





A fuzáriumos fejfoltosságot vagy az apró szemű gabonafélék rücskösödését a következő gomba okozza Fusarium graminearum (Schwabe), bár Hollandiából és Közép-Európa más területeiről a F. culmorum mint a legelterjedtebb faj (Snidjers, 1989). Lengyelországban, F. culmorum, F. graminearum és F. nivale hasonló közepes vagy súlyos virulenciaszintet mutattak ki, míg a F. avenaceum közepesen vagy közepesen virulensnek bizonyult. A kórokozó azonosítására irányuló számos vizsgálatban azonban a kórokozót okozó szervezetet, akár 18 Fusarium spp. izolálták és azonosították (Mihuta-Grimm és Foster, 1989; Reis, 1985).
A cikket írta L. Gilchrist, H.J. Dubin A rühatka a meleg, nedves régiókban gyakori, ahol a virágzás egybeesik az esős időszakokkal. A betegség előfordulása az elmúlt tíz évben különböző okok miatt egyre nőtt. Talán a legfontosabb ok az, hogy megnőtt az a terület, ahol a búzát kukoricával vagy más gabonafélékkel váltogatják. További okok a talajvédelmi célú termesztési rendszerben bekövetkezett változások, valamint a búza termesztésének változása a hagyományos termesztésről a nedvesebb, nem hagyományos területekre (Gilchrist et al., 1997).
A fuzáriumos rühatka világszerte súlyos termésveszteséget okoz, amely akár az 50 százalékot is elérheti. Hasonlóképpen, Paraguayban az 1972-es és 1975-ös időjárási körülmények kedveztek a Fusarium- és Septoria-járványoknak, amelyek együttesen akár 70 százalékos veszteséget is okozhattak (Viedma, 1989). Lengyelország, Hollandia, az Egyesült Királyság, a volt Csehszlovákia, az Oroszországi Föderáció, Franciaország és Ausztria - néhány európai ország, amely a varasodás előfordulását jelentette. Az amerikai kontinensen Kanada, az Egyesült Államok, Mexikó, Guatemala, Brazília, Ecuador, Uruguay és Argentína területeit nagymértékben érinti a Fusarium (Ireta és Gilchrist, 1994).
A varasodás jelentős termés- és minőségi károkat, valamint mérgezéseket okozhat az állatokban és az emberekben (Ireta és Gilchrist, 1994; Baht és mtsai., 1989; Luo, 1988; Snidjers, 1989; Marasas és mtsai., 1988). Az Egyesült Államokban a varasodás okozta károkat 1993-ban több mint $1 milliárd USD-re, 1994-ben pedig $500 millió USD-re becsülték. Kínában a becslések szerint a varasodás akár 7 millió hektárt is érinthet, és 2,5 millió tonna gabona veszhet el a járványos években. A fuzáriás mikotoxinnal kapcsolatos emberi megbetegedéseket Kínából, Indiából és Japánból jelentettek, míg állatoknál a világ számos pontjáról jelentettek betegségeket (Dubin és mtsai., 1997).
Ma már világszerte léteznek a mikotoxinokra vonatkozó szabályozások (Van Egmond és Dekker, 1995). Sok országban azonban nem alkalmazzák a szabályozást, és sokan, különösen a vidéki emberek, mindenféle ellenőrzés nélkül fogyasztják a gabonát, akár gabonaszemek formájában, akár közvetett formában, a szennyezett gabonával etetett állatokból származó hús formájában. A 16.1. táblázat a sertésekre és baromfikra gyakorolt főbb hatásokat mutatja be, amelyeket az alábbiakban felsorolt fontosabb toxinok váltanak ki Fusarium fajok.
A fejés és az érés közötti nedves és meleg időjárási körülmények fokozzák a varasodás súlyosságát. A F. graminearum behatolási pontja a tüske, különösen a virágszervek. Ez hatással van a magkötésre és a szemtöltésre. A fertőzött tüskék gyorsan veszítenek klorofillt és fakó színűvé válnak. Később rózsaszínűvé vagy barackszínűvé válnak, különösen a levélkék tövénél és széleinél (41. tábla). Ha a környezeti feltételek kedvezőek maradnak, a fertőzés átterjed a szomszédos tüskékre, és egyes esetekben az egész tüskét megfertőzheti, beleértve a vesszőt és annak szárát is. Ha a fertőzés súlyos, a károsodott szemeket micéliummal borítják be, és rózsaszínű, pelyhes massza megjelenését veszik fel. Ha a betegség mértéke mérsékelt, a szemek fonnyadtak, kis tömegűek és fehéres színűek lehetnek (sírköves szemek).
Az egyes Fusarium fajok által termelt fontosabb toxinok által a sertésekre és baromfikra gyakorolt főbb toxikus hatások
| Mikotoxin | Klinikai tünetek |
| Zearalenon | Duzzadt, vörös szeméremtest; hüvelyprolapsus sertéseknél; szopós malacoknál a szeméremtest megnagyobbodása; termékenységi problémák. |
| Vomitoxin (dezoxinivalenol, DON) | Csökkent takarmányfelvétel és súlygyarapodás a sertéseknél, ha a DON-t >2 mg/kg takarmányt kaptak; hányás; a takarmány visszautasítása nagyon magas DON-koncentráció esetén (>20 mg/kg takarmány)a |
| Egyéb trichotecének T-2 toxin HT-2 toxin Diacetoxyescirpenol> | Mérgezőbb, mint a DON; csökkent takarmányfelvétel; hányás; bőr- és gyomor-bélrendszeri irritáció; neurotoxicitás; abnormális utódok; fokozott betegségérzékenység; vérzés. |
| Ocharatoxin | Elsősorban a vesék proximális tubulusait érinti sertésben és baromfiban; a vesék durván megnagyobbodtak és sápadtak; baromfiban a máj zsírosodik. |
amg/kg = milliomodrész (ppm).
Forrás: Trenholm et al., 1984.
Fusarium graminearum (42. tábla) a búzanövényeket minden növekedési szakaszban megtámadhatja, magonc-, szár- és gyökérrothadásos betegségeket okozva. Az elsődleges fertőzések aszkozospórákból vagy makrokonídiumokból származhatnak, amelyek a levélkékre és az extrudált porzókra rakódnak le. A 10° és 30 °C közötti hőmérséklet és a 95 százalék feletti relatív páratartalom 40-60 órán keresztül általában elegendő ahhoz, hogy a makrokonídiumok sikeresen megfertőzzék a tüskéket (Ireta, 1989).
Fusarium graminearum egyike azon kevés Fusarium olyan faj, amely szabadföldi körülmények között peritéciákat termel (43. tábla, 44. tábla). A peritéciák a Gibberella zeae nevű gomba ivaros stádiumát képviselik, és a búzaszemeken termelődnek. A peritéciumok fontos szerepet játszanak a kórokozó évről évre történő túlélésében (Khonga és Sutton, 1988), és az előző termés maradványaiban lévő micéliumokkal együtt léteznek, és így képezik a varasodás kezdeti inokulumforrását. A Kínában végzett kutatások kimutatták, hogy a peritéciák termelődéséhez a legalacsonyabb hőmérséklet 7 °C és 10 °C között, a legmegfelelőbb pedig 15 °C és 20 °C között van. A peritéciák termelődéséhez a talaj legalacsonyabb nedvességtartalma 50-60 százalék, a legmegfelelőbb pedig 70-80 százalék volt (Wang, 1997).
A betegség kialakulását meghatározó tényezők közé tartozik az éghajlat, az inokulumszint és a búza növekedési szakasza. A búzarozsda-járványok elsősorban az elsődleges inokulum mennyiségétől függenek, nem pedig a másodlagos inokulumtól. Az invázió főként a virágzás időszakában következik be. A másodlagos fertőzés konídiumokkal (45. tábla) a beteg tüskéknek a szántóföldön való megjelenése után következik be.
A növényi maradványok és a termesztési gyakorlatok fontos szerepet játszanak a F. graminearum és következésképpen járványok. A kukoricamaradványokkal borított szántóföldre vetett búzán a fertőzés kétszer-háromszor súlyosabb lehet (Teich és Nelson, 1984). Általában a kórokozó csak rizs-búza vetésforgóban lévő rizstáblákon és Kína száraz vetésű területein lévő kukoricaszárakon volt szaprofita (Wang, 1997). Ha a maradékot beszántják, a peritéciumok túlélése csökken, és ez csökkenti az elsődleges inokulumforrást (Reis, 1989).
Fusarium graminearum fakultatív parazita, és számos más fűfélén, köztük a közönséges gyomnövényeken és gabonaféléken (rozs, rizs, árpa és tritikálé) is kórokozó. Ha az olyan gyakorlatok, mint a nem gazdanövényekkel való vetésforgó vagy a növényi maradványok kezelése, önmagukban nem hatékonyak, kombinálva csökkenthetik az elsődleges inokulum forrását. A betegség elleni védekezés hatékonyan az integrált kezelésen alapul, beleértve a megfelelő agronómiai gyakorlatokat, a rezisztens vagy toleráns fajták használatát és a vegyszeres alkalmazásokat.
Fuzáriumos fejfoltosság biológiája
Alkalmi kórokozók
Az FHB-t a Fusarium nemzetségbe tartozó gombafajok okozzák. Az FHB-t leggyakrabban okozó faj a Fusarium graminearum (szexuális szakasz - Gibberella zeae). Ez a gomba ugyanaz, amely gyakran a kukoricaszár rothadásával is összefüggésbe hozható. Egy másik Fusarium az FHB-t okozó faj Fusarium culmorum. Mindkét F. graminearum és F. culmorum szintén okozhat gyökérrothadást a kis szemeknél. Az árpán két másik Fusarium fajok, F. poae és F. avenaceum, szintén okozhat magfoltosságot.
Túlélés és terjedés
A gomba megmarad és elszaporodik a fertőzött apró szemű gabonafélék és kukorica termésmaradványain. Nedves időjárás esetén a gomba spóráit a szél elfújja vagy a gabonafélék fejére szórja. A spórák származhatnak a terményből, vagy a környező terményekből, néha nagy távolságokra is elfújhatják őket. A búza és a durumnövények a virágzási (beporzási) időszaktól a magfejlődés kemény tésztás szakaszáig fogékonyak a fertőzésre. A kórokozó gomba spórái a virágzás idején a szabadon lévő porzókon landolhatnak, majd a magokba, a búzaszemekbe vagy a fej egyéb részeibe fejlődhetnek. A tavaszi árpa esetében, amely akkor virágzik, amikor a fej a csizmában van, a fertőzés a virágzás után a leggyakoribb, amint a fej áttöri a levélhüvelyt. A fertőzés bármelyik kultúrában folytatódhat egészen a szemérésig, amennyiben a szervezet(ek) számára kedvezőek a környezeti feltételek.
A fertőzéshez a legkedvezőbb feltételeket a magas páratartalom és a meleg hőmérséklet (75-85 Fahrenheit fok (24°C-30°C)) jelenti. A fertőzés azonban hűvösebb hőmérsékleten is előfordul, ha a magas páratartalom 72 óránál hosszabb ideig fennáll. A korai fertőzések levegőben terjedő spórákat termelhetnek, amelyek felelősek a betegség másodlagos terjedéséért, különösen akkor, ha a növény a késői kelés miatt egyenlőtlenül virágzik.
Mivel az FHB kialakulása a virágzástól (árpában a fej megjelenésétől) a magok fejlődéséig tartó időszakban a kedvező környezeti feltételektől függ, a betegség előfordulása és súlyossága évről évre változik. A legsúlyosabb termés- és minőségi veszteségekhez a következő tényezők kombinációja vezethet: a bőséges inokulum, a virágzás (árpában a fej megjelenése) és a magfejlődés közötti hosszabb vagy ismétlődő nedves és magas páratartalmú időszakok, valamint a nagyon fogékony fajták használata.
Fusarium head blight fertőzési modell
A búzán a Fusarium head blight komplexhez tartozó gombakórokozókról ismert, hogy a 20 °C és 30 °C közötti meleg hőmérséklet és a hosszú párás időszakok kedveznek nekik. A többnapos levélnedves időszakok korai látható szeptómákhoz vezetnek. A tünetek azonban hosszabb lappangási idő után is jelentkezhetnek, ha a fertőzést 18 órás vagy még rövidebb levélnedves időszak követi. A tünetek 15 °C-os mesterséges beoltás után is jelentkezhetnek.
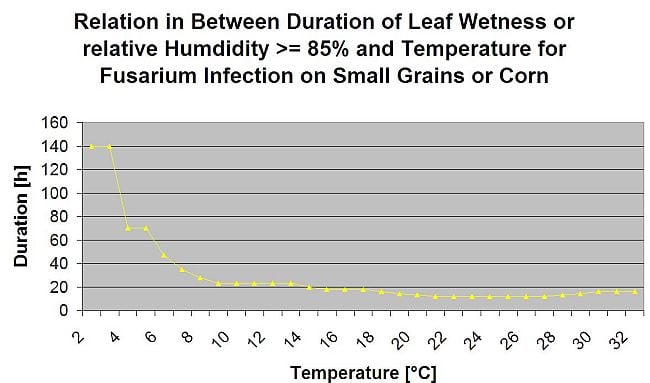

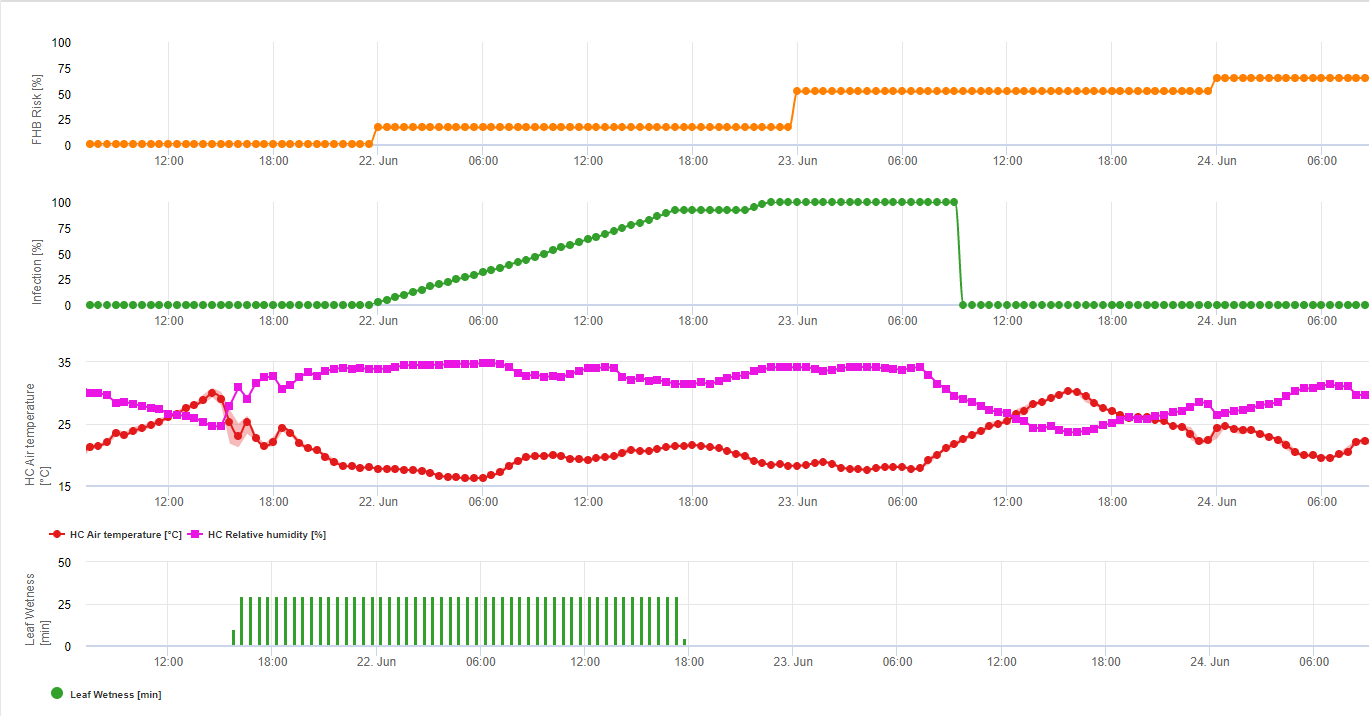
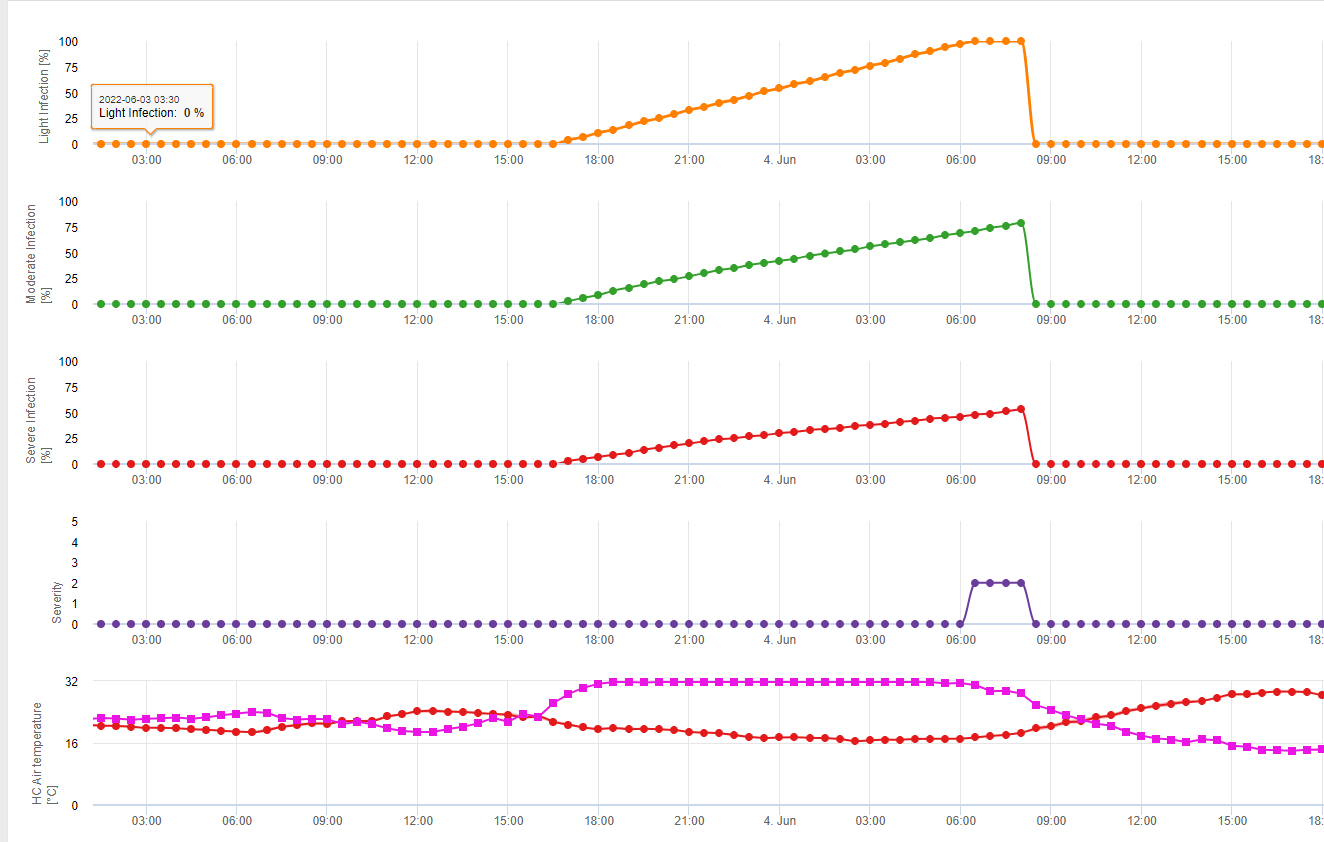
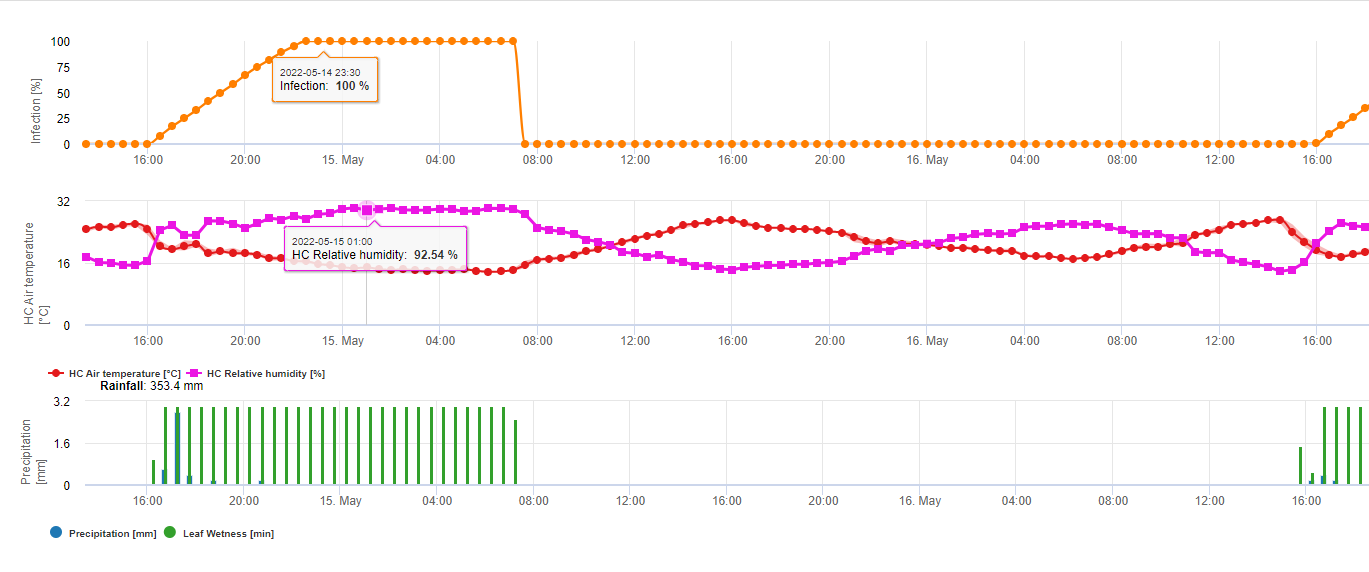
Összefoglalva a szakirodalomban talált különböző hőmérséklet- és nedvességkombinációkat, úgy döntöttünk, hogy a Fusarium head bligh fertőzésre akkor mutatunk rá, ha a hőmérséklet és a levélnedvesség, illetve a 85%-nél nagyobb relatív nedvességtartalmú időszakok meghaladják a következő grafikonon látható értékeket.
A fertőzéseket 2 mm-es eső indítja el. Fuzáriumos fejfoltosság fertőzésről akkor beszélhetünk, ha a fertőzés előrehaladási értéke eléri az 100% értéket. A fertőzés előrehaladási értékének kiszámítása a fenti grafikonon látható, a nedves körülmények időtartama és a hőmérséklet közötti összefüggést követi.
Ezt a modellt a fertőzéses napok és az éghajlati viszonyok megjelenítésére használják az éghajlat alatt. A termelők ismerete a különböző búzafajták fejlődési stádiumáról lehetőséget ad arra, hogy a fertőzés után azonnal döntsenek a gyógyító permetezés alkalmazásáról.
Fuzárium mikotoxin riasztás
A Fusarium fejfoltosság hosszan tartó levélnedvességgel járó fertőzési kísérletek magas mikotoxin-tartalomhoz vezettek. Ezen információk alapján a 61. és 69. stádiumban 48 órás vagy annál hosszabb levélnedvesedési időszak feltételezhetően magas mikotoxin kockázatot jelent. A DON-analízisek során a kereskedelemben termesztett búzában szerzett tapasztalatok azt mutatták, hogy a 61-69. stádiumban bekövetkező kezdeti fertőzést követő, a fertőzéshez elég hosszú levélnedvesedési időszakok a DON-értékeket is növelhetik. Hosszabb levélnedvesedési időszakok esetén a mikotoxinok mennyisége a 85. stádiumig növekedhet. Az FieldClimate a fertőzés előrehaladásával arányos kockázati értéket halmoz fel minden egyes sikeres fertőzési időszakon keresztül az e számításokhoz megfelelőnek választott időszak alatt. 6 éppen lezajlott fertőzési időszak 100% kockázatot eredményezne. A fuzáriumfertőzéshez vezető levélnedvesedési időszak általában hosszabb, mint a minimálisan szükséges. Ezért a legtöbb fuzáriumfertőzés 17%-nél nagyobb kockázatnövekedést eredményez. A problémás mikotoxin helyzetre utaló kockázati érték a szántóföldi előzményektől függ. A talajművelés nélküli kukorica után termesztett búza vagy a talajművelés nélküli kukorica után termesztett talajművelés nélküli búza csak akkor hordozhat kis kockázatot, ha nem az optimális helyzetben permetezik. A nem permetezett búzában már 35% kockázat után megnövekedett DON értékekkel kell számolnunk. A kukoricán vagy búzán kívüli egyéb kultúrát követő nem talajművelő búza után a búzánál magasabb, 50% kockázatot hordozhat. Ha kukorica vagy talajműveléses búza után van búza, akkor a kockázat 70%-ig terjedhet. Az elsőéves búzát DON-tesztnek kell alávetni, ha a kockázat meghaladja az 100% értéket.
1) Fuzáriumos fejfoltosság kockázati modell amely kiszámítja a fertőzés kockázatos időszakait. Amikor az 100% fertőzés (zöld vonal) elérte a kockázat (kék vonal) nagyon magas, és a gomba számára kedvezőek voltak a fertőzéshez szükséges feltételek. Az alkalmazási módtól függően (gyógyító, megelőző) a kockázatos időszakot a kék vonal mutatja.
2) Fuzáriumos fejfoltosság: ebben a modellben az FHB fertőzését a csapadék (2 mm szükséges), a relatív páratartalom (85% felett) vagy a levélnedvesség, a hőmérséklet alapján számítják ki a folyamat során. Ha a fertőzés eléri az 100% értéket, akkor a gombakórokozó számára optimális feltételek valósultak meg. A továbbiakban a modell kiszámítja az FHB mikotoxin kockázatát.
A búzában két fő Septoria-betegség létezik. Ezek a Septoria tritici foltosság, amelyet a gomba okoz. Septoria tritici (teleomorf: Mycophaerella graminicola), és Septoria nodorum a Septoria nodorum (teleomorf: Leptosphaeria nodorum) gomba által okozott foltosság. Mindkét betegség súlyos terméskiesést okoz, amely a jelentések szerint 31-53 százalék között mozog (Eyal, 1981; Babadoost és Herbert, 1984; Polley és Thomas, 1991). Világszerte több mint 50 millió hektárnyi, főként a csapadékban gazdag területeken termesztett búza érintett. Az elmúlt 25 év során ezek a betegségek egyre terjedtek, és bizonyos területeken a búzatermesztés egyik fő korlátozó tényezőjévé váltak. Súlyos járványok esetén a fogékony búzafajták magjai összezsugorodnak, és nem alkalmasak őrlésre. A Septoria tritici foltosság és a Septoria nodorum foltosság járványai kedvező időjárási körülményekhez (gyakori esőzések és mérsékelt hőmérséklet), különleges termesztési gyakorlatokhoz, az inokulum elérhetőségéhez és a fogékony búzafajták jelenlétéhez kapcsolódnak (Eyal et al., 1987).
Septoria spp. Biológia
Erick De Wolf után, Septoria Tritici Blotch, Kansasi Állami Egyetem, 2008. április Septoria tritici foltos levélfoltosság néven ismert foltosság, amelyet a gomba okoz. Septoria tritici. A világ valamennyi búzatermő vidékén elterjedt, és számos régióban komoly problémát jelent. A Septoria tritici foltosság akkor a legkártékonyabb, amikor a betegség a szezon végén a fogékony fajták felső leveleit és fejét támadja meg.
Tünetek
Septoria tritici a foltosság tünetei először ősszel jelennek meg. A kezdeti tünetek kis sárga foltok a leveleken. Ezek a sérülések az öregedés során gyakran világosbarnára színeződnek, és a gombák termőtestei a levélnyélen lévő sérülésekbe ágyazódva láthatók. A sérülések szabálytalan alakúak, és az elliptikustól a hosszú és keskenyig terjednek (1. ábra). A sérülések apró, kerek, fekete pöttyöket tartalmaznak, amelyek a gomba termőtestjei. A fekete termőtestek úgy néznek ki, mint a fekete borsszemek, és általában nagyító nélkül is láthatóak. A betegség az alsó leveleken kezdődik, és fokozatosan halad előre a zászlóslevél felé. A levélhüvelyek is fogékonyak a támadásra. Csapadékos években a pettyes levélfoltosság gomba átvándorolhat a fejekre, és barna elváltozásokat okozhat a levélkéken és a levélnyélen, az úgynevezett levélfoltosságot. Ezek az elváltozások az öregedés során gyakran világosbarnára színeződnek, és a gombák termőtesteit gyakran látni a levélnyélen lévő elváltozásokba ágyazódva.
A levélfoltosság fázisa jelentős terméskiesést okozhat, de a betegség súlyossága és a terméskiesés közötti kapcsolat nem jól ismert. Septoria tritici a foltosság összetéveszthető más levélbetegségekkel, amelyeknek nagyon hasonló tünetei vannak: a barnafoltosság és az Stagonspora nodorum foltos, például. Gyakori, hogy a növényeket több ilyen lombbetegség is megfertőzi, és laboratóriumi vizsgálatra lehet szükség annak pontos diagnosztizálásához, hogy melyik betegség a legelterjedtebb. Szinte mindig laboratóriumi vizsgálatra van szükség a levélfoltosság okának megkülönböztetéséhez. A faj ismerete nem fontos a permetezési döntések szempontjából, mivel mindhárom betegség hasonlóan reagál a fungicidekre. Azonban annak ismerete, hogy mely betegségek a legelterjedtebbek, fontos része a fajtaszelekciónak, mivel különböző gének szabályozzák a betegségekkel szembeni ellenállást.
A Septoria tritici foltosságot a legmegbízhatóbban a fekete termőtestek jelenléte alapján lehet megkülönböztetni a többi betegségtől. A barnafoltosságot okozó gomba nem termel ilyen típusú szaporodási struktúrát. A Stagonospora nodorum foltosságot okozó gomba azonban nedves körülmények között világosbarna termőtesteket hoz létre. A színkülönbségen kívül ezek a struktúrák kisebbek is, mint a Septoria tritici által termeltek.
Életciklus
Septoria tritici a nyarat átvészeli az előző búzatermés maradványain, és ősszel fertőzést indít. Bizonyos bizonyítékok vannak arra, hogy a gomba más fűgazdákkal és búzamagvakkal társulva is képes túlélni. A gomba ezen forrásai valószínűleg akkor a legfontosabbak, amikor a búzamaradványok hiányoznak. A vetésforgótól vagy a maradványkezelési gyakorlattól függetlenül általában elegendő inokulum áll rendelkezésre az őszi fertőzések elindításához. Septoria tritici a foltosságnak kedvez a hűvös, nedves időjárás. Az optimális hőmérsékleti tartomány 16 és 21 °C között van; a téli hónapokban azonban akár 5 °C-os hőmérsékleten is előfordulhat fertőzés. A fertőzéshez legalább 6 órán át tartó levélnedvesedés szükséges, és a maximális fertőzéshez akár 48 órán át tartó nedvesség is szükséges. A fertőzés után a gombának 21-28 napra van szüksége ahhoz, hogy kifejlődjenek a jellegzetes fekete termőtestek, és új spóratermést hozzon létre. Az ezekben a termőtestekben termelődő spórák ragacsos tömegben válnak ki, és esőre van szükségük ahhoz, hogy a felső levelekre és a fejekre fröccsenjenek.
Fertőzés Septoria tritici
Püknidiospórák a S. tritici szabad vízben csíráznak a spóra mindkét végéből vagy az interkaláris sejtekből (Weber, 1922). A spórák csírázása csak a levéllel való érintkezés után kb. 12 órával kezdődik. A csíracsövek véletlenszerűen nőnek a levél felületén. Weber (1922) csak az epidermális sejtek közötti közvetlen behatolást figyelte meg, mások azonban arra a következtetésre jutottak, hogy a nyitott és zárt sztómákon keresztül történő behatolás a gazdaszervezetbe való behatolás elsődleges módja (Benedict, 1971; Cohen és Eyal, 1993; Hilu és Bever, 1957). Kema és munkatársai (1996) csak sztómás penetrációt figyeltek meg. A sztómákon keresztül növekvő hifák körülbelül 1 μm átmérőre szűkülnek, majd a szubszomatális üreg elérését követően kiszélesednek.
A levél felszínével párhuzamosan, a levél epidermális sejtjei alatt, majd a mezofillumon keresztül az alsó epidermisz sejtjeihez nőnek, de nem az epidermiszbe. Nem képződnek haustóriumok, és a hifák növekedését az érkötegek körüli szklerenchimasejtek korlátozzák, kivéve, ha a hifák nagyon sűrűek. Az érkötegek nem hatolnak be. A hifák intercellulárisan nőnek a sejtfalak mentén a mezofillumon keresztül, a szeptumnál vagy a sejt közepén elágazva. Körülbelül 9 napig nem jelentkeznek makroszkópos tünetek, kivéve egy-egy elhalt sejtet, de a mezofill sejtek 11 nap után gyorsan elpusztulnak. A piknídiumok a szubszomatális kamrákban fejlődnek. A hifák ritkán nőnek be a gazdasejtekbe (Hilu és Bever, 1957; Kema et al, 1996; Weber, 1922).
A sikeres fertőzés csak legalább 20 óra magas páratartalom után következik be. Csak néhány barna folt alakult ki, ha a levelek a spórák lerakása után 5-10 órával (Holmes és Colhoun, 1974) vagy akár 24 órán át (Kema és mtsai., 1996) nedvesek maradtak. A gazdatest-parazita viszony a rezisztens és a fogékony búzákon azonos. A spórák csírázása a levélfelületen a fogékonyságtól függetlenül azonos. A sikeres behatolások száma nagyjából azonos, de a fogékony fajtákban gyorsabb a levéltetvek növekedése, ami több elváltozást eredményez. A hifák minden kultúrában a nekrotikus területen túlra terjednek 44 2. ülésszak - B.M. Cunfer. A patogenezisben szerepet játszhat egy toxin (Cohen és Eyal, 1993; Hilu és Bever, 1957). Ezzel szemben egy rezisztens vonalon a kolonizáció jelentősen csökkent (Kema és mtsai., 1996).
A gabonafélék Stagonospora (Septoria) és Septoria kórokozói: A fertőzési folyamat
B.M. Cunfer, Növénykórtani Tanszék, University of Georgia, Griffin, GA
A fertőzés folyamata legintenzívebben a Stagonospora (Septoria) nodorum és a Septoria tritici. Egy mélyreható tanulmány a Septoria passerinii rendelkezésre áll. A bejelentett információk szinte mindegyike a piknidiospórák általi fertőzésre vonatkozik. A fertőzés folyamata azonban más spóraformák esetében meglehetősen hasonló. A bemutatott információk többnyire a levelek optimális körülmények közötti fertőzésére vonatkoznak. Néhány vizsgálatot ép csíranövényekkel, míg másokat leválasztott levelekkel végeztek. A búza koleoptiliumának és csíranövényének fertőzése a S. nodorum Baker (1971) részletesen leírta, és Cunfer (1983) felülvizsgálta. Bár pontos összehasonlításokat nem végeztek, úgy tűnik, hogy a fertőzési folyamat sok hasonlóságot mutat az egyes gazdatest-parazita rendszerekben, és számos nekrotróf kórokozóra jellemző. A tünetek kialakulását és a betegség kifejeződését befolyásoló tényezőkről nincs információ, de más szerzők már áttekintették őket (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). A levélfelületen lévő spórák élettartamát befolyásoló tényezők összefoglalása szerepel.
A Cirrus szerepe és a spórák túlélése a levélfelületen A piknídiumból kinyert piknídiumspórákat burkoló cirrus funkciójáról a legrészletesebb információk a következőkre vonatkoznak S. nodorum. A cirrus egy fehérje- és szacharidvegyületekből álló gél. Összetétele és funkciója hasonló a Sphaeropsidales más gombákéhoz (Fournet, 1969; Fournet és mtsai., 1970; Griffiths és Peverett, 1980). A cirrus komponensek elsődleges szerepe a piknidiospórák védelme a kiszáradástól és a korai csírázás megakadályozása.
A cirrus védi a piknidiospórákat, így néhányuk legalább 28 napig életképes marad (Fournet, 1969). Ha a cirrus oldatot vízzel hígították, és a cirrus oldat koncentrációja >20% volt, a piknidiospórák kevesebb, mint 10% csírázott ki. Alacsonyabb koncentrációban az összetevők olyan tápanyagokat biztosítanak, amelyek serkentik a spórák csírázását és a csíracsövek megnyúlását. A csíracsövek hossza 15% cirrus koncentrációig nőtt, majd magasabb koncentrációknál mérsékelten csökkent (Harrower, 1976). Brennan és munkatársai (1986) híg cirrus folyadékban nagyobb csíraképződésről számoltak be. A cirrus komponensek 10-60% relatív páratartalomnál csökkentették a csírázást. Miután a spórák szétszóródtak, a cirrus folyadék stimuláló hatása valószínűleg elhanyagolható (Griffiths és Peverett, 1980).
35-45% relatív páratartalom mellett a spórák a S. tritici a cirriben legalább 60 napig életképes maradt (Gough és Lee, 1985). A cirri összetevői a spórák csírázásának gátlójaként hathatnak, vagy a cirri magas ozmotikus potenciálja megakadályozhatja a csírázást. A püknidiospórák a S. nodorum nem maradtak életben 24 órán keresztül 80% feletti relatív páratartalom mellett 20 C-on. A spórák két hétig vagy tovább éltek <10% relatív páratartalom mellett (Griffiths és Peverett, 1980). Amikor a cirrus folyadék a S. nodorum vízzel hígították, a piknidiospórák mintegy kétharmada 8 órán belül elvesztette életképességét, és 30 óra nappali fényben töltött idő után csak 5% csírázott. Amikor a spórákat sötétben tárolták, 40% 30 óra elteltével is életképes maradt (Brennan és mtsai., 1986).
Száraz konídiumok S. nodorum, árnyékban és közvetlen napfényben legalább 56 órán át éltek a szabadban (Fernandes és Hendrix, 1986a). A S. nodorum piknídióspórák csírázását folyamatos UV-B sugárzás (280-320 nm) gátolta, míg a S. nodorum piknídióspórák csírázását a S. tritici nem volt. A csíracsövek kiterjedése folyamatos UV-B sugárzás mellett mindkét gomba esetében gátolt volt a sötétben történő növekedéshez képest (Rasanayagam és mtsai., 1995).
Fertőzés Septoria nodorum által
A gazdaszervezet behatolásának és fejlődésének folyamata S. nodorum a levélen belül több kutató is részletesen vizsgálta (Baker és Smith, 1978, Bird és Ride 1981, Karjalainen és Lounatmaa, 1986; Keon és Hargreaves, 1984; Straley, 1979; Weber, 1922). A piknidiospórák általában két epidermális sejt közötti mélyedésekben rakódnak le, és sok levélbehatolási kísérlet ott kezdődik. A spórák a szabad nedvesség hatására a levél felszínén csíráznak (Fernandes és Hendrix, 1986b). Csírázásuk a lerakódás után 2-3 órával kezdődik, és 8 óra elteltével a csírázóképesség elérheti a 90%. A levélpenetráció körülbelül 10 órával a spórák lerakódása után kezdődik (Bird és Ride, 1981; Brönnimann és mtsai., 1972; Holmes és Colhoun, 1974).
A csírázás kezdetén a csíracsövet egy amorf anyag veszi körül, amely a levélhez tapad. A spóra mindkét végéből és az interkaláris sejtekből kinövő csíracsövek általában a sejtek közötti mélyedések mentén nőnek, és gyakran a levél hossztengelye mentén helyezkednek el (O'Reilly és Downes, 1986). A nem a mélyedésekben lévő spórákból származó hifák véletlenszerűen nőnek, esetenként elágazással (Straley, 1979). Appresszórium képződik egy fertőzési pálcikával, amely áthatol az epidermális sejtek kutikuláján és periklinális falán közvetlenül a sejtlumenbe, ami gyors sejthalált eredményez.
Sok behatolás először szubkutikuláris vagy oldalsó növekedés történik a sejtfalon belül, mielőtt a növekedés a citoplazmába történik (Bird és Ride, 1981; O'Reilly és Downes, 1986). A nyitott és zárt sztómákon keresztül történő behatolás is előfordul, és gyorsabb lehet, mint a közvetlen behatolás (Harrower, 1976; Jenkins, 1978; O'Reilly és Downes, 1986; Straley, 1979). A csíracsövek a sztómáknál és az epidermális sejtek csomópontjainál ágaznak el. A csíracső behatolása egy sztómába appresszorium kialakulása nélkül is megtörténhet. A behatolás néha a trichomákon keresztül történik (Straley, 1979). Úgy tűnik, a legtöbb behatolási kísérlet sikertelen, a behatolási kísérlet helyén a sejtekben sűrű papillák képződnek (Karjalainen és Lounatmaa, 1986; Bird és Ride, 1981).
A behatolás után az epidermális sejtek gyorsan elhalnak és elszarusodnak, a hifák pedig a mezofillumba nőnek. A mezofill sejtek eldeformálódnak, és a lignifikálódott anyag néhány sejten kívül rakódik le, amelyek ezután összeesnek. A lignifikáció még azelőtt bekövetkezik, hogy a hifák elérnék a sejtet. A folyamat minden fajtában ugyanúgy zajlik, de az ellenálló fajtákban lassabban fejlődik. A hifák intercellulárisan nőnek az epidermális sejtek között, majd a mezofillumba. A mezofillumba való behatoláskor a kloroplasztiszok pusztulása 6-9 nap alatt kezdődik (Karjalainen és Lounatmaa, 1986).
A fotoszintetikus sebesség azonban a fertőzést követő egy napon belül csökkenni kezd, mielőtt a tünetek láthatóvá válnának (Krupinsky és mtsai., 1973). Az érkötegek körüli szklerenchimaszövet megakadályozza az érszövet fertőzését. Az érkötegek megakadályozzák a hifák terjedését a mezofillumon keresztül, kivéve, ha a szklerenchimaszövet fiatal és nem teljesen kialakult (Baker és Smith, 1978).
A Stagonospora nodorum sejtfalbontó enzimek széles skáláját szabadítja fel, beleértve az amilázt, pektin-metilészterázt, poligalakturonázt, xilanázt és cellulázt in vitro és a búzalevelek fertőzése során (Baker, 1969; Lehtinen, 1993; Magro, 1984). Az enzimek által végzett sejtfallebontással kapcsolatos információk megegyeznek a szövettani megfigyelésekkel. ezek az enzimek toxinokkal együtt is működhetnek. Az enzimérzékenység összefügghet a rezisztenciával és a gombakolonizáció sebességével (Magro, 1984). Sok nekrotrófhoz hasonlóan a Septoria és Stagonospora kórokozók is termelnek fitotoxikus vegyületeket in vitro. A sejtek károsodása és elhalása a mezofillszövetbe történő hipófális növekedés előtt (Bird és Ride, 1981) összhangban van a toxintermeléssel. A toxinoknak a fertőzési folyamatban betöltött végleges szerepét és a gazdaszervezet rezisztenciájával való kapcsolatát azonban nem állapították meg (Bethenod és mtsai, 1982; Bousquet és mtsai, 1980; Essad és Bousquet, 1981; King és mtsai, 1983). A búzára és árpára adaptált törzsek közötti gazdanövénytartománybeli különbségek a S. nodorum a toxintermeléssel függhet össze (Bousquet és Kollmann, 1998). A spórák csírázásának megindulását és a csírázott spórák százalékos arányát nem befolyásolja a gazdaszervezet fogékonysága (Bird és Ride, 1981; Morgan 1974; Straley, 1979; Straley és Scharen, 1979; Baker és Smith, 1978).
Bird és Ride (1981) arról számolt be, hogy a csíracsövek kiterjedése a levélfelületen lassabban terjedt a rezisztens, mint a fogékony fajtákon. Ez a mechanizmus, amely legalább 48 órával a spórák lerakódása után fejeződik ki, a csíracsövek megnyúlásának behatolás előtti rezisztenciájára utal. A rezisztens fajtákon kevesebb sikeres behatolás történt, és a behatolás lassabban haladt a rezisztens fajtákon (Baker és Smith, 1978; Bird és Ride, 1981). A lignifikáció a fertőzés korlátozására javasolták mind a rezisztens, mind a fogékony kultúrákban, de más tényezők lassították a gomba fejlődését a rezisztens vonalakban. A fogékony vonalakon a gyorsabban növekvő hifák elkerülhetik a gazdasejtek lignifikációját.Négy nappal az árpa beoltása után egy búza biotípusú izolátummal történő beoltás után az S. nodorum, a hifák a kutikulán és néha az epidermális sejtfalak külső cellulózrétegén keresztül nőttek. A penetrációs hifák alatt vastag papillák rakódtak le, és a sejtekbe nem hatoltak be (Keon és Hargreaves, 1984).
Fertőzés Septoria passerinii által
Green és Dickson (1957) részletes leírást ad a fertőzés folyamatáról a S. passerinii árpán. A fertőzés folyamata hasonló a S. tritici. A S. triticihez hasonlóan a levélbe való behatoláshoz szükséges idő jelentősen hosszabb, mint a S. tritici esetében. S. nodorum. A csíracsövek véletlenszerűen ágaznak el és nőnek a levél felszínén, de néha az epidermális sejtek közötti mélyedések mentén is. A levélbe való behatolás szinte kizárólag a sztómákon keresztül történik. A csírázó hifák megduzzadnak, és ha a behatolás sikertelen, a hifák tovább nyúlnak. A spórák lerakódása után 48 órával nem történik behatolás. 72 óra elteltével a csíracsövek megvastagodnak a sztómák fölött, az őrsejtek között és a járulékos sejtek urfáján, valamint a szubszomatális üregekbe nőnek. Közvetlen behatolás az epidermális sejtek között csak ritkán figyelhető meg.
A spórák csírázása és a gazdaszervezetbe való behatolás azonos a rezisztens és a fogékony fajtákon. A rezisztens fajtákon a leveleken belül sokkal kevésbé terjednek ki a levélhifák, és sok, de nem minden sejtfalon megfigyelhetőek papillák. A hifák az epidermisz alatt nőnek egyik sztómától a másikig, de nem hatolnak be az epidermisz sejtjei közé. A mezofil kolonizálódik, de nem képződnek haustóriumok. Miután a mezofill sejtek nekrotikussá válnak, az epidermális sejtek összeesnek. A levélben a micéliumfejlődés ritkás, és általában az érkötegek elzárják. Fiatalabb leveleken, ha az érhüvely kevésbé fejlett, a hifák a köteg és az epidermisz között haladnak át. A piknídiumok a szubszomatális üregekben képződnek, többnyire a levél felső felületén (Green és Dickson, 1957).
A spórák élettartamát befolyásoló tényezők a levélfelületen A gabonafélék Stagonospora- és Septoria-kórokozói közül a fertőzési folyamatról csak a következőkre vonatkozóan közöltek végleges információkat S. nodorum, S. tritici, és S. passerinii. Számos más nekrotróf kórokozóhoz hasonlóan a kórokozók egyik csoportja sem vált ki túlérzékenységi reakciót. A Septoria és a Stagonospora kórokozók közötti fertőzési folyamat jelentős különbsége az, hogy a spórák csírázása és behatolása sokkal gyorsabban megy végbe a Septoria és a Stagonospora kórokozók esetében. S. nodorum mint a S. tritici és S. passerinii. Ez jelentős hatással van a betegség járványtanára.
A Septoria-kórokozók elsősorban a sztómákon keresztül hatolnak be a növénybe, míg a S. nodorum közvetlenül és a sztómákon keresztül is behatol. S. nodorum behatol és gyorsan elpusztítja az epidermális sejteket, de a S. tritici és a S. passerinii nem pusztítják el az epidermális sejteket, amíg a levél mezofillumán át nem ágaznak a hifák, és meg nem kezdődik a gyors nekrózis. A gazdaszervezetbe való behatolást követő gombanövekedés szövettani vizsgálatai megfelelnek a gazdaszervezet rezisztenciájára vonatkozó járványtani vizsgálatokból származó adatoknak. A rezisztencia lassítja a gazdaszervezet kolonizációjának sebességét, de nincs számottevő hatása az elváltozás kialakulásának folyamatára.
A gazdaszervezet válaszát irányító mechanizmusok - akár enzimek és toxinok, akár a kórokozók által a fertőzés során felszabaduló egyéb metabolitok - még mindig nem tisztázottak. Az aszkospórák általi fertőzésről kevés információ áll rendelkezésre. A fertőzési folyamat valószínűleg nagyon hasonló a piknídiumspórákéhoz. A Phaeosphaeria nodorum aszkospórái a hőmérséklet széles tartományában csíráznak, és csíracsövük közvetlenül behatol a levélbe. Rapilly és munkatársai (1973) szerint azonban az aszkospórák a piknídióspórákkal ellentétben nem csíráznak szabad vízben.
Septoria spp. fertőzési modell
A Septoria-fertőzés alacsony hőmérsékleten lehetséges, míg a 7 °C alatti hőmérséklet 2 napon belül nem feltétlenül vezet fertőzéshez. A betegség optimális hőmérséklete 16 és 21°C között alakul ki. A fertőzések a magas relatív páratartalom vagy a 14 órás vagy annál hosszabb levélnedvesség esetén lehetségesek. A feltételek teljesítése érdekében úgy döntöttünk, hogy a gyenge, közepes és súlyos fertőzésre vonatkozó modelleket különítjük el. Gyenge fertőzés akkor adható, ha a kórokozó képes megfertőzni a gazdaszövetet. Ez azt jelenti, hogy gyenge fertőzésre akkor kerülhet sor, ha a hőmérséklet a minimumon van, és a levélnedvesség kritikus időtartamú. Mérsékelt fertőzésre olyan körülmények között kerül sor, ahol a legtöbb fertőzési kísérlet elfogadható eredményre vezet, súlyos fertőzésre pedig olyan körülmények között kerül sor, ahol a kórokozónak optimálisak a fertőzési feltételei.
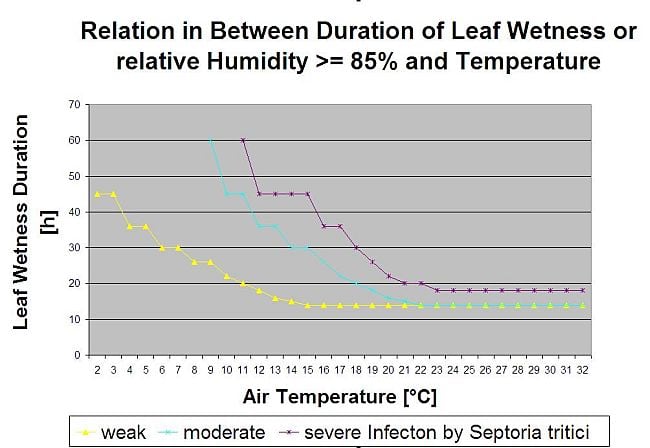

A fertőzés elkezdődik 0,5 mm eső után. Úgy döntöttünk, hogy nem használunk modellt a piknídiumképződésre. A piknídiumképződéshez szükséges feltételnek a 85%-nél magasabb relatív nedvességtartalmú időszakot feltételeztük. A piknídiumok élettartamát 24 órára becsüljük. Minden olyan éghajlaton, ahol a Septoria triticinek esélye van a fertőzésre, szinte minden nap napfelkelte körül 2 órát találunk, amely megfelel ennek a feltételnek.
A fertőzés súlyosságának értékelése
Ahhoz, hogy a Septoria tritici fertőzési nyomást a 10. stádium (az első levél átmérője a koleoptiliumon) és a 32. stádium (a második csomópont legalább 2 cm-rel az első csomópont felett), valamint a 32. és 51. stádium (a fejés kezdete) között fel tudjuk mérni, a fertőzések súlyosságát az éghajlati viszonyok alapján kell értékelnünk. Ezt az értékelést 1-5-ig terjedő skálán végezzük. Az 1-es súlyossági fokozatot akkor kapjuk, ha a gyenge fertőzés feltételei teljesülnek, és 5 mm-nél kevesebb eső esett, ellenkező esetben a megfelelő súlyossági fokozat 2 lesz. A 3-as súlyossági fokozatot akkor kapjuk, ha a közepes fertőzés teljesül, és kevesebb mint 5 mm eső esett. Ha a mérsékelt fertőzés során több mint 5 mm eső esett, vagy kevesebb mint 5 mm eső esett a súlyos fertőzés során, akkor 4-es súlyossági fokozatot kapunk.
Az 5 mm-nél több csapadékkal járó súlyos fertőzés 5-ös súlyossági értéknek felel meg.
Septoria tritici a betegség nyomásának értékelése
Az éghajlat csak az egyik tényező, amely meghatározza a betegségnyomást a szántóföldön. A másik két tényező a termőterület története és a termesztett fajta fogékonysága. Ha a betegség súlyossági értékeit a 10. stádiumtól a 32. stádiumig 4-es értékre tudjuk felhalmozni, akkor az éghajlat által okozott gyenge betegségnyomásra számíthatunk. Ha ez az érték eléri a 6-os értéket, akkor mérsékelt betegségnyomásra számíthatunk, ha pedig eléri a 10-es értéket, akkor magas betegségnyomásra számíthatunk az éghajlat részéről. A fajta fogékonyságának és a termőhely előzményeinek ismeretében ebben a helyzetben gyenge vagy mérsékelt betegségnyomás esetén permetezzünk vagy ne permetezzünk. A 10-es kumulált érték mindenképpen a 32-es stádiumban történő permetezéshez vezethet. A későbbi stádiumban történő permetezésről szóló döntés inkább a tavaszi éghajlattól függ. Ha a 10. stádium óta a súlyossági értékeket 6-os értékre tudjuk felhalmozni, akkor gyenge betegségnyomásra számíthatunk. Ha ez az érték eléri a 10-es értéket, akkor mérsékelt betegségnyomásra számíthatunk, ha pedig ez az érték eléri a 15-ös értéket, akkor az éghajlati helyzetből adódóan magas betegségnyomásra számíthatunk.
Az FieldClimate-ben megmutatjuk a Septoria tritici Súlyosság a három különböző fertőzési súlyossággal együtt egy grafikonon (lásd fent). A csapadék és a hosszú levélnedvesedési időszakok miatt a súlyos fertőzés feltételei a következőkkel biztosítottak S. tritici május 14-én és 16-án már teljes mezőny volt. A súlyossági szintek május 14-én érik el a legmagasabb, 5-ös értéket, ami azt jelenti, hogy most magas a fertőzésveszély.
A Stagonospora nodorum fertőzésbiológiája bizonyos mértékben eltér a S. tritici fertőzésbiológiájától, de ez a különbség nem elég nagy ahhoz, hogy külön modellt alkossunk. Ezért azt javasoljuk, hogy ezt a modellt a gabonafélék Stagnospora és Septoria betegségeinek teljes komplexumára használjuk, beleértve a következőket is S. passerinii. S. tritici és S. passerinii általában hosszabb levélnedvesítési időszakot igényelnek, mint S. nodorum. A magas nyomású területeken S. nodorum A 2-es súlyossági értéket adó gyenge osztályba sorolt fertőzéseket súlyosabban kell kezelni, mint más területeken.
A oldalon. Septoria nodorum a kockázati modell az FieldClimate-ben látható (lásd fent). A június 17-i és a július 7-i (100%) találkozón magas kockázatot állapítottak meg. A fertőzésre fogékony növény stádiumától függően növényvédelmi intézkedéseket kell figyelembe venni, ha a kockázat eléri a 80% értéket (lásd még időjárás-előrejelzés, növényvédelmi védekezés). Ha a kockázat 100% és a fertőzést már megállapították, akkor a növény védelme érdekében szisztémás növényvédelmi intézkedéseket (gyógyító alkalmazást) kell végezni.