność. Im jest cieplej i im bardziej suche powietrze, tym szybciej giną (w fieldclimate.com resetujemy do 0, gdy r.h. jest poniżej 50%). Na pewno giną w następnym okresie rosy lub zwilżenia liści, który jest zbyt krótki dla infekcji. Fitness starszych sporangiów jest więc dość ograniczony. Nasz model zakłada, że czas życia sporangiów jest ograniczony do jednego dnia.
Aby wywołać nowe infekcje, sporangia muszą zostać rozprowadzone. Istnieją dwa sposoby rozprowadzania zarodników: Jeśli bezpośrednio po uformowaniu się sporangiów pada deszcz, rozprzestrzeniają się one wraz z rozpryskami deszczu. Jeśli liście winorośli pozostaną wystarczająco długo mokre, wysoki poziom Plasmopara viticola dochodzi do infekcji. Jeśli kolejny poranek zaczyna się bez deszczu i ze zmniejszającą się wilgotnością, zaschnięte sporangia odrywają się od rozgałęzień. Nawet niewielki ruch powietrza przeniesie je na zdrowe liście. Jeśli wkrótce nie spadnie deszcz, zginą.
Mączniak prawdziwy Infekcja pierwotna
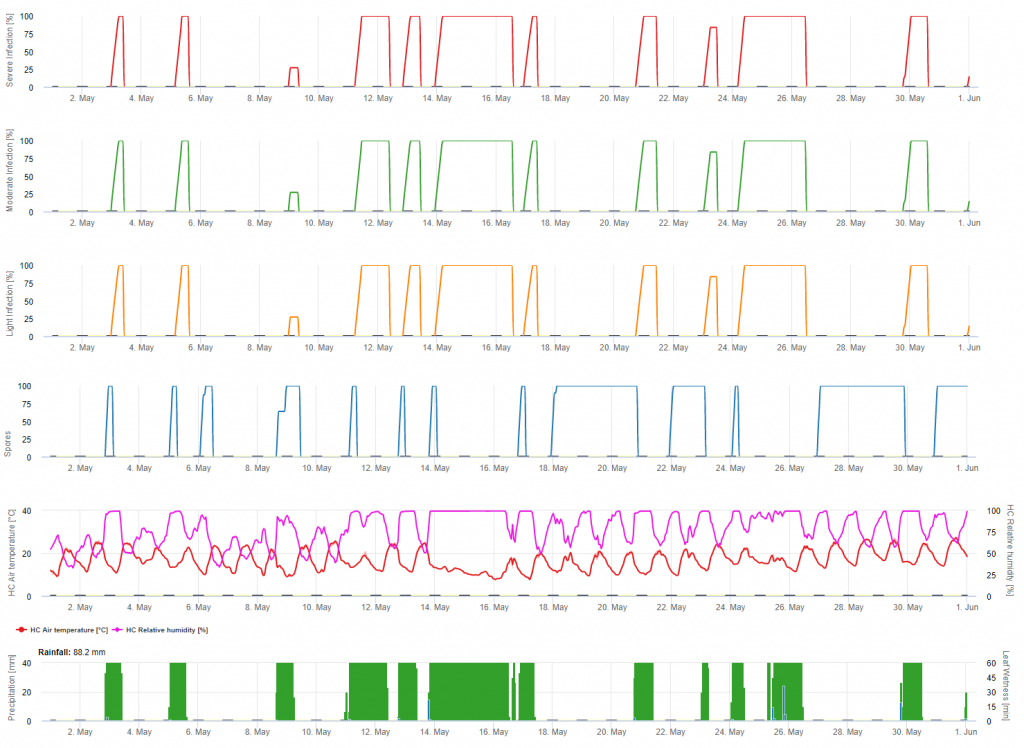
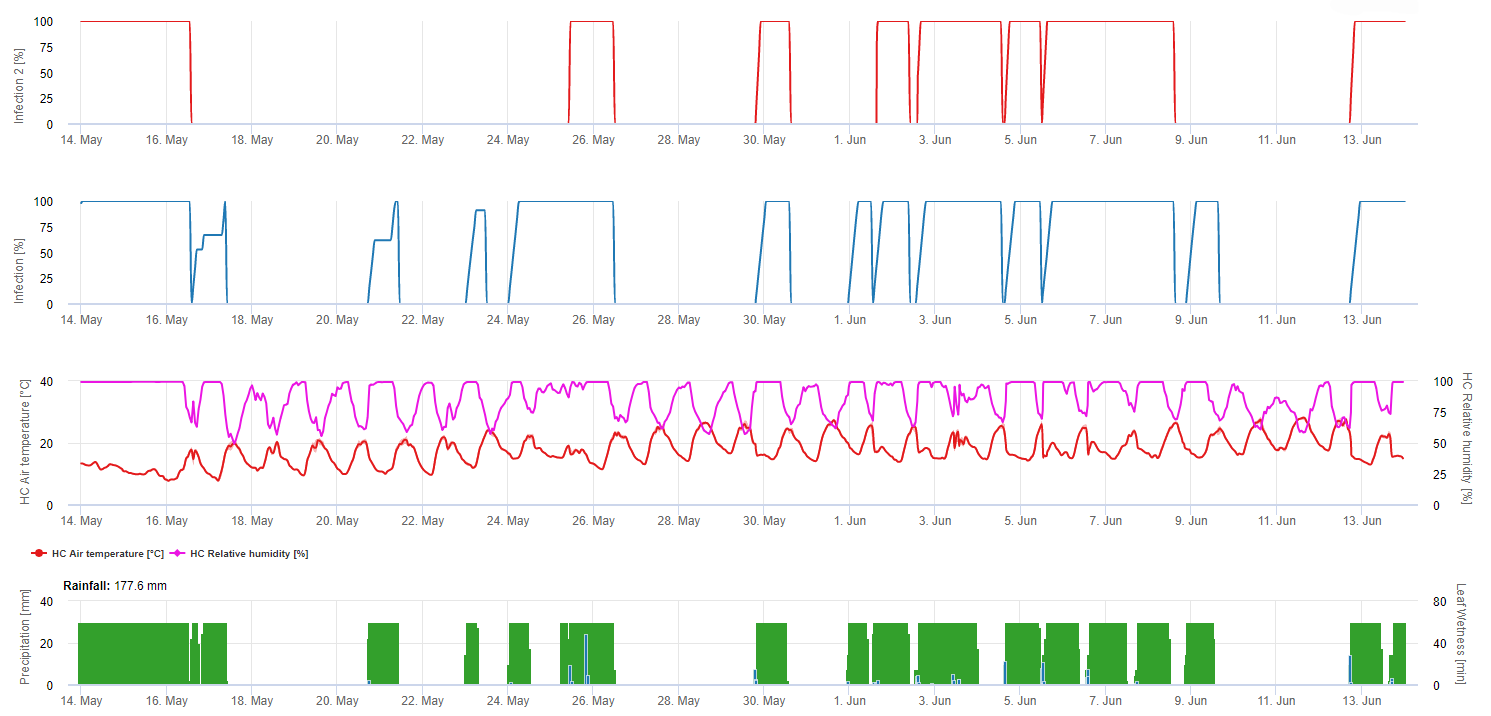
Infekcje są przedstawione po rozwinięciu makrospor (kolor pomarańczowy, piąty wykres) w trzech różnych klasach ciężkości (słaba: pomarańczowa, umiarkowana: zielona, ciężka: czerwona) w zależności od ilości deszczu, a tym samym dystrybucji zarodników w winnicy. W przypadku stwierdzenia infekcji 100% zapewniono optymalne warunki do wniknięcia w tkankę roślinną (środek ochrony roślin) i krzywa inkubacji (pierwszy wykres) zaczyna się zwiększać. W przypadku zakończenia inkubacji (100%) i braku ochrony roślin należało sprawdzić na polu objawy (plamy olejowe).
1. Krzywa postępu infekcji, a po pełnej infekcji wynikający z niej okres inkubacji (czas pomiędzy infekcją a widocznymi objawami w terenie).
2. Krzywa postępu infekcji jest wyznaczana, gdy obecne są makrosporangia lub sporangia i liście są wilgotne. Wzrasta ona wraz z postępującą wilgotnością liści i temperaturą. Jeśli krzywa postępu infekcji osiągnie 100%, można uznać, że infekcja została zakończona. Krzywa postępu inkubacji, która należy do konkretnej infekcji, zaczyna być obliczana wraz z rozpoczęciem infekcji. Jeśli infekcja nie może być zakończona, krzywa postępu inkubacji również zostanie zatrzymana.
3. Warunki pogodowe są wyświetlane na tej samej stronie co wyniki modelu choroby. Dlatego jednym spojrzeniem można sprawdzić temperaturę, wilgotność względną, deszcz i zwilżenie liści podczas infekcji.
4. Model wskazuje terminy infekcji dla pierwotnych i wtórnych infekcji mączniaka prawdziwego winorośli. Jest to najbardziej krytyczna informacja. Infekcje, które nie zostały jeszcze zwalczone przez fungicydy zapobiegawcze lub lecznicze, mogą być katastrofalne dla plonu/jakości. Dalsze informacje dotyczące nasilenia infekcji (słabe/umiarkowane/ciężkie) mogą być pomocne w podjęciu decyzji o strategii ochrony roślin w zależności od historii winnicy (ilość inokulum), odmiany i doświadczeń z poprzednich lat.
Strategie ochrony roślin:
- Do zwalczania mączniaka prawdziwego powszechnie stosuje się fungicydy profilaktyczne i systemiczne lub lecznicze. Profilaktyczne środki chemiczne stosuje się przed, ale jak najbliżej zdarzenia infekcyjnego w okresie największej podatności gospodarza, pomiędzy długością pędów 10 cm a wielkością jagód grochu.
- Profilaktyczne środki chemiczne można stosować również poprzez opryskiwanie jak najbliżej przed prognozowanym zdarzeniem pogodowym o deszczu większym niż 2 mm (dla kręgu podstawowego).
- Fungicyd leczniczy powinien być zastosowany jak najszybciej po wystąpieniu infekcji (infekcja 100%) i przed pojawieniem się plam olejowych. Należy pamiętać o zapobieganiu powstawania odporności na fungicydy.
- Monitorowanie winnicy pod kątem obecności mączniaka prawdziwego (plamki olejowe) powinno odbywać się regularnie, z uwzględnieniem co najmniej 200 winorośli. Za zagrożenie uznaje się, jeśli zaobserwuje się więcej niż dwie plamy olejowe na 50 winorośli.
Model opiera się na publikacjach MUELLER-THURGAU, ARENS, MUELLER i SLEUMER, BLAESER, HERZOG, GEHMANN i wielu innych pracowników związanych z siecią badawczą epidemiologii mączniaka rzekomego winorośli od początku XIX wieku.
Biologia Uncinula necator
Mączniak rzekomy winorośli jest powodowany przez grzyby Uncinula necator. Jest pasożytem obligatoryjnym, co oznacza, że potrzebuje zielonej tkanki gospodarza, aby się odżywiać. U. necator W Europie pojawił się w XIX wieku i do końca tego stulecia rozprzestrzenił się we wszystkich krajach uprawiających wino. Ma bardzo silny wpływ na plony i może prowadzić do defoliacji winorośli, jeśli nie ma kontroli chemicznej. Wszystkie europejskie odmiany winorośli są podatne na ten patogen. Hodowla odporności na ten patogen jest dość nowa, a jej wyniki wymagają kolejnych lat pracy.
U. necator musi pokonać okres wolny od wegetacji na większości obszarów uprawy winorośli. Aby to zrobić patogen wykorzystuje dwa mechanizmy: a) przetrwanie w owocnikach, tzw. cleistothecias lub b) przetrwanie zimy zbyt zamknięte w uśpionych pąkach, gdzie znajduje żywą tkankę przez całą zimę. Obecność tylko jednego lub obu sposobów hibernacji w jednej winnicy jest oparta na historii choroby tej winnicy. Pąki mogą być zakażone tylko tak długo, jak długo nie są zamknięte i otoczone pierścieniem tkanki starczej. Tak jest tylko przez kilka tygodni po uformowaniu. W normalnych warunkach uprawy winorośli przycina się je do gałęzi z mniej niż 14 pąkami lub do ostróg. Te pozostałe pąki mogą być zainfekowane tylko do momentu zakwitnięcia. Z drugiej strony możemy znaleźć pędy flagowe od stadium BBCH 15 do 19. Pędy flagowe to częściowo lub całkowicie pokryte mączniakiem pędy powstałe z utajonych zainfekowanych pąków. Pędy te są wczesnym i bardzo silnym źródłem inokulum. W większości przypadków nie są one rozmieszczone w winnicy. Prowadzi to do powstawania miejsc o wczesnym i wysokim występowaniu choroby wewnątrz winnicy oraz miejsc o niskim lub umiarkowanym występowaniu choroby na początku sezonu. Pędy flagowe reprezentują tylko jeden typ kojarzenia ze względu na ich historię wegetacyjną. Może to prowadzić do winnic, w których znajdziemy tylko kilka lub żadnych cleistothecias z powodu dominacji jednego typu kojarzenia.
Tam, gdzie cleistothecia jest jedynym inokulum, znajdziemy ciągłą historię hibernacji cleistothecia, która może zmienić się w pędy flagowe tylko w latach, w których choroba rozwija się bardzo wcześnie. Tworzenie się cleistothecia jest dobrze widoczne ze względu na żółte zabarwienie tkanki płciowej połączonej. W nieopryskiwanych winnicach można ją znaleźć od połowy lata. W winnicach opryskiwanych potrzebuje więcej czasu, aby zbudować występowanie choroby, umożliwiając spotkanie typów kojarzeniowych. Rozprzestrzenianie się Uncinula necator W okresie letnim wpływ ma przede wszystkim temperatura.
Optymalne temperatury dla mączniaka rzekomego wynoszą od 21°C do 32°C. Im większa liczba godzin dziennie w tym zakresie temperatur, tym większe ryzyko wystąpienia mączniaka. Tworzenie konidiów, infekcja i szybkość inkubacji wymagają podobnych zakresów temperatur. Ze względu na fakt, że U. necator nie potrzebuje wolnej wody do infekcji i wysokiej wilgotności względnej do tworzenia konidiów, nie ma użytecznego wpływu parametru wilgotności bezpośrednio na patogen.
Obecność wolnej wilgoci oddziałuje na U. necator poprzez wspólnego hiperpasożyta Ampelomyces quisqualis. Ten hiperpasożyt grzybowy rozwija się wewnątrz hyfy grzyba mączniaka prawdziwego. Przekształca konidia i cleistothecia we własne pycnidia. Potrzebuje wolnej wilgoci lub bardzo wysokiej wilgotności względnej do infekcji i wzrostu z cienkimi, szybko rosnącymi strzępkami na liściach, aby znaleźć zmiany mączniaka. Silnie zainfekowany U. necator Zmiany chorobowe zmieniają kolor na bardzo ciemny brąz. W okresie deszczu pokryte są białym wysiękiem zarodników pochodzących z piknidiów hiperpasożyta. Ampelomyces quisqualis jest dostępny również jako produkt handlowy do zwalczania mączniaka.
Praca praktyczna
Wprowadziliśmy różne modele dotyczące procesu infekcji do FieldClimate.
a) grzyb zimuje głównie w postaci grzybni. Wiosną grzybnia zaczyna rosnąć i buduje konidiofory z konidiami (nie ma więc infekcji pierwotnej).
b) jeśli grzyb rozwija się jesienią na jagodach, może być obecna forma płciowa (budująca cleistothecia), a więc wiosną uwalnianie askospor może prowadzić do infekcji pierwotnej (wykorzystanie: model POWDERY MILDEW/ASCOSPORE INFECTION).
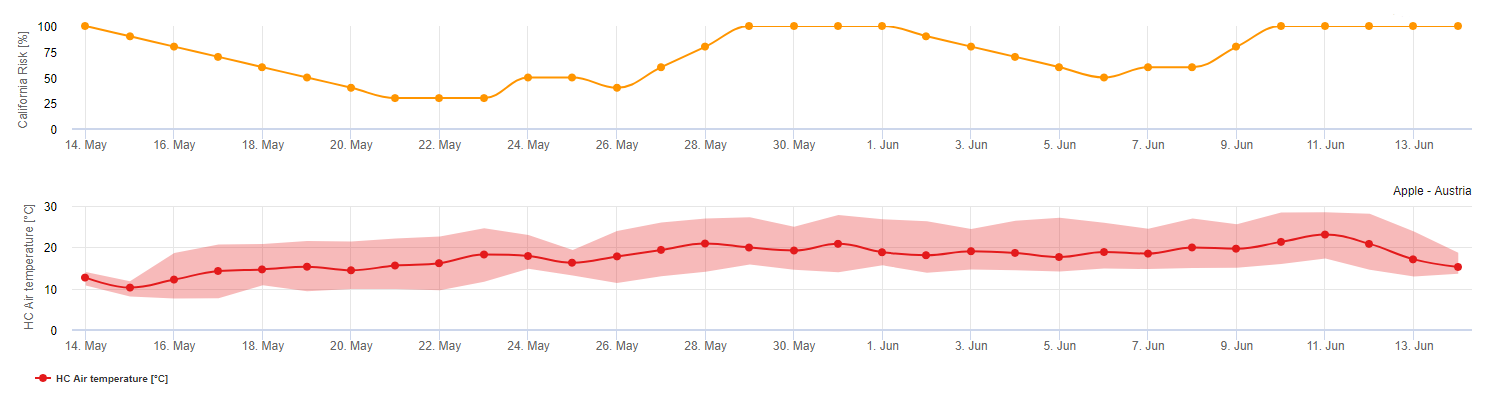
Kalifornijski model ryzyka (autorstwa Thomasa Gublera)
Model oceny ryzyka mączniaka rzekomego UC Davis (model Gubler-Thomas) jest oparty na laboratoryjnych i terenowych badaniach epidemiologicznych mączniaka rzekomego winorośli w Kalifornii.
Indeks oceny ryzyka:
Po uwolnieniu i wykiełkowaniu askospor (w tym modelu należy użyć ASCOSPORE INFECTION), na dalszy rozwój i rozmnażanie mączniaka wpływa przede wszystkim temperatura otoczenia. Model UC Davis ocenia temperatury otoczenia i szacuje ryzyko rozwoju mączniaka przy użyciu indeksu 0-100 punktów.
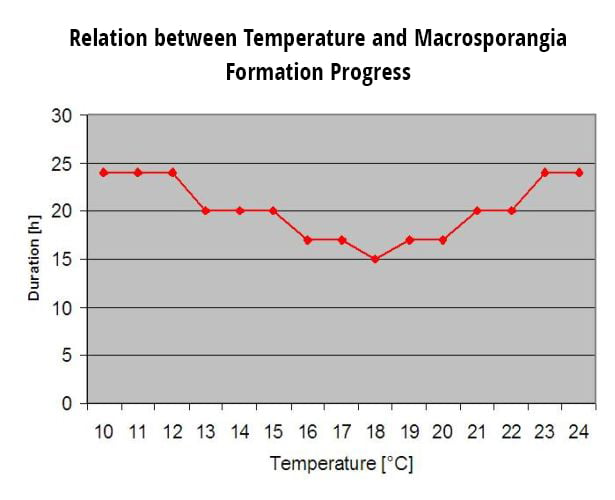
Aby zainicjować indeks oceny ryzyka, po pęknięciu pąka i po wystąpieniu choroby muszą wystąpić trzy kolejne dni z co najmniej sześcioma kolejnymi godzinami z temperaturami pomiędzy 21 a 30 °C, aby można było rozpocząć epidemię mączniaka. Podczas obliczania wskaźnika, za każdy dzień z 6 lub więcej kolejnymi godzinami z temperaturą pomiędzy 21 a 32°C, wskaźnik wzrasta o 20 punktów. Następnie model waha się między 0 a 100 punktami indeksu w oparciu o czas trwania temperatur dziennych. Indeks zyskuje 20 punktów za każdy dzień, który spełnia wymóg sześciu kolejnych godzin temperatur pomiędzy 21 a 32°C, a traci 10 punktów za każdy dzień, który nie spełnia wymogu sześciu godzin, lub jeśli w dowolnym momencie w ciągu dnia temperatura przekroczy 32°C lub będzie poniżej 21°C.
Interpretacja wskaźnika:
Niskie wartości indeksu 0-30 wskazują, że patogen nie rozmnaża się. Hodowcy mogą rozważyć odłożenie aplikacji fungicydów podczas dłuższych okresów z niskimi wartościami indeksu. Indeks 40-50 jest uważany za umiarkowany i oznacza, że tempo rozmnażania mączniaka wynosi około 15 dni. Wartości indeksu 60-100 wskazują, że patogen rozmnaża się szybko (co 5 dni) i że ryzyko epidemii choroby jest duże.
Model ryzyka Pessl Instruments
Rozproszenie Uncinula necator W okresie letnim wpływ na to mają przede wszystkim temperatura. Antagonista U. necator o nazwie Ampelomyces quisqualis sprzyja głównie wolna wilgoć. Optymalne temperatury dla mączniaka wynoszą od 21°C do 32°C. Im większa liczba godzin dziennie, w których temperatura mieści się w tym zakresie, tym większe jest ryzyko wystąpienia mączniaka (patrz model Gublera). Dni z temperaturą powyżej 6 godzin w tym zakresie zwiększają ryzyko o około 20 punktów. Dni, w których temperatura nie mieści się w tym optymalnym zakresie, zmniejszają ryzyko o około 10 punktów. Są to dni z temperaturą poniżej 21°C przez cały dzień lub dni, w których temperatura przekracza 32°C przez 6 godzin. Natomiast dni z mokrość długich liści okresy prowadzą do zasiedlenia antagonistycznego grzyba A. quisqualis. Doprowadzą one do zmniejszenia U. necator.
W przypadku tego modelu zalecamy przestrzeganie tych zasad: Jeśli ryzyko wystąpienia mączniaka jest mniejsze niż 20 punktów, można wydłużyć okres opryskiwania. Przy 20-60 punktach obowiązuje normalny odstęp między opryskami. Jeśli ryzyko jest większe niż 60 punktów, należy skrócić okres opryskiwania.
Zaczyna się ryzyko: z 3 dni z równą lub większą niż 6 godzin: 21°C <= Temperatura 60 pkt.
Ryzyko wzrasta: Przy każdym dniu o równej lub większej niż 6 godzin: 21°C <= Temperatura +20 pkt.
Ryzyko maleje: Z każdy dzień, w którym temperatura nie osiąga 21°C lub każdy dzień, w którym 6 godzin: 32°C -10 pkt.
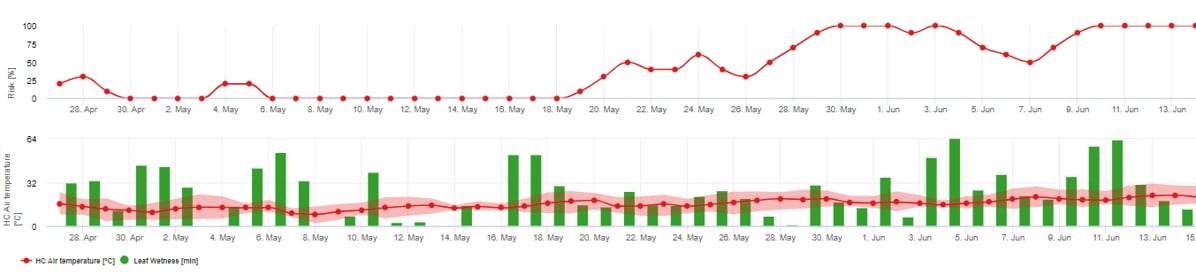
Trzeci model ryzyka: Pierwotne inokulum askospor.
Zalecamy stosowanie tego modelu, gdy askospory powstają w czasie mroźnej zimy (a więc nie są zakażane przez grzybnię ("pędy flagowe").
Przewiduje się uwolnienie askospor i początkowe infekcje, które są oparte na średnich temperaturach podczas przedłużonych okresów wilgotności liści.
Do obliczenia w fieldclimate.com około 2,5 mm opadów są wymagane do uwolnienia askospor, a następnie przy co najmniej 8-12 godzin zwilżenia liści po uwolnieniu askospory w temperaturze pomiędzy 10 -15°C do zakażenia. Po wystąpieniu infekcji model przechodzi w fazę oceny ryzyka choroby (zastosowanie modelu Gubler/Risk) i opiera się całkowicie na wpływie temperatury na tempo reprodukcji patogenu.
Zastosowanie praktyczne
W tym modelu wyświetlamy infekcję przez Uncinula necator w wierszu od 0 do 100%.
Wyświetlamy dwa różne modele w FieldClimate:
1. Możliwe infekcje askosporami według modeli zaproponowanych przez Davida Gadoury'ego i Douga Gublera są wyświetlane na wykresie dla infekcji askosporami. Jeśli krzywa postępu dla modelu infekcji osiągnie 100%, istnieje możliwość infekcji askosporami. 3 do 5 dni po tym zdarzeniu pierwsze objawy mączniaka mogą być widoczne na liściach w pobliżu starego kordonu.
2. Zagrożenie mączniakiem według modelu kalifornijskiego i modelu dla umiarkowanych stref klimatycznych przedstawiono na powyższym wykresie. Okresy z ryzykiem muszą być objęte aplikacją fungicydów. W okresach z ryzykiem wyższym niż 60 punktów zaleca się częstsze opryski, szczególnie w okolicach i krótko po kwitnieniu.
Czarna zgnilizna winogron dotyka wielu plantatorów winogron w całych Stanach Zjednoczonych oraz w większości innych ciepłych i suchych obszarów uprawy winogron na całym świecie. Aby poradzić sobie z tą chorobą, ważne jest zrozumienie jej cyklu życiowego i warunków środowiskowych. Gdy dojdzie do infekcji, dostępne są różne metody kontroli choroby.
Biologia
Patogen czarnej zgnilizny winogron zimuje w wielu częściach winorośli, a także jest w stanie przezimować na ziemi, zwłaszcza w mumiach. Ponadto patogen może przezimować przez "co najmniej 2 lata w obrębie zmian chorobowych zainfekowanych pędów, które zachowały się jako pędy lub ostrogi."
Deszcze mogą kiedyś uwolnić zimujące zarodniki (askospory), które tworzą się w mumiach na ziemi i mogą być rozproszone przez wiatr. Niektóre z mumii na ziemi mogą mieć znaczny wysyp askospor, który rozpoczyna się około 2 do 3 tygodni po pęknięciu pąków i dojrzewa 1-2 tygodnie po rozpoczęciu kwitnienia. Drugi rodzaj zarodników (Conidia) może również tworzyć się w obrębie uszkodzeń trzciny lub mumii, które pozostały w obrębie "trelinki, a te są rozpraszane na niewielkie odległości (cale do stóp) przez rozpryskujące się krople deszczu". Infekcja następuje, gdy któryś z typów zarodników wyląduje na zielonej tkance winogron i tkanka pozostanie wilgotna przez "wystarczająco długi czas, który zależy od temperatury." Okres, w którym te zimujące zarodniki mogą spowodować infekcję, zależy od źródła. Jeśli jest duże źródło infekcji, infekcja pojawi się wcześnie. W obecności wilgoci, te askospory powoli kiełkują, zajmując 36 do 48 godzin, ale w końcu wnikają w młode liście i szypułki owoców (pedicels). Infekcje stają się widoczne po 8 do 25 dniach. Gdy pogoda jest wilgotna, askospory będą produkowane i uwalniane przez całą wiosnę i lato, zapewniając ciągłą infekcję pierwotną. (Wilcox, Wayne F. "Czarna zgnilizna Guignardia bidwellii." Karta identyfikacyjna choroby nr 102GFSG-D4. 2003. Cornell. 24 Oct. 2010).
Tak więc grzyb czarnej zgnilizny wymaga ciepłej pogody dla optymalnego wzrostu; chłodna pogoda spowalnia jego wzrost. Do infekcji wymagany jest również okres 2 do 3 dni deszczu, mżawki lub mgły." (Ries, Stephen M. "IPM : Reports on Plant Diseases : Black Rot of Grape." Integrated Pest Management at the University of Illinois. Dec. 1999. 24 Oct. 2010). Krople deszczu przenoszą te zarodniki, przenosząc je na różne części roślin, zwłaszcza na podatne młode liście. W przypadku obecności wody konidia kiełkują w ciągu 10-15 godzin i wnikają w młode tkanki. Nowe infekcje czarnej zgnilizny trwają do późnej wiosny i lata podczas dłuższych okresów ciepłej, deszczowej pogody. Konidia są zdolne do kiełkowania i wywoływania infekcji przez kilka miesięcy po ich uformowaniu. W sierpniu piknidia przekształcają się w stadium zimujące, które z kolei daje początek pseudotecjom, wewnątrz których wytwarzane są zarodniki wiosenne (askospory). Badania wykazały, że askospory są ważnym źródłem pierwotnych infekcji wiosną. Wiosną, podczas wilgotnej pogody, "piknidia na zainfekowanych tkankach wchłaniają wodę i konidia są wyciskane. Konidia są rozpryskiwane przez deszcz i mogą zainfekować każdą młodą tkankę w czasie krótszym niż 12 godzin w temperaturze 60-90 stopni. (Ellis, Michael A. "Fact sheet Agricultural and Natural Resources: Grape Black Rot." Departament Patologii Roślin. The University of Ohio State Extension. 2008).
Zastosowanie praktyczne
Dla naszego modelu podstawą jest literatura opublikowana przez Spottsa (Spotts. R. A. 1977. Effect of leaf wetness duration and temperature on the infectivity of Guignardia). Dzięki rozmowom z Danielem Molitorem zmodyfikowaliśmy oryginalny model Spottsa w kierunku modelu dzielącego się na lekkie, umiarkowane i ciężkie infekcje, podobnego do konstrukcji modeli parcha jabłoni. (Daniel Molitor: Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Dissertation Justus-Liebig-Universität Gießen, 2009).
Zakażenie spełniające "kryteria Spotts" jest oceniane jako zakażenie lekkie. Zakażenia spełniające "kryteria Spotts" wg 150% ocenia się jako umiarkowane, a zakażenia spełniające "kryteria Spotts" wg 200% jako ciężkie.
Model pokazuje terminy infekcji dla lekkich, umiarkowanych i ciężkich infekcji. Gdy 100% infekcji zostały spełnione warunki do wysypu zarodników, kiełkowania i infekcji tkanek roślinnych. Nasilenie infekcji zależy od okresu optymalnej temperatury i okresu wilgotności grzyba. W zależności od natężenia infekcji należy wykonać oprysk zapobiegawczy lub leczniczy w krótkim czasie po infekcji. W umiarkowanym, półsuchym klimacie Mozeli lub większości austriackich obszarów uprawy winorośli oprysk po stwierdzeniu lekkiej infekcji nie byłby zalecany.
Lobesia botrana (Totricidae, Olethreutinae) został po raz pierwszy opisany z Austrii, ale pochodzi z południowych Włoch i wyemigrował do całej Europy, północnej i zachodniej Afryki, Bliskiego Wschodu i wschodniej Rosji. Ostatnio został wprowadzony do Japonii, a w 2008 roku po raz pierwszy odnotowano go w Chile.
Gospodarze
Grape (Vitis vinifera) i wawrzynka wilczełyko (Daphne gnidium) są preferowanymi gospodarzami, ale odnotowano ją również na jeżynach (Rubus fruticosus), agrest (Ribes sp.), porzeczka czarna i czerwona (Ribes nigurm), oliwka (Olea europaea), wiśniowy (Prunus avium), prune (Prunus domestica), persymona (Diospyrus kakis), kiwi (Actinidia chinensis), granat (Punica granatum), goździk (Dianthus spp.), oraz szereg innych dzikich gospodarzy.
Szkody
W maju i czerwcu larwy pierwszego pokolenia tworzą sieć i żerują na gronach kwiatowych. Larwy drugiego pokolenia (lipiec-sierpień) żerują na zielonych jagodach. Młode larwy wnikają do wnętrza jagody i drążą je, pozostawiając skórkę i nasiona. Największe szkody wyrządzają larwy trzeciego pokolenia (sierpień-wrzesień), które żerują wewnątrz jagód i w kiściach, które zostają zanieczyszczone odchodami. Dodatkowo uszkodzenia jagód przez żerujące larwy sprzyjają infekcji przez Botrytis i inne grzyby wtórne, np. Aspergillus, Alternaria, Rhizopus, Cladosporium, oraz Penicillium.
Biologia i cykl życia
Dorosła ćma ma około 6-8 mm długości, a rozpiętość skrzydeł wynosi 11-13 mm. Samica jest nieco większa. Zarówno samce jak i samice mają podobne skrzydła o mozaikowym wzorze. Pierwsza para skrzydeł (przedplecze) jest kremowo-brązowa, cętkowana szaro-niebieskimi, brązowymi i czarnymi plamami. Druga para skrzydeł jest szara z frędzlami. Skrzydła są trzymane w kształcie dzwonu nad odwłokiem, gdy w spoczynku. Samice składają jaja oddzielnie. Jaja są eliptyczne i płaskie, o średnicy około 0,6-0,8 mm. Te soczewkowate jaja są widoczne i opalizujące kremowobiałe, w miarę rozwoju zarodka zmieniają kolor na żółty, a później na czarny, gdy powstaje głowa rozwijającej się larwy. Wyróżnia się pięć stadiów larwalnych. Pierwsze stadium ma wielkość około 1 mm, a ostatnie około 12-15 mm. Larwy są z czarną głową, po czym zmieniają kolor na żółtobrązowy, a ciało przyjmuje barwę zgodną z odżywianiem (zawartość jelit). Piąty ostatni instar larwalny przędzie szarobiały silikonowy kokon w celu poczęcia. Poczwarka ma długość około 4-9 mm.
W naszej strefie klimatycznej europejska ćma winniczek ma dwa do trzech pokoleń rocznie. Z cieplejszych rejonów Hiszpanii, Grecji, Egiptu odnotowano cztery pokolenia. Stadium poczwarkowe zimuje wewnątrz silikonowych kokonów (diapauza) pod korą, w szczelinach gleby lub na ukrytych, chronionych miejscach. Gdy temperatura powietrza wzrośnie powyżej 10°C przez około 10-12 dni wylęgają się dorosłe osobniki. Najpierw pojawiają się samce, a po tygodniu wylęgają się samice. Tak więc pierwszy lot samców może rozpocząć się już w momencie pęknięcia pąka, a lot trwa przez około 4 do 5 tygodni. Aktywność lotna przypada na czas wschodu słońca i temperatury około 12°C. Podczas lotu owady łączą się w pary, a większość samic łączy się w pary tylko raz. Składanie jaj rozpoczyna się wkrótce po kopulacji (około jednego do dwóch dni). Jaja składane są pojedynczo na kwiatach lub w ich pobliżu. Samica składa około 100 jaj, a dorosłe życie trwa od 1 do 3 tygodni w zależności od warunków klimatycznych.
Wiosną wylęganie jaj trwa 10 i więcej dni, natomiast latem jaja wylęgają się w ciągu 3 do 4 dni w zależności od warunków temperaturowych. Larwy pierwszego pokolenia tworzą sieć na kwiatach i żerują na nich, mogą wnikać w szypułkę i powodować zasychanie grona. Rozwój larw potrzebuje około 20-30 dni w zależności od warunków pogodowych. Następnie przepoczwarczają się pod korą, w glebie lub na kwiatach, a osobniki dorosłe pojawiają się ponownie 6 do 14 dni po.
Samice ćmy drugiego i trzeciego lotu składają jaja pojedynczo bezpośrednio na zacienionych jagodach. Wkrótce po wyłonieniu się larwa wchodzi na jagodę i żerując wydrąża ją. W jednym gronie może znajdować się kilka larw. Pajęczyny, odchody i infekcje grzybowe mogą powodować rozległe zanieczyszczenie grona.
Dolny próg rozwoju to około 10°C, a górny próg rozwoju to 30°C. Optymalne warunki rozwoju to 26-29°C i wilgotność od 40 do 70%. Jeśli długość dnia maleje i robi się coraz zimniej, rozpoczyna się stadium diapauzy. Stadia larwalne giną, gdy temperatura spada poniżej 8°C, ale poczwarka diapauzująca wytrzymuje nawet mroźne zimy północnej Europy.
Pierwsze pokolenie jest krótsze od pokoleń letnich. Jaja wylęgają się w około 66 stopniach Celsjusza (DDC). Larwy żerujące na gronach kwiatowych rozwijają się podobno szybciej niż te żerujące na jagodach winogron w późniejszym okresie sezonu, co wpływa na czas trwania pokolenia. Poczwarki, które nie diapauzują, potrzebują do rozwoju około 130 DDC. Dorosłe samice mogą składać jaja około 61 dni po pojawieniu się.
Szacunki DD dla jednego pokolenia różnią się znacznie w literaturze, od 427 DDC do 577 DDC w pierwszym pokoleniu do 482 DDC do 577 DDC w późniejszych pokoleniach.
Monitorowanie i zarządzanie
Samce wabione są przez feromony, co wykorzystuje się do monitorowania lotów samców. Przed pęknięciem pąków należy umieścić pułapki z L. botrana przynęty wysoko w koronach drzew (feromony iScout). Umieść co najmniej jedną pułapkę na 30 akrów lub na blok winnicy, jeśli jest mniejszy. Zmieniaj przynęty zgodnie z zaleceniami producenta. Sprawdzać pułapki co tydzień, zapisując liczbę złapanych motyli i usuwając je z lepkiego dna pułapki. Odnotować tygodniowe połowy w celu określenia początku i szczytu lotów samców w każdym pokoleniu. Kontynuować monitoring z użyciem pułapek do szczytu trzeciego lotu.
Stosowanie insektycydów powinno być zaplanowane na czas pojawienia się larw, dlatego też monitorowanie składania jaj i określanie stopnia ich wylęgania jest niezbędne w zwalczaniu tego szkodnika. W przypadku pierwszego pokolenia, składanie jaj powinno być monitorowane od szczytu do końca lotu. Należy szukać jaj na szypułkach 100 gron, wybierając jedno grono z każdej winorośli. Należy zwrócić uwagę na stadium większości znalezionych jaj. Jaja są białe, gdy zostały niedawno złożone, zmieniają kolor na żółty, a następnie czarny, gdy larwy są bliskie pojawienia się. Półprzezroczysta otoczka jaja wskazuje, że larwa już się pojawiła. Po wykluciu się jajka, szukajcie pajęczyny z części kwiatowych. Otwórz pajęczynę i poszukaj uszkodzeń spowodowanych żerowaniem i larw.
Rozpocząć monitoring jaj drugiego i trzeciego pokolenia na jagodach tydzień po złapaniu w pułapki pierwszych motyli z danego lotu. Kontynuować monitoring pod kątem obecności jaj co tydzień, aż do tygodnia po szczycie lotu. Skontrolować 100 kiści, wybierając jedną z każdej winorośli. Kontynuować monitorowanie kiści pod kątem uszkodzeń spowodowanych żerowaniem (dziury lub puste jagody), pajęczyn i obecności larw.
W krajach, w których L. botrana Środki zwalczania są ukierunkowane na drugie pokolenie. Wynika to częściowo z przedłużonego okresu pojawiania się pierwszego pokolenia oraz z możliwości ponownego zakażenia przez nieleczone sąsiednie winnice. Jednakże zwalczanie pierwszego pokolenia jest zalecane, jeśli populacja jest wysoka lub jeśli zabiegi przeprowadzane są na całym obszarze. Insektycydy są mniej skuteczne po zamknięciu kiści. Kilka insektycydów o zmniejszonym ryzyku jest zarejestrowanych do stosowania w winogronach w celu zwalczania larw mączlika. Należą do nich regulatory wzrostu owadów, spinosyny i Bacillus thuringiensis.
Zakłócanie kojarzenia jest badane w Europie od kilku lat. Okazało się, że jest ono najbardziej skuteczne, gdy populacja motyli jest niska i gdy stosuje się je na dużych obszarach o powierzchni ponad 10 akrów lub na całym obszarze.
W literaturze europejskiej podawane są liczne drapieżniki i parazytoidy. Wśród parazytoidów są 4 gatunki muchówek tachinidalnych i blisko 100 gatunków pasożytniczych os z rodzin ichneumonidów, braconidów, pteromalidów i chalicydów. Pasożyty, które według doniesień powodują największe szkody, to te atakujące zimujące poczwarki. W Hiszpanii są to pteromalidy Dibrachys affinis oraz D. cavusktóre według doniesień powodują śmiertelność poczwarek do 70%, natomiast we Włoszech ichneumonidy Dicaelotus inflexus oraz Campoplex capitator są najważniejsze.
Literatura:
- Briere JF, Pracros P. 1998. Porównanie modeli wzrostu zależnych od temperatury z rozwojem Lobesia botrana (Lepidoptera: Tortricidae). Environ. Entomol. 27: 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. en las comarcas vitícolas valencianas. Bol. Serv. Plagas 7: 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). In Los parasitos de la vid, estrategias de proteccion razonada. Madrid, Spain. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33: 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. Study of the relationship between sex pheromone trap catches of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) and the accumulation of degree-days in Sherry vineyards (SW of Spain). J. Appl. Ent. 125: 9-14.
- Gabel B, Mocko V. 1986. Symulacja funkcjonalna europejskiej ćmy winorośli Lobesia botrana Den. Et Schiff. (Lep., Torticidae) rozwoju populacji. J. Appl. Ent. 101: 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. Forecasting the flight activity of Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) in Southwestern Spain. J. Appl. Entomol. 133: 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. Control of Sparganothis pilleriana Schiff. and Lobesia botrana (Den. & Schiff). in German vineyards using sex pheromone-mediated mating disruption. Bull. IOBC/WPRS 25: 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera: Tortricidae): influence de l'infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thése N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. Daphne gnidium, możliwa rodzima roślina żywicielska europejskiej ćmy winorośli Lobesia botrana, stymuluje jej owipozycję. Czy zmiana żywiciela ma znaczenie? Chemoecol. 16: 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. Attraction of the grapevine moth to host and non-host plant parts in the wind tunnel: effects of plant phenology, sex, and mating status. Entomol. Exp. Appl. 122: 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Ent. 125: 515-518.
- Moreau J, Benrey B, Thiéry. 2006. Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 96: 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. Effects of methoxyfenozide on Lobesia botrana (Den & Schiff) (Lepidoptera: Torticidae) egg, larval and adult stages.
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. In Les ravageurs de la vigne. Féret, Bordeaux. pp. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. The relation between dispersal and survival of Lobesia botrana larvae and their density in vine inflorescences. Entomol. Exp. Appl. 84: 109-114.
- Xuéreb A, Thiéry D. 2006. Czy naturalne pasożytowanie larw Lobesia botrana (Lepidoptera: Torticidae) różni się w zależności od roku, pokolenia, zagęszczenia żywiciela i cultrivar winorośli? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). In Lepidotteri dei fruttiferi e della vite. Milan, Italy. Pp. 85-88.
Modelowanie europejskiego mola winogronowego w platformie FieldClimate
Potrzebny czujnik: Temperatura powietrza
Dane godzinowe Wyjście: aktywność lotnicza, aktywność w zakresie składania jaj, pierwsze pokolenie, drugie pokolenie i trzecie pokolenie, skumulowana liczba stopniodni
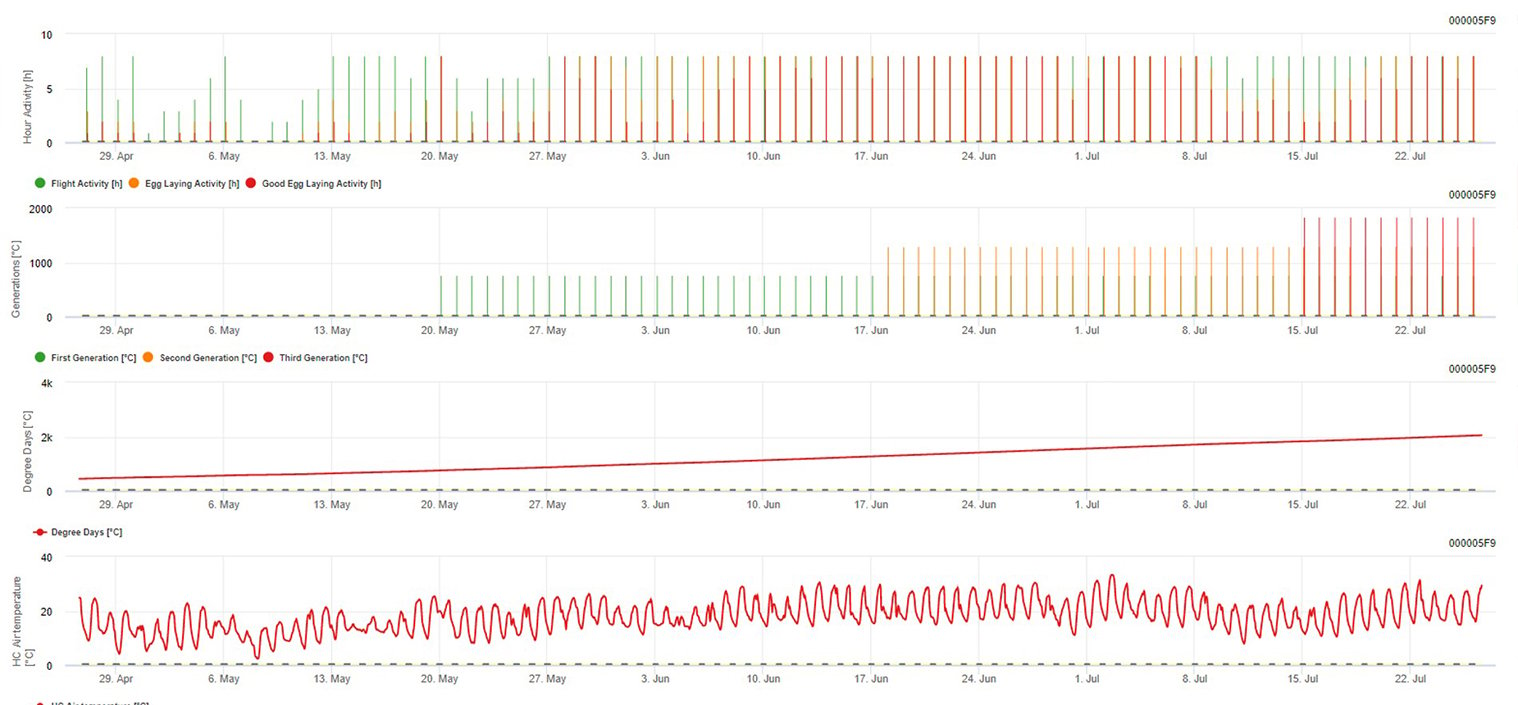
Za aktywność w locie, aktywność w znoszeniu jaj i dobre znoszenie jaj: Sprawdzamy dane z ostatnich 8 godzin (między 19:00 a 03:00). Próg dla aktywności lotowej wynosi >12°C, dla aktywności w składaniu jaj > 15°C, a dla dobrej aktywności w składaniu jaj > 17°C. Jeśli wszystkie warunki dla aktywności składania jaj zostały spełnione, generujemy pierwsze pokolenie (= skumulowana liczba stopniodni). Pierwsze pokolenie +530 DD to drugie pokolenie (jeśli nie jest przewidywane z powodu wcześniejszych warunków). Drugie pokolenie + 530 DD to trzecie pokolenie.
Na wykresie widać występowanie pierwszego pokolenia (dobre warunki dla wszystkich trzech aktywności lotu, składania jaj i dobrego składania jaj) na około 469 stopniodni skumulowanych. Lot drugiego pokolenia rozpoczyna się w połowie czerwca na około 1010 stopniodni skumulowanych i dobrych warunkach temperaturowych do składania jaj.
Botrytis cinerea jest nekrotroficznym grzybem, który atakuje wiele gatunków roślin, choć jego najbardziej godnymi uwagi gospodarzami mogą być winogrona. W winiarstwie jest powszechnie znany jako zgnilizna pęczkowa botrytis; w ogrodnictwie jest zwykle nazywany szarą pleśnią lub szarą pleśnią.
Grzyb ten powoduje dwa różne rodzaje infekcji na winogronach. Pierwszy z nich, szara zgnilizna, jest wynikiem stale wilgotnych lub mokrych warunków i zazwyczaj prowadzi do utraty dotkniętych nią kiści. Drugi rodzaj, szlachetna zgnilizna, pojawia się, gdy po wilgotnych warunkach następują bardziej suche, a jego efektem mogą być charakterystyczne słodkie wina deserowe, takie jak Sauternes czy Aszú z Tokaju. Nazwa gatunkowa Botrytis cinerea pochodzi od łacińskiego "winogrona jak popiół"; choć poetyckie, "winogrona" odnosi się do zgrupowania zarodników grzyba na ich konidioforach, a "popiół" po prostu odnosi się do szarawego koloru mas zarodników. Grzyb określany jest zwykle nazwą anamorf (forma bezpłciowa), ponieważ faza płciowa jest rzadko obserwowana. Teleomorfem (formą płciową) jest ascomycete, Botryotinia cinerea.
Biologia B. cinerea
W przypadku owoców i winogron infekcje kwiatowe, po których następuje okres utajenia, mają duży wpływ na epidemiologię szarej pleśni. Można wyróżnić kilka dróg infekcji od kwiatu do owocu. W winogronach, kiwi, malinach postuluje się infekcje przez trzonek do zalążni. W zalążni patogen pozostaje w stanie latencji, co wydaje się być wynikiem wstępnie ukształtowanej strategii obronnej gospodarza (podobnie jak zawartość resweratrolu w młodych jagodach winogron). W winogronach, kiwi i truskawkach istotne są infekcje przez pręcik, płatki lub przegrody. W winogronach badania wykazały, że B. cinerea może infekować pręciki i rozrasta się podstawowo, aby zainfekować zawiązek, a następnie rozrasta się systemowo do szypułki i tkanek naczyniowych w jagodach.
Sześcioletni projekt badawczy w rejonie Cape wykazał, że jagody winogron mogą być zakażone przez stomię i przetchlinki szypułki oraz w dolnej części rachityki. Infekcje szypułki są możliwe także w okresie kwitnienia. Później ta tkanka zwiększa odporność na B. cinerea zakażenia.
Inne drogi infekcji postulują saprofityczny rozwój patogenu na resztkach kwiatowych i późniejsze zakażenie jagód, gdy podatność wzrasta wraz z dojrzewaniem lub w wyniku uszkodzenia jagód przez owady lub grad. Inną tezą jest założenie o gromadzeniu się konidiów wewnątrz owoców w okresie letnim i zakażaniu podatnych jagód w późniejszym okresie sezonu. Zakażenie konidialne dojrzewających owoców jest możliwe z każdego źródła inokulum. Najprawdopodobniej powstaje niewielka liczba utajonych zakażonych jagód, które wykazują rozległą sporulację, gdy podatność jagód wzrasta wraz z dojrzałością. Można przyjąć, że jagody stają się podatne od zawartości cukru 6%.
W owocach kiwi mamy duży wpływ warunków zbioru na występowanie B. cinerea. Owoce zbierane z mokrą powierzchnią mogą zostać zainfekowane przez B. cinerea na mikro zmiany ustawione przez palce zbieraczy.
W praktycznej kontroli B. cinerea musimy wydzielić dwa ważne okresy infekcji: Kwitnienie i starość. 1) Podczas kwitnienia mamy podatne młode owoce, gdzie po infekcji następuje okres latencji. 2) Podczas gdy infekcja na dojrzałych (starzejących się) owocach prowadzi do objawów bez okresu latencji. Znaczenie infekcji podczas kwitnienia w winogronach może się zmieniać w zależności od sezonu i regionu. W przypadku owoców, na których musimy spodziewać się pewnej trwałości (winogrona stołowe, kiwi czy truskawki) objawy widoczne są podczas przechowywania w warunkach chłodniczych w sklepach lub magazynach. Chemiczne zwalczanie winogron wykazujących dobrą odporność na B. cinerea w czasie kwitnienia nie przyniesie żadnych efektów ekonomicznych. Dlatego przy podejmowaniu decyzji o zastosowaniu przeciwko należy wziąć pod uwagę wszystkie warunki ryzyka i prawdopodobieństwa infekcji, podatność owoców oraz okres trwałości, warunki przechowywania. Botrytis cinerea podczas kwitnienia.
W owocach pestkowych zakażenie przez B. cinerea występuje najczęściej w okresie kwitnienia. W tym czasie zabiegi przeciwko Monilina spp. są brane pod uwagę, które również infekują Botrytis cinerea infekcja.
Model B. cinerea i praktyczne zastosowanie
Potrzebne czujniki: Wilgotność liści, temperatura, wilgotność względna
Botrytis cinerea jest fakultatywnym pasożytem. Rośnie również na martwym materiale roślinnym. Z tego powodu jest zawsze obecny w winnicach i sadach. Botrytis cinerea jest związana z wilgotnym klimatem. Do infekcji potrzebuje bardzo wysokiej wilgotności względnej lub obecności wolnej wody (czujnik: wilgotność liści, wilgotność względna). Grzyb nie jest w stanie zainfekować zdrowego, dorosłego materiału roślinnego przez zarodniki. Infekcja następuje na młodych pędach winorośli podczas dłuższych okresów wilgotnych lub na pędach uszkodzonych przez burze gradowe.
Do obliczenia ryzyka infekcji model wykorzystuje następującą korelację pomiędzy czasem trwania zwilżenia liści a temperaturą.
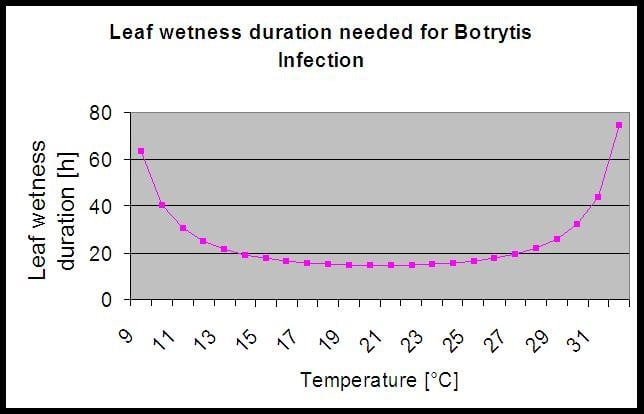
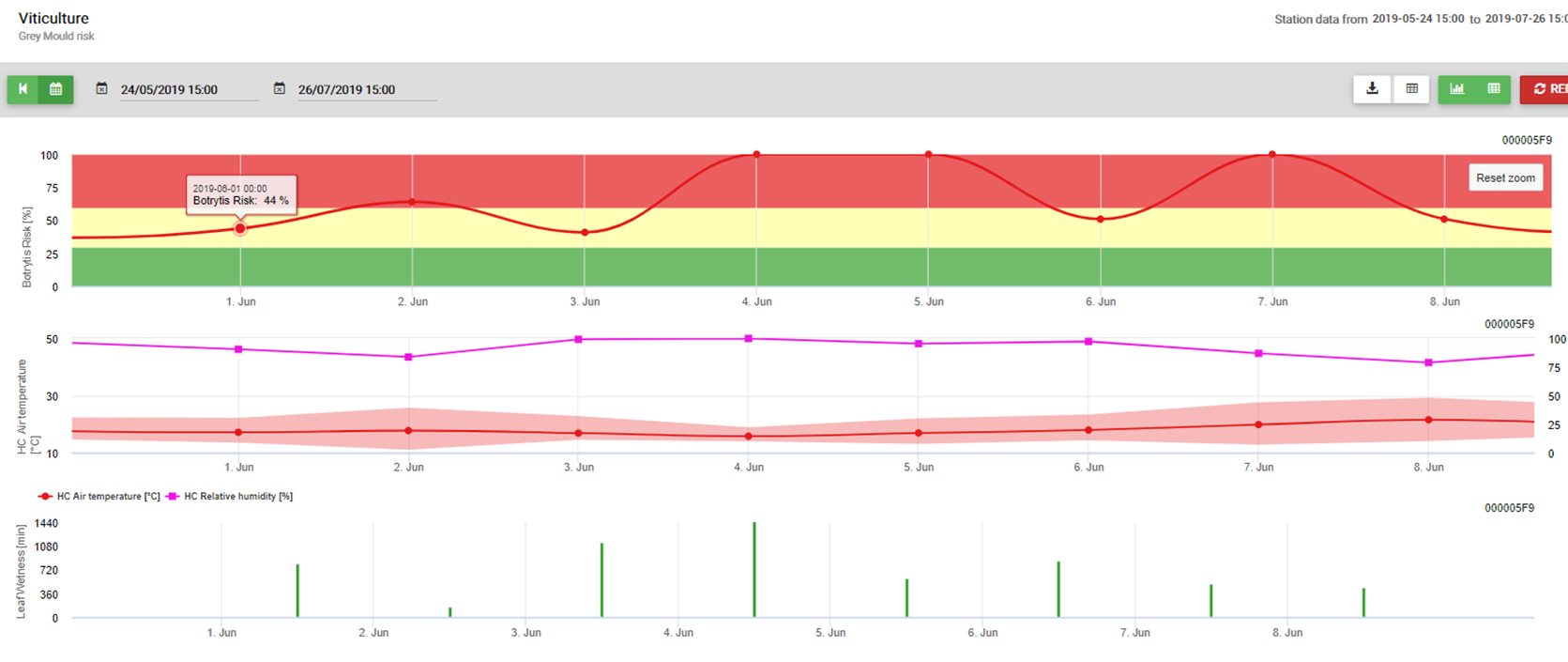
Na stronie Botrytis Model ryzyka daje w wyniku wartość ryzyka od 0 do 100%. Wartość ta wskazuje na ciśnienie B. cinerea w danym czasie. Jeśli mamy wartość 100% oznacza to, że kilkakrotnie wystąpił okres mokry na tyle długi, aby zainfekować podatną tkankę (obliczamy tzw. "punkty mokre" (macierz pomiędzy mokrością liścia, temperaturą z maksimum początkowo 38400 punktów (początek sezonu, który wykazuje ryzyko 30%). Po tym okresie każdy okres mokry z około 4000 punktów mokrych (macierz) zwiększa ryzyko o 10% lub z drugiej strony każdy okres suchy zmniejsza ryzyko o 1/5 poprzedniej wartości.
Wniosek przeciwko B. cinerea zależy od owoców i celu produkcji.
Antraknoza, czarna plamistość, zgnilizna ptasiego oka, ptasia plamistość występuje na całym świecie. Choroba obniża jakość owoców i plon, osłabia winorośl poprzez znaczne zniszczenie nowych pędów i liści. Większość gatunków Vitis jest podatna na antraknozę.
W wilgotnych sezonach uprawy winorośli choroba jest dobrze rozwinięta, powoduje uszkodzenia bardzo podatnych odmian po wczesnych deszczach.
Raz zadomowiona w winnicach choroba może być bardzo trudna do opanowania. Jednak w Europie choroba ta jest prawie nieistotna ze względu na stosowanie fungicydów ze związkami miedzi, ditiokarbaminianami, ftalimidami do zwalczania mączniaka prawdziwego oraz brak opadów w klimacie jałowym.
Symptomy
Choroba ta atakuje nadziemne, soczyste części winorośli, w tym młode pędy, liście, ogonki liściowe, rozłogi i grona; najczęściej występują zmiany na pędach i jagodach. Pierwsze objawy pojawiają się jako pojedyncze, małe, okrągłe plamy, które stają się brązowawe, zapadnięte z szarym środkiem i ciemnymi, okrągłymi lub kanciastymi brzegami. Czasami objawy przypominają uszkodzenia spowodowane gradem. Na liściach pojawiają się liczne, małe, brązowe plamy, które w środku są szare, środek nekrotyczny zwykle odpada, tworząc "dziurę po strzale". Młode liście są bardziej podatne na infekcje. Na jagodach pojawiają się małe czerwono-brązowe, okrągłe plamy, które stają się lekko zapadnięte. Środek staje się szary i jest otoczony czerwono-brązowymi do czarnych obwódkami (ptasie oko).
Cykl życiowy patogenu
Choroba jest wywoływana przez patogen Elsinöe ampelina (de Bary) Shear.
Zainfekowane pędy są głównym źródłem choroby. Przetrwałe w zmianach chorobowych skleroty lub grzybnia uaktywniają się wiosną i wytwarzają konidia w warunkach wilgotnych (deszcz lub rosa przez 24 godziny) w reżimie temperaturowym 2°C-40°C. Konidia są rozpryskiwane przez deszcz do nowych tkanek. Kiełkują i wytwarzają nowe infekcje. Ciepła pogoda skraca czas zwilżenia potrzebny do początkowej infekcji oraz okres inkubacji zanim pojawią się objawy na liściach. Optymalne kiełkowanie zarodników następuje w temperaturze 25-30°C, przy minimalnej wymaganej wilgotności liści wynoszącej 3- 4 godziny. Również czas inkubacji jest najkrótszy w tym zakresie temperatur (3-4 dni w warunkach idealnej wilgotności). Przy infekcji w temperaturze około 10°C objawy chorobowe potrzebują około 14 dni, aby być widoczne na liściach.
Konidia lub askospory powstałe na zainfekowanych jagodach, które zimują na winorośli lub podłodze, mogą również powodować infekcje pierwotne. Zarodniki infekują nowe liście, pędy, czułki, młode jagody i wytwarzają zmiany chorobowe w wilgotnych warunkach. Konidia te służą jako inokulum wtórne i są odpowiedzialne za kolejne infekcje w sezonie. Konidia są roznoszone przez krople deszczu i rozpraszane w winnicy.
Rozwój epidemii spowodowany jest podatnością tkanek winorośli, rozproszeniem konidiów oraz panującymi warunkami atmosferycznymi. Najważniejszym czynnikiem jest częstotliwość występowania okresów deszczowych.
Modelowanie w warunkach polowych
Rozwój zarodników zimujących: Temperatura: 2-40C°, wilgotność względna powyżej 90% lub wilgotność liści- gdy zarodniki są rozwinięte (100%) i nadal warunki wilgotności liści i temperatury infekcja zaczyna być obliczana. Rozwój zarodników (wilgotność względna poniżej 50%) i infekcja przestają być obliczane, gdy wilgotność jest zbyt niska. Nasilenie infekcji zależy od warunków wilgotnych (deszcz).