Biológia a Plasmopara viticola (Downy Mildew)
P. viticola kötelező parazita. Ami azt jelenti, hogy a növekedéshez zöld, friss szőlőszervekre van szükség. A vegetációmentes időszakban termőtesteket, ún. oospórákat képezve fennmarad. Az oomycéták oospórái nagyon hosszú ideig képesek túlélni a talajban. Ezért olyan helyeken is találkozhatunk lisztharmattal, ahol nem minden évben lehetséges a fertőzés. Tavasszal, amikor a felső talaj elég nedves és meleg, az oospórák úgynevezett makroszpórákat képeznek, amelyek akár 200 zoospórát is kibocsátanak a szabad vízbe. A zoospórák a szél által a vízcseppekben a levelekre és a fürtökre szállnak fel. Két flagellával rendelkeznek, és a levelek vagy a fürtök és a fiatal bogyók alján lévő vízfilmben mozognak, hogy megtalálják a növényi szövetekbe való behatoláshoz szükséges sztómát. Bejutnak a sztómába, és ott csíráznak, ahol kevesebb mint egy órán belül átadják az összes plazmájukat. A mikroszkópos vizsgálatokban a sztómák megtalálása, az encisztáció és a sztómába való csírázás 90 percen belül befejeződött.
P. viticola a sejtek közötti térben növekszik, és az epidermisz és a parenchimasejtekbe behatoló haustoriákból táplálkozik. A hőmérséklet és a relatív páratartalom függvényében elegendő intercelluláris növekedést fejt ki annyi haustoriával, hogy egy szubszisztomatikus testet képezzen, amely kitölti az egész szubszisztomatikus területet, és amely kiemeli az epidermális szövetet a parenchimaszövetből. Ez vezet az olajfolt látható tünetéhez.
Az oomycetesek fény hiányában sporulálnak, ha a relatív páratartalom nagyon magas. A oldalon P. viticola nincs sporuláció, ha a hőmérséklet 12°C alatt van, és a relatív páratartalom 95% alatt van. A sporangiaforák a szubszomatikus vezikulákból képződnek, és a sztómából fognak kijönni. A frissen képződött sporangiumok ragadósak, és csak vízzel lehet eltávolítani őket a sporangiaforákból. A relatív páratartalom csökkenése során a sporangiumok próbaképessé válnak, és széllel is eltávolíthatók.
A sporangiumok akár 20 zoospórát is kibocsátanak a szabad vízbe. Ezeket a zoospórákat a szélnek is szét kell terjesztenie a vízcseppekben, hogy friss levelekre jussanak, vagy a sporangiumokat maga az eső vagy a szél terjesztheti. Az elsődleges és a másodlagos fertőzés fertőzési folyamata azonos.
Mivel a kórokozó hibernációja szempontjából az ivaros szakasznak nagy jelentősége van, feltételezhetjük, hogy a párosodási típusok minden olyan szőlőültetvényben előfordulnak, ahol a szőlő lisztharmat előfordul. A zoospóraképződés a nyár folyamán és kora ősszel az idősebb leveleken zajlik.
Az elsődleges fertőzés részletesen
Az oospórákból kiinduló fertőzéseket elsődleges fertőzéseknek nevezzük. Ez a kifejezés félrevezető, mivel a nyár elején több elsődleges fertőzés is előfordulhat. A járványügyi eseményekben a primer fertőzésnek nincs jelentős szerepe, ha a szőlőültetvényekben elegendő olajfolt van, és a nyári spórák (sporangiumok) fertőzési potenciálja meghaladja az oospórákét.
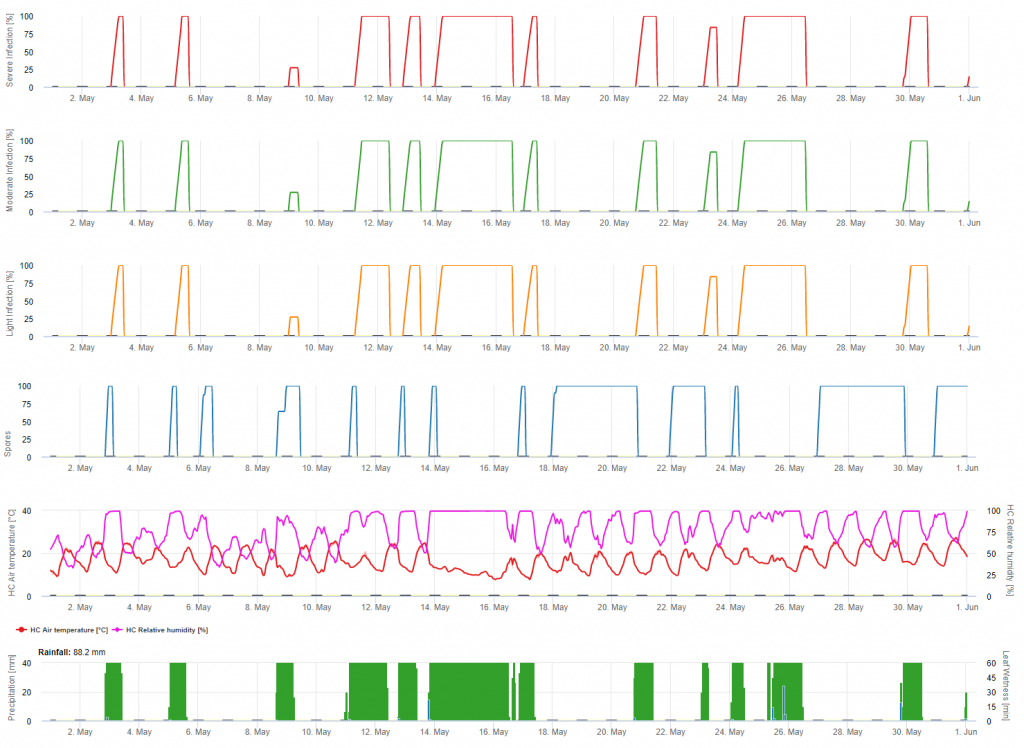
A telelő oospórák úgynevezett makroszpórákká fejlődnek, ha 24 órán keresztül elegendő relatív páratartalom van. A makrosporangiumok szabad vízbe és heves esőbe (például zivatarban) bocsátják ki zoospóráikat. A spórák a szőlő leveleire és/vagy a zöld hajtásokra kerülnek. Elsődleges fertőzések tehát, hosszabb esős időszakok vagy több egymást követő zivatar. Az első eső hatására a lehullott levelek vízzel telítődnek, és a következő napok erős esőzései hatására a makrospórák kibocsátják zoospóráikat, amelyek elérik a szőlő leveleit vagy hajtásait. A sikeres fertőzéshez kellően hosszú levélnedvesítési időre van szükség ahhoz, hogy a zoospórák elérjék a levelek vagy hajtások sztómáit és megfertőzzék azokat (a fenti ábra a szőlő lisztharmat fejlődési ciklusát mutatja (Plasmopara viticola).
A lisztharmat elsődleges fertőzésének modellje először azt ellenőrzi, hogy az időjárás alkalmas-e a makrospórák fejlődésére. Ez mindaddig így van, amíg a levelek nedvesek, vagy az eső utáni relatív páratartalom nem csökken 70% alá. A hőmérséklettől függően a makroszpórangiumok 16-24 órán belül kifejlődhetnek. Ha érett makrosporangiumok állnak rendelkezésre, az grafikusan megjelenik a lisztharmat elsődleges fertőzésének megjelenítésénél. Ha makroszpórangiumok vannak jelen, egy erős esőzés szétterjesztheti zoospóráikat. Egy 5 mm-es folyamatos eső erős esőnek minősül, és a zoospórák elterjednek, az elsődleges fertőzés akkor lehetséges, ha a levélnedvesség elég hosszú ideig tartott.
 Másodlagos fertőzések
Másodlagos fertőzések
 Másodlagos fertőzések
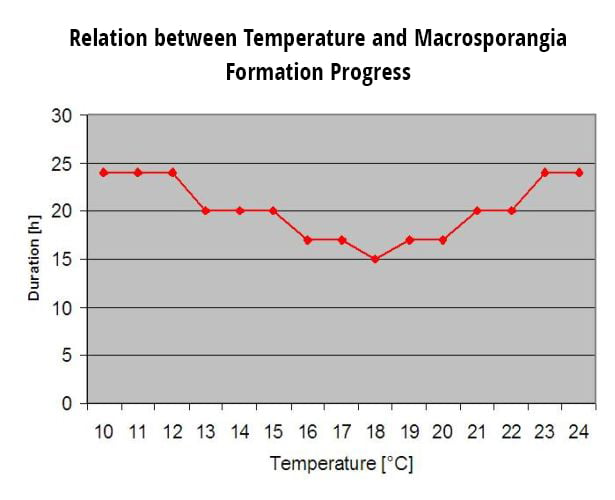
Másodlagos fertőzésekMásodlagos fertőzések a Plasmopara viticola csak akkor lehetséges, ha a szőlőültetvényen vagy annak közelében már érett olajfoltok vannak. Az olajfoltok akkor érettek, ha képesek sporulálni (sporangiumokat termelni). A sporangiumokat csak a következők termelik éjszaka. A napfény gátolja a sporulációt. Sporangiumok termelődnek, ha 12°C-nál melegebb és a relatív páratartalom nagyon magas. A sporangiumtermelés mértéke a hőmérséklettel 24°C-ig növekszik. A sporuláció optimális hőmérséklete az európai szőlőfajtákon (Vitis vinifera) körülbelül 24°C. Ha a hőmérséklet meghaladja a 29°C-ot, akkor a sporulációra nem kerülhet sor. A mi modellünkben azt ellenőrizzük, hogy több mint 95% páratartalom az éjszaka folyamán történik. Ha ez az állapot 50 °C-nál magasabb óránkénti hőmérsékleten keresztül tart, a sporuláció befejeződik, és az új sporangiumok a Plasmopara léteznek a szőlőben. Például az 50°C-os órák 4 órának felelnek meg 13°C-os vagy 3 órának 17°C-os hőmérsékleten.
Sporangia a Plasmopara viticola korlátozott élettartamúak. Minél melegebb van és minél szárazabb a levegő, annál gyorsabban pusztulnak el (fieldclimate.com-ben 0-ra állunk vissza, ha a r.h. 50% alatt van). A következő harmat- vagy levélnedvesedési időszakban biztosan elpusztulnak, ami túl rövid a fertőzéshez. Az idősebb sporangiumok fitneszképessége tehát meglehetősen korlátozott. Modellünk feltételezi, hogy a sporangiumok élettartama egy napra korlátozódik.
Ahhoz, hogy új fertőzéseket okozhasson, a sporangiumoknak el kell terjedniük. A spórák terjesztésének két módja van: Ha közvetlenül a sporangiumok kialakulása után esik az eső, akkor azok az esőcseppekkel terjednek. Ha a szőlőlevelek elég sokáig maradnak nedvesen, akkor a Plasmopara viticola fertőzések zajlanak. Ha a következő reggel eső nélkül és csökkenő páratartalom mellett kezdődik, a kiszáradt sporangiumok leválnak az ágakról. Már enyhe légmozgás is az egészséges levelekre viszi őket. Hacsak nem esik hamarosan eső, elpusztulnak.
Lábszárrothadás Elsődleges fertőzés
A fertőzéseket a makrospórák kifejlődése után (narancssárga, ötödik ábra) három különböző súlyossági osztályban (gyenge: narancssárga, közepes: zöld, súlyos: piros) mutatják be, az eső mennyiségétől és ezáltal a spórák szőlőültetvényben való eloszlásától függően. Ha az 100% fertőzés megállapítható optimális feltételek adottak a növényi szövetekbe való bejutáshoz (növényvédelmi kúraszerűen) és az inkubációs görbe (első grafikon) emelkedni kezd. Ha az inkubáció befejeződik (100%) és nem vették figyelembe a növényvédelmet, a tüneteket (olajfoltok) a szántóföldön kellett volna ellenőrizni.
1. A fertőzés előrehaladási görbéje és a teljes fertőzés után az ebből következő lappangási idő (a fertőzés és a szántóföldön látható tünetek közötti idő).
2. A fertőzés előrehaladási görbéjét akkor határozzák meg, amikor makrosporangiumok vagy sporangiumok vannak jelen és a levelek nedvesek. A levél folyamatos nedvességtartalmával és a hőmérséklettel együtt növekszik. Ha a fertőzés előrehaladási görbéje eléri az 100% értéket, a fertőzés befejezettnek tekinthető. Az adott fertőzéshez tartozó lappangási előrehaladási görbét a fertőzés kezdetével kezdjük el kiszámítani. Ha a fertőzés nem fejeződik be, a lappangási görbe is leáll.
3. Az időjárási viszonyok ugyanazon az oldalon jelennek meg, mint a betegségmodell eredményei. Ezért egyetlen pillantással ellenőrizheti a hőmérsékletet, a relatív páratartalmat, az esőt és a levélnedvességet a fertőzés során.
4. A modell rámutat az elsődleges és másodlagos szőlő lisztharmatfertőzések fertőzési időpontjaira. Ez a legkritikusabb információ. Azok a fertőzések, amelyekre még nem terjedtek ki sem a megelőző, sem a gyógyító gombaölő szerek, katasztrofálisak lehetnek a termés/minőség szempontjából. A fertőzés súlyosságára vonatkozó további információk (gyenge/közepes/súlyos) hasznosak lehetnek a növényvédelmi stratégia eldöntésében, a szőlőültetvény előzményeinek (az inokulum mennyisége), a fajtának és az előző évek tapasztalatainak függvényében.
Növényvédelmi stratégiák:
- A lisztharmat elleni védekezésre széles körben alkalmaznak profilaktikus és szisztémás vagy gyógyító fungicideket. A profilaktikus vegyszereket a fertőzés előtt, de a lehető legközelebb a fertőzéshez, a gazdaszervezet legnagyobb fogékonyságának időszakában, a 10 cm-es hajtáshossz és a borsó nagyságú bogyók között alkalmazzák.
- A megelőző vegyszereket úgy is lehet használni, hogy a lehető legközelebb permeteznek az előre jelzett, 2 mm-nél több esőt hozó időjárási esemény előtt (az elsődleges kör esetében).
- A gyógyító gombaölő szereket a lehető leghamarabb a fertőzés (100% fertőzés) után és az olajfoltok megjelenése előtt kell kijuttatni. Figyeljen a gombaölőszer-rezisztencia kialakulásának megelőzésére.
- A szőlőt a lisztharmat (olajfoltok) jelenlétének ellenőrzését rendszeresen és legalább 200 tőkét figyelembe véve kell elvégezni. Kockázatosnak tekinthető, ha 50 tőkénként több mint két olajfoltot észlelünk.
A modell MUELLER-THURGAU, ARENS, MUELLER és SLEUMER, BLAESER, HERZOG, GEHMANN és sok más, a szőlő lisztharmat és lisztharmat járványtanával foglalkozó kutatóhálózathoz a múlt század tizenkilencedik század eleje óta kapcsolódó munkatársak publikációin alapul.
Biológia a Uncinula necator
A szőlőt lisztharmat a gomba okozza. Uncinula necator. Kötelező parazita, ami azt jelenti, hogy a táplálkozáshoz zöld gazdaszövetre van szüksége. U. necator a tizenkilencedik században került Európába, és a század végéig minden bortermelő országban elterjedt. Nagyon erős hatással van a terméshozamra, és a szőlő lombtalanodásához vezethet, ha nincs kémiai védekezés. Minden európai szőlőfajta fogékony erre a kórokozóra. A kórokozóval szembeni rezisztencia nemesítése még eléggé új, és az eredmények elérése több éves munkát igényel.
U. necator a legtöbb szőlőtermő területen túl kell lépnie a vegetációmentes időszakon. Ehhez a kórokozó kétféle mechanizmust használ: a) a termőtestekben, az úgynevezett kleisztotéciumokban marad életben, vagy b) a telet is túléli a nyugvó rügyekbe zárva, ahol egész télen élő szöveteket talál. Az, hogy egy szőlőültetvényen belül csak az egyik vagy mindkét téli álmot alvási mód jelen van-e, az adott szőlőültetvény betegségtörténetén alapul. A rügyek csak addig fertőződhetnek, amíg nem zártak és nem veszi őket körül az öregedő szövetek gyűrűje. Ez csak a kialakulás után néhány héttel van így. Szokásos szőlőtermesztési helyzetben a szőlőt 14 rügynél kevesebb rügyet tartalmazó ágakra vagy hajtásokra metszik vissza. Ezek a megmaradt rügyek csak a virágzásig fertőződhetnek. A másik oldalon a BBCH 15-19. stádiumú zászlós hajtásokat találunk. A zászlós hajtások részben vagy teljesen lisztharmattal borított hajtások, amelyek látens fertőzött rügyekből képződtek. Ezek a zászlós hajtások korai és nagyon erős inokulumforrást jelentenek. A szőlőültetvényen belül többnyire nem terjednek el. Ez azt eredményezi, hogy a szőlőültetvényeken belül a betegség korai és magas előfordulási gyakoriságú foltok, a szezon elején pedig alacsony vagy közepes mértékű betegség előfordulási gyakoriságú foltok alakulnak ki. A zászlós hajtások vegetatív történetük miatt csak egy párosodási típust képviselnek. Ez olyan szőlőültetvényekhez vezethet, ahol egyetlen párosodási típus dominanciája miatt csak kevés vagy egyáltalán nem találunk cleistotheciát.
Ahol a kleisztotécium az egyetlen inokulum, ott a kleisztotécium téli álmodásának folyamatos előzményeivel találkozunk, amelyek csak azokban az években válthatnak zászlós hajtásokra, amikor a betegség nagyon korán kialakul. A kleisztotéciumok képződése jól látható az ivaros kombinált szövetek sárga elszíneződése miatt. A nem permetezett szőlőültetvényekben nyár közepétől megtalálható. A permetezett szőlőültetvényekben hosszabb időre van szüksége a betegség előfordulásának kialakulásához, lehetővé téve a párosodó típusok találkozását. Az elterjedése Uncinula necator a nyár folyamán elsősorban a hőmérséklet befolyásolja.
A lisztharmat számára az optimális hőmérséklet 21°C és 32°C között van. Minél több a napi órák száma ebben a hőmérsékleti tartományban, annál nagyobb a lisztharmat kockázata. A konídiumképződés, a fertőzés és a lappangási sebesség hasonló hőmérséklettartományt igényel. Annak a ténynek köszönhetően, hogy U. necator nem igényel szabad vizet a fertőzéshez, és nincs magas relatív páratartalom a konídiumok kialakulásához, nincs hasznosítható hatása a nedvesség paraméterének közvetlenül a kórokozóra.
A szabad nedvesség jelenléte kölcsönhatásba lép a U. necator közös hiperparazitáján keresztül Ampelomyces quisqualis. Ez a gombás hiperparazita növekedés a lisztharmatgomba hifáinak belsejében. A konídiumokat és kleisztotéciumokat saját piknídiumokká alakítja át. Szabad nedvességre vagy nagyon magas relatív páratartalomra van szüksége a fertőzéshez, és vékony, gyorsan növekvő hifákkal növekszik a leveleken, hogy megtalálja a lisztharmat okozta elváltozásokat. Erősen fertőzött U. necator Az elváltozások nagyon sötétbarnára színeződnek. Eső idején a hiperparazita piknídiumaiból származó fehér spórakiválás borítja őket. Ampelomyces quisqualis a lisztharmat elleni védekezéshez kereskedelmi termékként is kapható.
Gyakorlati munka
A fertőzési folyamatra vonatkozóan különböző modelleket vezettünk be FieldClimate.
a) a gomba főként micélium formájában telel át. Tavasszal a micélium növekedésnek indul, és konídiumokat képez konídiumokkal (tehát nincs elsődleges fertőzés).
b) ha a gomba ősszel fejlődik ki a bogyókon, akkor az ivaros forma jelen lehet (kleisztotéciumok képződése), és így tavasszal az aszkospórák kibocsátása vezethet elsődleges fertőzéshez (lásd: modell: POROSZMÁLYI MÉLYEK/ASKOSPÓRÁK INFEKCIÓJA).
Kaliforniai kockázati modell (Thomas Gubler)
A UC Davis lisztharmat kockázati értékelési modellje (Gubler-Thomas-modell) a szőlő lisztharmatával kapcsolatos laboratóriumi és terepi járványtani vizsgálatokon alapul Kaliforniában.
Kockázatértékelési index:
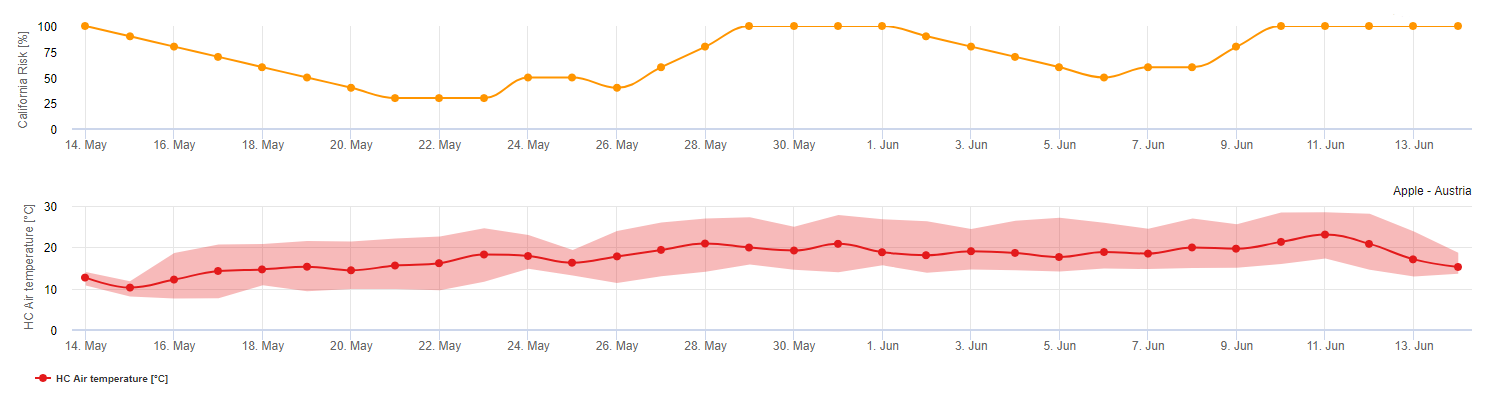
Az aszkospórák felszabadulását és csírázását követően (ehhez a modellhez használja az ASKOSZPÓRÁK BEFERTŐZÉSÉT) , a lisztharmat későbbi fejlődését és szaporodását elsősorban a környezeti hőmérséklet befolyásolja. A UC Davis modellje értékeli a környezeti hőmérsékletet, és egy 0-100 pontos index segítségével értékeli a lisztharmat kialakulásának kockázatát.
A kockázatértékelési index indításához a rügyfakadást és a betegség kezdetét követően három egymást követő napon legalább hat egymást követő órán át 21 és 30 °C közötti hőmérsékletnek kell lennie ahhoz, hogy a lisztharmatjárvány meginduljon. Az index kiszámítása során minden olyan nap után, amikor 6 vagy több egymást követő órán át 21 és 32 °C között van a hőmérséklet, az index 20 ponttal növekszik. A modell ezután 0 és 100 indexpont között ingadozik a napi hőmérsékletek időtartama alapján. Az index 20 pontot nyer minden olyan napért, amely megfelel a 21 és 32 °C közötti hőmérséklet hat egymást követő órára vonatkozó követelményének, és 10 pontot veszít minden olyan napért, amely nem felel meg a hat órás követelménynek, vagy ha a nap folyamán bármikor 32 °C fölé vagy 21 °C alá emelkedik a hőmérséklet.
Az index értelmezése:
Az alacsony, 0-30-as indexértékek azt jelzik, hogy a kórokozó nem szaporodik. A termelők fontolóra vehetik a gombaölő szerek alkalmazásának elhalasztását az alacsony indexértékekkel jellemezhető hosszabb időszakokban. A 40-50-es index közepesnek tekinthető, és a lisztharmat körülbelül 15 napos szaporodási időt jelent. A 60-100-as indexértékek azt jelzik, hogy a kórokozó gyorsan (5 naponta) szaporodik, és nagy a járványos betegség kialakulásának kockázata.
Pessl Instruments kockázati modell
A szétszóródás Uncinula necator a nyár folyamán elsősorban az alábbiak befolyásolják hőmérséklet. Az antagonista a U. necator a címen. Ampelomyces quisqualis elsősorban a szabad nedvesség kedvez. A lisztharmat számára az optimális hőmérséklet 21°C és 32°C között van. Minél több órát tölt naponta a hőmérséklet ebben a tartományban, annál nagyobb a lisztharmat kockázata (lásd a Gubler-modellt). Az olyan napok, amikor a hőmérséklet több mint 6 órán át ebben a tartományban van, körülbelül 20 ponttal növelik a kockázatot.. Azok a napok, amikor a hőmérséklet nem ebben az optimális tartományban van, körülbelül 10 ponttal csökkentik a kockázatot. Ezek a napok, amikor a hőmérséklet egész nap 21 °C alatt van, vagy amikor a hőmérséklet 6 órán keresztül meghaladja a 32 °C-ot. Ugyanakkor a következő napok hosszú levél nedvesség időszakok az antagonista gomba megtelepedéséhez vezetnek. A. quisqualis. Ezek a következők csökkenéséhez vezetnek U. necator.
Ehhez a modellhez a következő szabályokat ajánljuk: Ha a lisztharmat veszélye 20 pontnál kisebb, a permetezési időköz meghosszabbítható. 20 és 60 pont között a szokásos permetezési időköz érvényes. Ha a kockázat meghaladja a 60 pontot, akkor a permetezési intervallumot rövidíteni kell.
A kockázat elindul: 3 olyan nappal, amikor legalább 6 órát töltött: 21°C <= Hőmérséklet 60 pont
A kockázat növekszik: Minden olyan nap, amikor legalább 6 óra: Hőmérséklet +20 pont
A kockázat csökken: Minden olyan napon, amikor a hőmérséklet nem éri el a 21°C-ot, vagy minden olyan napon, amikor 6 órán át: 32°C -10 pont
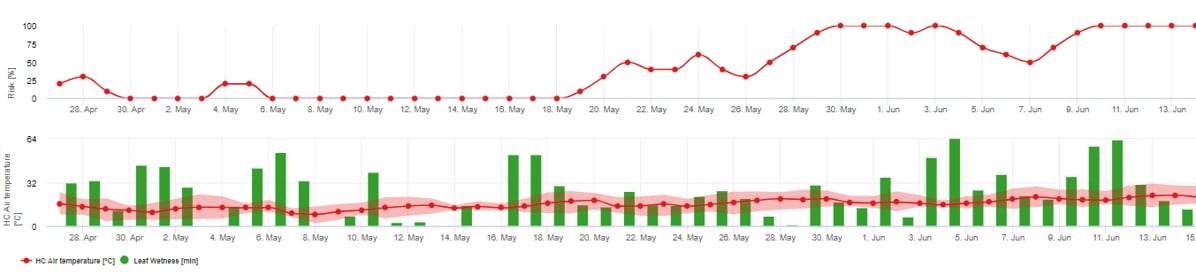
Harmadik kockázati modell: Az aszkospórák elsődleges beoltása
Ezt a modellt akkor javasoljuk használni, ha az aszkospórák a hideg téli időszakban képződnek (tehát nem a micéliummal fertőznek ("zászlós hajtások").
Az aszkospórafertőzés előrejelzi az aszkospórák felszabadulását és a kezdeti fertőzéseket, amelyek a hosszabb levélnedves időszakok alatti átlaghőmérsékleteken alapulnak.
Az fieldclimate.com-ben történő számításhoz körülbelül 2.5mm csapadék szükséges az aszkospórák felszabadításához, majd a legalább 8-12 órányi levélnedvesség az aszkospórák felszabadulása után a következő hőmérsékleti értékek között 10 -15°C fertőzésre. Ha a fertőzés megtörtént, a modell átvált a betegség kockázatértékelési fázisára (Gubler/Risk modell használata), és teljes mértékben a hőmérsékletnek a kórokozó szaporodási sebességére gyakorolt hatásán alapul.
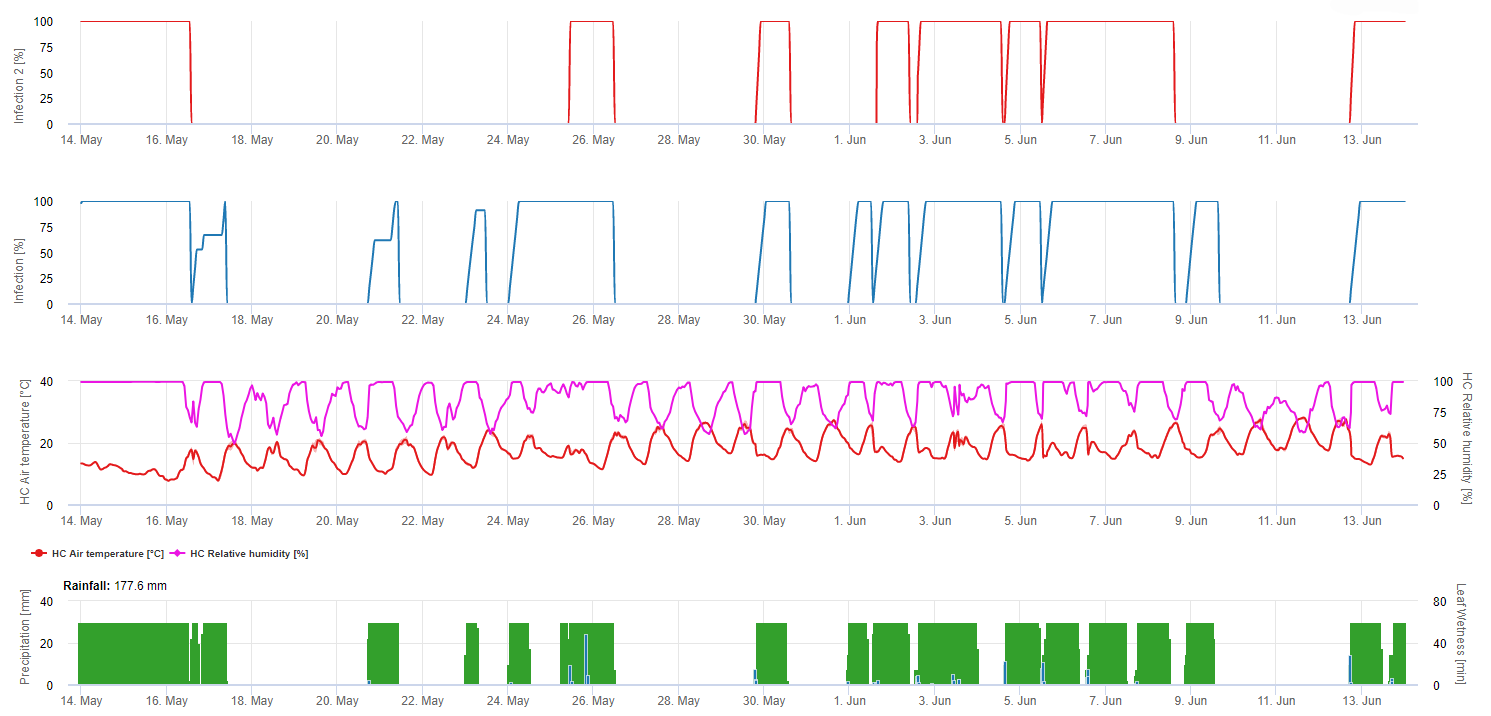
Gyakorlati használat
Ebben a modellben a fertőzést a következő módon jelenítjük meg Uncinula necator egy 0 és 100% közötti sorban.
Két különböző modellt mutatunk be FieldClimate:
1. A David Gadoury és Doug Gubler által javasolt modelleket követő lehetséges aszkospórafertőzések az aszkospórafertőzés grafikonján láthatóak. Ha a fertőzési modell fejlődési görbéje eléri az 100% értéket, akkor fennáll az aszkospórafertőzés lehetősége. Az esemény után 3-5 nappal a lisztharmat első tünetei már láthatóak lehetnek a régi kordonhoz közeli leveleken.
2. A fenti grafikonon a kaliforniai modell és a mérsékelt éghajlati övezetekre vonatkozó modell alapján a lisztharmat kockázata látható. A kockázatot jelentő időszakokat gombaölő szerek alkalmazásával kell lefedni. A 60 pontnál magasabb kockázatú időszakokban ajánlott gyakrabban permetezni, különösen a virágzás körül és röviddel a virágzás után.
A szőlőfeketerothadás számos szőlőtermelőt érint az Egyesült Államokban és a világ legtöbb más meleg és száraz szőlőtermő vidékén. A betegség kezeléséhez fontos, hogy megértsük a betegség életciklusát és a környezeti feltételeket. A fertőzés bekövetkezése után különböző módszerek állnak rendelkezésre a betegség elleni védekezésre.
Biológia
A szőlőfeketerothadás kórokozója a szőlőtőkék számos részén telel át, és a talajon is képes telelni, különösen a múmiákban. Ezenkívül a kórokozó "legalább 2 évig képes telelni a fertőzött hajtások sérüléseiben, amelyek vesszőként vagy hajtásként maradnak meg".
Egyszer az esőzések felszabadíthatják a telelő spórákat (aszkospórákat), amelyek a talajon lévő múmiákban képződnek, és a szél által szétszóródhatnak. A talajon lévő múmiák némelyikében jelentős aszkospórakibocsátás kezdődhet, amely körülbelül 2-3 héttel a rügyfakadás után kezdődik, és a virágzás kezdete után 1-2 héttel érik be. Egy másik spóratípus (konídiumok) is képződhet a vesszős elváltozásokban vagy a "rácson belül maradt múmiákban, és ezek rövid távolságokra (centiméterekről méterre) szóródnak szét a fröccsenő esőcseppek által". A fertőzés akkor következik be, ha bármelyik spóratípus a zöld szőlőszövetre kerül, és a szövet "megfelelő ideig nedves marad, ami a hőmérséklettől függ". A forrástól függ, hogy ezek a telelő spórák mennyi ideig képesek fertőzést okozni. Ha a fertőzésnek nagy a forrása, a fertőzés korán megindul. Nedvesség jelenlétében ezek az aszkospórák lassan csíráznak, ami 36-48 órát vesz igénybe, de végül behatolnak a fiatal levelekbe és a gyümölcsök szárába (pedicel). A fertőzések 8-25 nap után válnak láthatóvá. Ha az időjárás nedves, az aszkospórák egész tavasszal és nyáron termelnek és szabadulnak fel, folyamatos primer fertőzést biztosítva. (Wilcox, Wayne F. "Fekete rothadás Guignardia bidwellii." 102GFSG-D4 számú betegségazonosító lap. 2003. Cornell. Oct. 2010. október 24.).
A feketerothadás gombának tehát meleg időjárásra van szüksége az optimális növekedéshez; a hűvös időjárás lelassítja a növekedését. A fertőzéshez 2-3 napos eső, szitálás vagy köd is szükséges." (Ries, Stephen M. "IPM : Reports on Plant Diseases : Black Rot of Grape (A szőlő fekete rothadása)". Integrált növényvédelem az Illinois-i Egyetemen. Dec. 1999. 2010. okt. 24.). Az esőcseppek ezeket a spórákat a spórák különböző növényi részekre, különösen a fogékony fiatal levelekre történő átvitelével juttatják el. Ha víz van jelen, a konídiumok 10-15 óra alatt kicsíráznak és behatolnak a fiatal szövetekbe. A fekete rothadás új fertőzések késő tavasszal és nyáron is folytatódnak a meleg, csapadékos időjárás hosszabb időszaka alatt. A konídiumok a kialakulásuk után több hónappal is képesek kicsírázni és fertőzést okozni. Augusztus folyamán a piknídiumok átváltoznak egy telelő stádiumba, amelyből pszeudotéciumok fejlődnek, amelyekben a tavaszi spórák (aszkospórák) termelődnek. A kutatások kimutatták, hogy az aszkospórák a tavaszi primer fertőzések fontos forrását jelentik. Tavasszal, nedves időjárás esetén a "püknídiumok a fertőzött szöveteken vizet szívnak fel, és a konídiumok kipréselődnek. A konídiumokat az eső véletlenszerűen szétfröcsköli, és 60-90 fokos hőmérsékleten kevesebb mint 12 óra alatt bármely fiatal szövetet megfertőzhetnek." (Ellis, Michael A. "Fact sheet Agricultural and Natural Resources: Grape Black Rot." Növénykórtani Tanszék. Az Ohiói Állami Egyetem Állami Hagyományőrző Központja. 2008).
Gyakorlati használat
Modellünk alapjául a Spotts által publikált szakirodalom szolgál (Spotts. R. A. 1977. A levélnedvesség időtartamának és a hőmérsékletnek a hatása a levélfoltosság fertőzőképességére Guignardia). A Daniel Molitorral folytatott megbeszélések révén az eredeti Spotts-modellt módosítottuk egy olyan modell felé, amely az almabetegség-modellekhez hasonlóan könnyű, közepes és súlyos fertőzéseket különböztet meg. (Daniel Molitor: Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Dissertation Justus-Liebig-Universität Gießen, 2009).
A "Spotts-kritériumoknak" megfelelő fertőzés könnyű fertőzésnek minősül. A 150% által a "Spotts-kritériumoknak" megfelelő fertőzés közepes, a 200% által a "Spotts-kritériumoknak" megfelelő fertőzés pedig súlyosnak minősül.
A modell a fertőzés időpontját mutatja a könnyű, közepes és súlyos fertőzések esetében. Az 100% fertőzés teljesülésekor a spórakibocsátás, a csírázás és a növényi szövetek fertőzésének feltételei adottak. A fertőzés súlyossága a gomba optimális hőmérsékletének és nedvességtartalmának időszakától függ. A fertőzési nyomás függvényében a fertőzéseket megelőzően kell lefedni, vagy röviddel a fertőzés után gyógyító permetezést kell alkalmazni. A Mosel vagy a legtöbb osztrák bortermelő terület mérsékelt, félszáraz éghajlatán nem ajánlott a permetezés a könnyű fertőzés megállapítása után.
Lobesia botrana (Totricidae, Olethreutinae) először Ausztriából írták le, de Dél-Olaszországból származik, és egész Európába, Észak- és Nyugat-Afrikába, a Közel-Keletre és Kelet-Oroszországba vándorolt. Nemrégiben Japánba is behurcolták, és 2008-ban jelentették először Chiléből.
Házigazdák
Szőlő (Vitis vinifera) és a sarkantyús babér (Daphne gnidium) a kedvelt gazdanövények, de a szeder (Rubus fruticosus), egres (Ribes sp.), fekete és piros ribizli (Ribes nigurm), olajbogyó (Olea europaea), cseresznye (Prunus avium), metszés (Prunus domestica), datolyaszilva (Diospyrus kakis), kiwi (Actinidia chinensis), gránátalma (Punica granatum), szegfű (Dianthus spp.), és számos más vadon élő gazdaszervezet.
Kár
Májusban és júniusban az első generációs lárvák a virágfürtökön pókhálóznak és táplálkoznak. A második generációs lárvák (július-augusztus) a zöld bogyókon táplálkoznak. A fiatal lárvák behatolnak a bogyóba és kivájják azt, meghagyva a héjat és a magokat. A harmadik generációs lárvák (augusztus-szeptember) a legnagyobb kárt a bogyók belsejében és a fürtökben való táplálkozással okozzák, amelyek ürülékkel szennyeződnek. A bogyók táplálkozással történő károsítása továbbá kedvez a Botrytis és más másodlagos gombák, például a következők fertőzésének Aspergillus, Alternaria, Rhizopus, Cladosporium, és Penicillium.
Biológia és életciklus
A kifejlett lepke körülbelül 6-8 mm hosszú, szárnyfesztávolsága 11-13 mm. A nőstény valamivel nagyobb. Mind a hímek, mind a nőstények szárnyai hasonló mozaikos mintázatúak. Az első szárnypár (elülső szárny) barnáskrémszínű, szürkéskék, barna és fekete foltokkal tarkítva. A második szárnypár szürke, rojtos szegéllyel. A szárnyak nyugalmi állapotban harang alakban a has fölött helyezkednek el. A nőstények a tojásaikat külön rakják le. A tojások elliptikusak és laposak, körülbelül 0,6-0,8 mm átmérőjűek. Ezek a lencse alakú tojások láthatóak és irizáló krémfehér színűek, az embrió fejlődésével sárgává, majd később feketévé válnak, amikor a fejlődő lárva feje kialakul. Öt lárvastádium létezik. Az első stádium körülbelül 1 mm, az utolsó stádium pedig körülbelül 12-15 mm méretű. A lárvák fekete fejjel rendelkeznek, majd sárgásbarnára színeződnek, és a testük a táplálékuk (béltartalmuk) színét veszi fel. Az ötödik utolsó lárvastádium szürkés szilikongubót fon a bábozódáshoz. A báb körülbelül 4-9 mm hosszú.
A mi éghajlati övezetünkben az európai szőlőbogyó-moly évente két-három nemzedéket hoz létre. Spanyolország, Görögország és Egyiptom melegebb régióiból négy generációról számoltak be. A bábállapot a szilikongubókban telel át (diapauza) a kéreg alatt vagy a talaj repedéseiben, illetve rejtett, védett helyeken. Amikor a levegő hőmérséklete 10 °C fölé emelkedik, körülbelül 10-12 napig A felnőttek kikelnek. Először a hímek találhatók meg, és egy hét múlva kelnek ki a nőstények. Az első hímek repülése tehát már rügyfakadáskor megkezdődhet, és a repülés körülbelül 4-5 hétig tart. A repülési tevékenység akkor zajlik, amikor a nap felkel és a hőmérséklet körülbelül 12°C. A repülés során a rovarok párosodnak, és a nőstények többsége csak egyszer párosodik. A párosodás után röviddel (körülbelül egy-két nappal) megkezdődik a tojásrakás. A tojásokat egyesével rakják le a virágokra vagy azok közelébe. Egy nőstény körülbelül 100 tojást rak le, és a kifejlett életkor az éghajlati viszonyoktól függően 1-3 hétig tart.
Tavasszal a tojások kikelése 10 napot vagy annál is többet vesz igénybe, míg nyáron a tojások a hőmérsékleti viszonyoktól függően 3-4 napon belül kikelnek. Az első generációs lárvák a virágokon hálózzák be magukat és táplálkoznak, bejuthatnak a virágszárba és a fürt kiszáradását okozhatják. A lárvák fejlődéséhez az időjárási viszonyoktól függően 20-30 napra van szükség. Ezután a kéreg alatt vagy a talajban, illetve a virágon bábozódnak be, és a kifejlett egyedek 6-14 nappal később kelnek ki újra.
A másod- és harmadrepülő nőstény lepkék a tojásokat egyenként közvetlenül az árnyékos bogyókra rakják. Röviddel a lárva kikelése után behatol egy bogyóba, és táplálkozás közben kilyukasztja azt. Egyetlen fürtöt több lárva is megfertőzhet. A pókhálósodás, a pelyva és a gombafertőzés a fürt nagymértékű fertőzöttségét eredményezheti.
A fejlődés alsó küszöbértéke körülbelül 10 °C, a felső fejlődési küszöbérték pedig 30 °C. Az optimális fejlődési feltételek 26-29 °C és 40 és 70% közötti páratartalom között vannak. Ha a nappalok hossza csökken, és egyre hidegebb van, a diapauza szakasza beindul. A lárvastádiumok elpusztulnak, ha a hőmérséklet 8°C alá csökken, de a diapauzában lévő bábok még a hideg észak-európai teleket is kibírják.
Az első generáció rövidebb, mint a nyári generációk. A tojások körülbelül 66 Celsius-fokos nap alatt kelnek ki. A virágfürtökön táplálkozó lárvák a jelentések szerint gyorsabban fejlődnek, mint a szezon későbbi szakaszában a szőlőbogyókon táplálkozó lárvák, és ez befolyásolja a generációs időt. A nem diapausáló bábok fejlődéséhez körülbelül 130 DDC-re van szükség. A kifejlett nőstények körülbelül 61 DDC után rakhatnak petéket.
Az egy generációra vonatkozó DD becslések jelentősen eltérnek a szakirodalomban, az első generációban 427 DDC-től 577 DDC-ig, a későbbi generációkban 482 DDC-től 577 DDC-ig.
Monitoring és irányítás
A hímeket feromonok vonzzák, amelyekkel a hímek repülését figyelik. Rügyfakadás előtt helyezzenek ki csapdákat L. botrana csalétek magasan a lombkoronában (iScout feromon). Helyezzen el legalább egy csapdát 30 hektáronként vagy kisebb szőlőültetvényblokkonként. A csalétket a gyártó ajánlásainak megfelelően cserélje. Hetente ellenőrizze a csapdákat, jegyezze fel a befogott molyok számát és távolítsa el a csapdába esett molyokat a ragadós csapda aljáról. A heti fogásokat ábrázoljuk, hogy meghatározzuk a hímek repülésének kezdetét és csúcspontját az egyes generációkban. Folytassa a csapdákkal történő megfigyelést a harmadik repülés csúcspontjáig.
A rovarölő szerek kijuttatását a lárvák megjelenésének időpontjára kell időzíteni, ezért a tojásrakás nyomon követése és a peték kikelésének meghatározása alapvető fontosságú e kártevő elleni védekezésben. Az első generáció esetében a tojásrakást a csúcspontjától a repülés végéig kell nyomon követni. Keressük a tojásokat 100 fürt kocsányán, szőlőnként egy fürtöt kiválasztva. Jegyezze fel a talált tojások többségének stádiumát. A nemrégiben lerakott tojások fehér színűek, majd sárgává, később feketévé válnak, amikor a lárvák már a keléshez közelednek. Az áttetsző tojáshéj azt jelzi, hogy a lárva már kifejlődött. A tojás kikelése után keresse a virágrészek pókhálósodását. Nyissa fel a pókhálót, és keressen táplálkozási sérüléseket és lárvákat.
A második és harmadik generációs peték bogyókon történő megfigyelését egy héttel azután kell megkezdeni, hogy az adott repülés első lepkéi a csapdákba kerültek. Folytassa a tojások ellenőrzését hetente, amíg egy héttel a csúcsrepülést követően el nem hagyja. Vizsgáljon meg 100 fürtöt, tőkénként egyet kiválasztva. Folytassa a fürtök ellenőrzését a táplálkozási károk (lyukak vagy üreges bogyók), a szövedékek és a lárvák jelenléte szempontjából.
Azokban az országokban, ahol L. botrana megállapításra kerül, a védekezési intézkedések a második generációra irányulnak. Ennek oka részben az első generáció elhúzódó megjelenése, valamint a szomszédos, kezeletlen szőlőültetvényekről történő esetleges újrafertőződés. Az első generáció kezelése azonban akkor ajánlott, ha a populációk magasak, vagy ha a kezeléseket az egész területre kiterjedően végzik. A fürtzáródás után a rovarölő szerek kevésbé hatékonyak. Számos csökkentett kockázatú rovarölő szert törzskönyveztek a szőlőben a tortricidák lárvái elleni védekezésre. Ezek közé tartoznak a rovarnövekedés-szabályozók, a spinosinok és a Bacillus thuringiensis.
A párzási zavarokat már több éve tanulmányozzák Európában. Akkor bizonyult a leghatékonyabbnak, ha a szőlőmoly populáció alacsony, és ha nagy, több mint 10 hektáros vagy az egész területre kiterjedő területeken alkalmazzák.
Az európai szakirodalom számos ragadozóról és parazitoidról számol be. A parazitoidok között 4 tachinida légyfaj és közel 100 parazita darázsfaj található a ichneumonidák, braconidák, pteromalidák és chalicidoidák családjából. A beszámolók szerint a legnagyobb hatást azok a paraziták okozzák, amelyek az áttelelő bábokat támadják meg. Spanyolországban ezek közé tartoznak a pteromalidák. Dibrachys affinis és D. cavus, amelyek a jelentések szerint akár 70% bábpusztulást is okozhatnak, míg Olaszországban az ichneumonidák Dicaelotus inflexus és Campoplex kapitátor a legfontosabbak.
Irodalom:
- Briere JF, Pracros P. 1998. Hőmérsékletfüggő növekedési modellek összehasonlítása a Lobesia botrana (Lepidoptera: Tortricidae) fejlődésével. Environ. Entomol. 27: 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. en las comarcas vitícolas valencianas. Bol. Serv. Plagas 7: 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). In Los parasitos de la vid, estrategias de proteccion razonada. Madrid, Spanyolország. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33: 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. A Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) szexferomoncsapdákkal történő fogása és a foknapok halmozódása közötti kapcsolat vizsgálata Sherry szőlőültetvényekben (Spanyolország délnyugati része). J. Appl. Ent. 125: 9-14.
- Gabel B, Mocko V. 1986. A Lobesia botrana Den európai szőlőmoly funkcionális szimulációja. Et Schiff. (Lep., Torticidae) populációjának fejlődését. J. Appl. Ent. 101: 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. A Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) repülési aktivitásának előrejelzése Délnyugat-Spanyolországban. J. Appl. Entomol. 133: 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. A Sparganothis pilleriana Schiff. és a Lobesia botrana (Den. & Schiff). elleni védekezés német szőlőültetvényekben szexferomon-közvetítésű párzási zavarással. Bull. IOBC/WPRS 25: 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera: Tortricidae): influence de l'infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thése N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. A Daphne gnidium, a Lobesia botrana európai szőlőmoly lehetséges őshonos gazdanövénye, serkenti a Lobesia botrana petézését. Fontos-e a gazdanövényváltás? Chemoecol. 16: 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. A szőlőmoly vonzása a gazdanövény és nem gazdanövény részeihez szélcsatornában: a növény fenológiájának, nemének és párzási állapotának hatása. Entomol. Exp. Appl. 122: 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Nappali-fokozatú modellek a Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) helyi populációinak nemzési idejének és repülési aktivitásának előrejelzésére Görögországban. J. Appl. Ent. 125: 515-518.
- Moreau J, Benrey B, Thiéry. 2006. A szőlőfajta befolyásolja a Lobesia botrana európai szőlőmoly (Lepidoptera: Tortricidae) lárvális teljesítményét és a nőstények szaporodási teljesítményét. Bull. Entomol. Res. 96: 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. A metoxifenozid hatása a Lobesia botrana (Den & Schiff) (Lepidoptera: Torticidae) petéire, lárva- és kifejlett stádiumaira.
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. In Les ravageurs de la vigne. Féret, Bordeaux. pp. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. A Lobesia botrana lárvák szóródása és túlélése, valamint a szőlővirágzatban lévő lárvák sűrűsége közötti kapcsolat. Entomol. Exp. Appl. 84: 109-114.
- Xuéreb A, Thiéry D. 2006. Változik-e a Lobesia botrana (Lepidoptera: Torticidae) természetes lárvaparazitizmusa az évek, a generáció, a gazdaszervezet sűrűsége és a szőlő kultúrrivar szerint? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). In Lepidotteri dei fruttiferi e della vite. Milánó, Olaszország. Pp. 85-88.
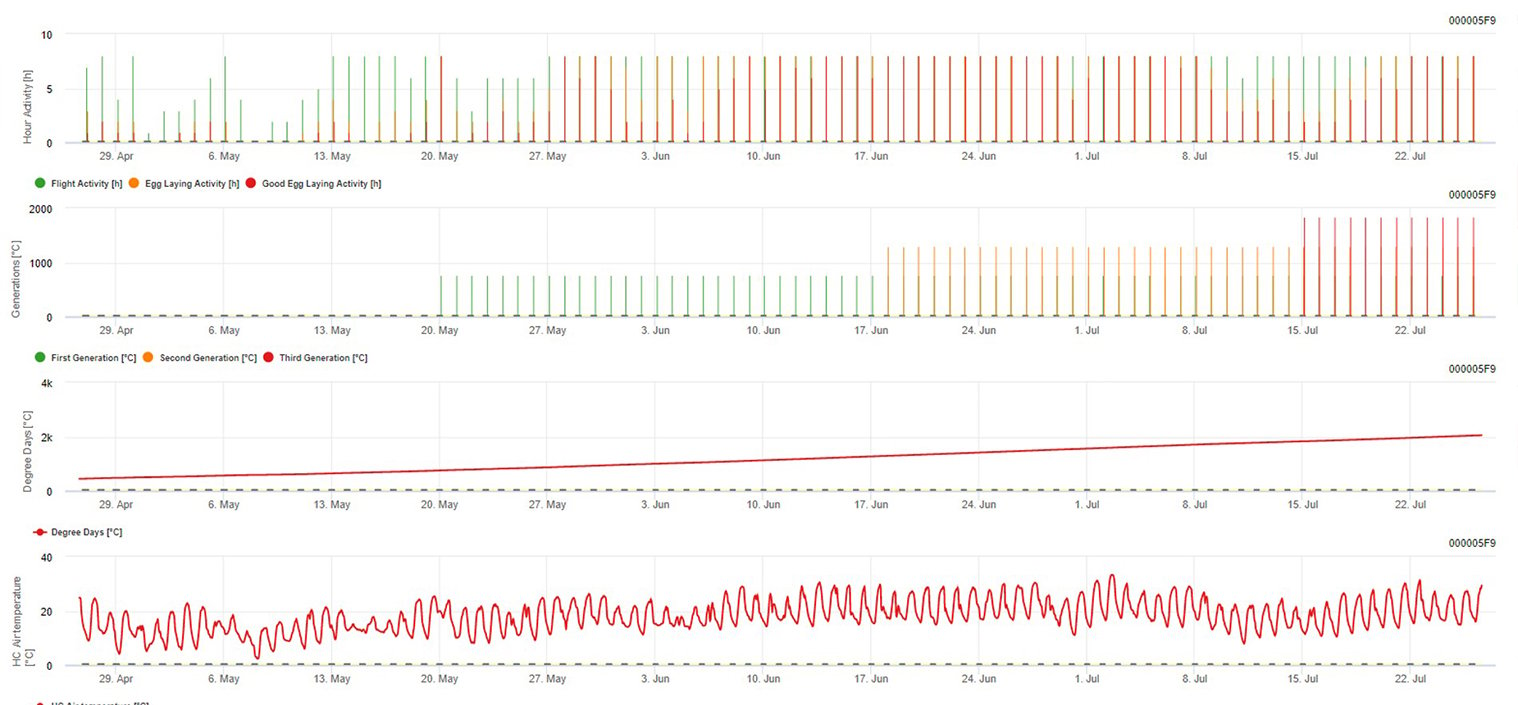
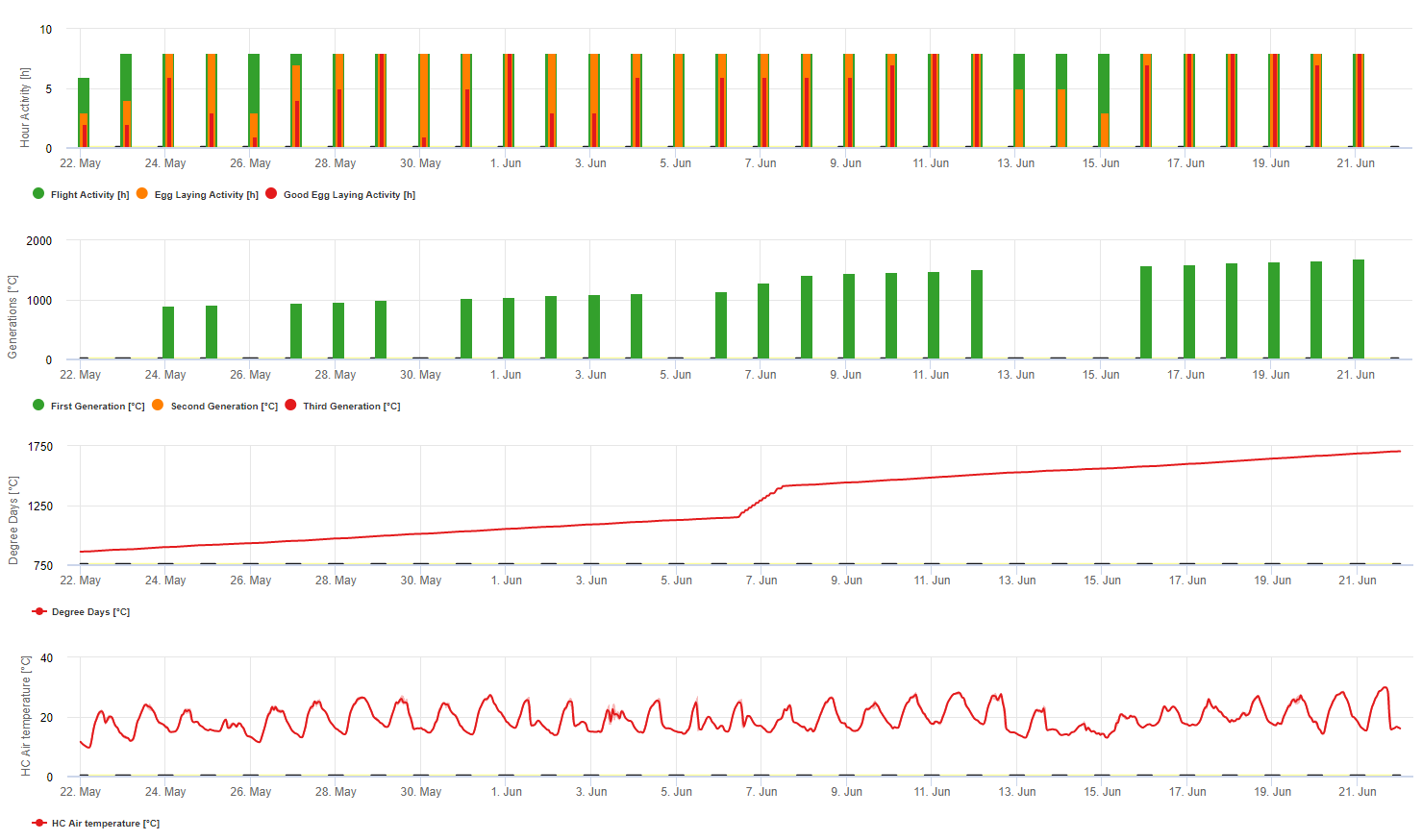
Az európai szőlőbogyó-moly modellezése az FieldClimate platformon
Szükséges érzékelő: Levegő hőmérséklete
Óránkénti adatok Kimenet: repülési aktivitás, tojásrakási aktivitás, első generáció, második generáció és harmadik generáció, felhalmozott foknapok
A repülési aktivitás, a tojás és a jó tojásrakási tevékenység: az utolsó 8 óra adatait ellenőrizzük (19:00 és 03:00 között) A repülési aktivitás küszöbértéke >12°C, a tojásrakási aktivitásé >15°C, a jó tojásrakási aktivitásé >17°C. Ha a tojásrakási aktivitás minden feltétele teljesül, akkor létrehozzuk az első generációt (= felhalmozott foknapok). Az első generáció +530 DD a második generáció (ha a korábbi feltételek miatt nem jósolható). Második generáció + 530 DD a harmadik generáció.
A grafikonon látható az első generáció (jó feltételek mindhárom tevékenységhez, a repüléshez, a tojásrakáshoz és a jó tojásrakáshoz) megjelenése körülbelül 469 fokos napokon halmozódott fel. A második generáció repülése június közepén kezdődik, körülbelül 1010 fokos napok halmozódásával és a tojásrakáshoz szükséges jó hőmérsékleti feltételekkel.
Botrytis cinerea nekrotróf gomba, amely számos növényfajt megtámad, bár legjelentősebb gazdanövénye a szőlő lehet. A szőlészetben általában botritiszes fürtrothadás néven ismert; a kertészetben általában szürkepenésznek vagy szürkepenésznek nevezik.
A gomba kétféle fertőzést okoz a szőlőn. Az első, a szürkerothadás, tartósan nedves vagy párás körülmények között alakul ki, és jellemzően a fertőzött fürtök elvesztését eredményezi. A második, a nemes rothadás, akkor jelentkezik, ha nedvesebb körülményeket szárazabbak követnek, és jellegzetes édes desszertborokat eredményezhet, mint például a Sauternes vagy a tokaji aszú. A faj neve Botrytis cinerea a latin "szőlő, mint a hamu" szóból származik; bár költői, a "szőlő" a gombaspóráknak a konídióforákon való csomósodására utal, a "hamu" pedig csak a spóratömegek szürkés színére utal. A gombát általában anamorf (aszexuális forma) néven szokták emlegetni, mivel az ivaros fázis ritkán figyelhető meg. A teleomorf (ivaros forma) aszkomycéta, Botryotinia cinerea.
A B. cinerea biológiája
A gyümölcsökben és a szőlőben a virágfertőzés, majd a lappangás nagy hatással van a szürkepenész járványtanára. A virágtól a gyümölcsig többféle fertőzési útvonal különíthető el. Szőlőben, kiviben és málnában a petecsomóba történő fertőzést feltételezik. Az ovuluszban a kórokozó látens marad, ami a jelek szerint a gazdaszervezet előre kialakított védekezési stratégiájának eredménye (hasonlóan a fiatal szőlőbogyó resveratroltartalmához). A szőlő, a kivi és a szamóca esetében a porzón, a sziromleveleken vagy a szeptálokon keresztül történő fertőzést találták fontosnak. Szőlőben a vizsgálatok azt mutatták, hogy a B. cinerea a porzókon fertőződhet, és a bazipetális irányban növekszik, hogy megfertőzze a terméshüvelyt, majd szisztémásan a bogyókban a virágszár és az érrendszeri szövetek felé terjedjen.
Egy 6 éves kutatási projekt a Cape térségében kimutatta, hogy a szőlőbogyók fertőződhetnek a száron és a levélnyél lencséin keresztül, valamint a vessző alsó részén. A pedálfertőzés a virágzás időszakában is lehetséges. Később ez a szövet növeli az ellenállóképességet a B. cinerea fertőzések.
Más fertőzési utak a kórokozó szaprofita szaporodását feltételezik a virágtörmeléken, és a bogyók későbbi fertőzését, amikor a fogékonyság az éréssel vagy a bogyók rovar- vagy jégeső okozta károsodásával nő. Egy másik tézis a konídiumok nyári felhalmozódását feltételezi a gyümölcsben, és a fogékony bogyók fertőzését a szezon későbbi szakaszában. Az érő gyümölcsök konídiumfertőzése bármilyen inokulumforrásból lehetséges. Valószínűleg kis számú látensen fertőzött bogyó képződik, amelyek kiterjedt sporulációt mutatnak, amikor a bogyók fogékonysága az éréssel növekszik. Feltételezhető, hogy a bogyók 6% cukortartalomtól kezdve válnak fogékonnyá.
A kiwi gyümölcsöknél a szedési körülményeknek nagy hatása van a következők előfordulására B. cinerea. A nedves felülettel szedett gyümölcsök megfertőződhetnek a következőkkel B. cinerea a szedők ujjai által okozott mikrosebeknél.
A gyakorlati ellenőrzés során B. cinerea el kell különítenünk két fontos fertőzési időszakot: A virágzás és az öregedés. 1) A virágzás alatt vannak fogékony fiatal termések, ahol a fertőzést latenciaidő követi. 2) Míg az érett (szeneszcens) gyümölcsök fertőzése latenciaidő nélkül vezet tünetekhez. A virágzás alatti fertőzés jelentősége a szőlőben szezononként és régiónként változhat. Azokon a gyümölcsökön, amelyeknél számolnunk kell némi eltarthatósági idővel (csemegeszőlő, kivi vagy eper) a tünetek akkor jelentkeznek, ha hűtött körülmények között tároljuk őket üzletekben vagy raktárakban. A szőlő jó ellenállóképességet mutató szőlőfajták kémiai védekezése a következőkkel szemben B. cinerea a virágzás alatt nem fog gazdasági eredményt hozni. Ezért a fertőzés kockázatának és valószínűségének minden körülményét, a gyümölcs fogékonyságát és eltarthatóságát, a tárolási feltételeket figyelembe kell venni a fertőzés elleni kérelemről szóló döntés meghozatalakor. Botrytis cinerea virágzás alatt.
Csonthéjas gyümölcsökben a fertőzés B. cinerea főként virágzáskor fordul elő. Ebben az időszakban a kezeléseket a Monilina spp. figyelembe veszik, amelyek szintén megfertőzik a Botrytis cinerea fertőzés.
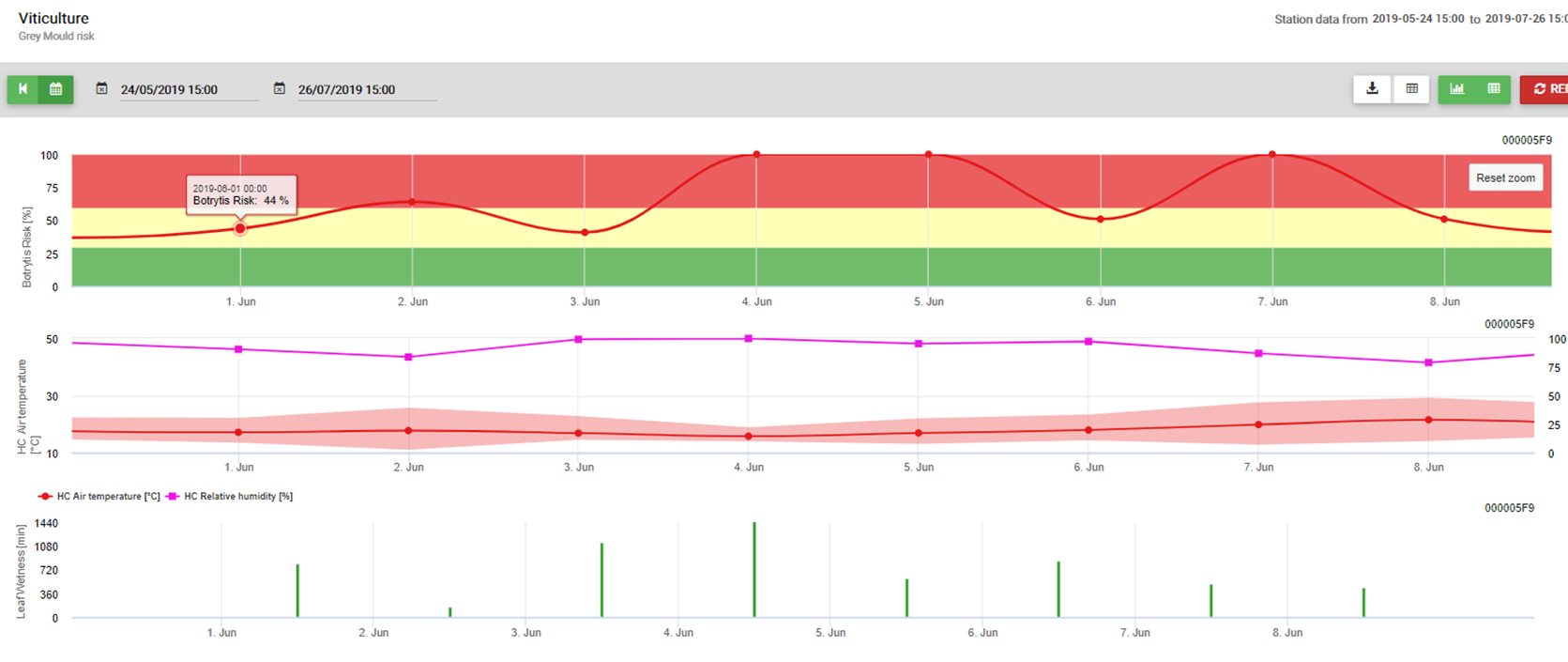
A modell B. cinerea és gyakorlati felhasználás
Szükséges érzékelők: Levélnedvesség, hőmérséklet, relatív páratartalom
Botrytis cinerea fakultatív parazita. Elhalt növényi anyagokon is megterem. Emiatt mindig jelen van a szőlőkben és gyümölcsösökben. A Botrytis cinerea a nedves éghajlathoz kötődik. A fertőzéshez nagyon magas relatív páratartalomra vagy szabad víz jelenlétére van szüksége (érzékelő: levélnedvesség, relatív páratartalom). A gomba nem képes spórák útján megfertőzni az egészséges kifejlett növényi anyagot. A fertőzés a szőlő fiatal hajtásain hosszabb nedves időszakokban vagy a jégeső által károsított hajtásokon történik.
A modell a következő összefüggést használja a levélnedvesség időtartama és a hőmérséklet között a fertőzés kockázatának kiszámításához.
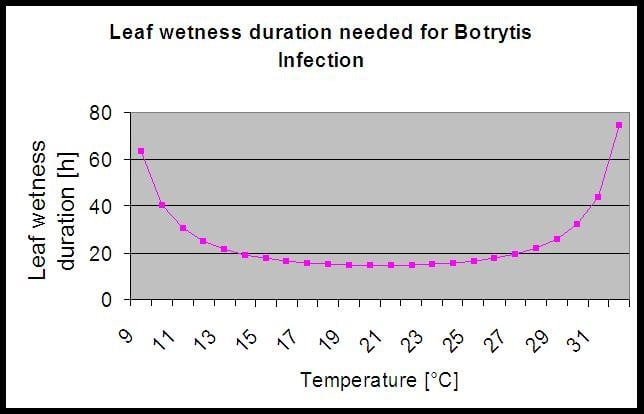
A Botrytis A kockázati modell 0 és 100% közötti kockázati értéket eredményez. Ez az érték jelzi a nyomást B. cinerea akkoriban. Ha 100% értéket kapunk, ez azt jelenti, hogy többször volt elég hosszú nedves időszak ahhoz, hogy megfertőzze a fogékony szöveteket (úgynevezett "nedves pontokat" számolunk (a levélnedvesség és a hőmérséklet közötti tömb, amelynek maximuma kezdetben 38400 pont (a szezon kezdete, amely 30% kockázatot mutat). Ezt követően minden egyes nedves időszak kb. 4000 nedves ponttal (tömb) növeli a kockázatot 10%-vel, vagy másrészt minden egyes száraz időszak csökkenti a kockázatot az előző érték 1/5-ével. Egy alkalmazás a
A kérelem a B. cinerea a terméstől és a termelési céltól függ.
Az antraknózis, a fekete foltosság, a madárszem rothadás, a madárszem foltosság világszerte megtalálható. A betegség csökkenti a gyümölcsminőséget és a terméshozamot, az új hajtások és levelek jelentős pusztulása révén gyengíti a szőlőt. A legtöbb Vitis faj fogékony az antraknózisra.
A párás szőlőtermesztési időszakokban a betegség jól elterjedt, a korai szezonban esők után a nagyon fogékony fajtákban okoz károkat.
A szőlőültetvényekben egyszer már megtelepedett betegség nagyon nehezen kezelhető. Európában azonban a betegség szinte elhanyagolható, mivel a rézvegyületeket, ditiokarbamátokat és ftalimideket tartalmazó gombaölő szerekkel a lisztharmat ellen védekeznek, és a száraz éghajlaton kevés a csapadék.
Tünetek
A betegség a szőlő légnemű, zamatos részeit támadja meg, beleértve a fiatal hajtásokat, leveleket, levélnyelet, indákat és fürtöket; a leggyakoribbak a hajtásokon és a bogyókon jelentkező elváltozások. Az első tünetek elszigetelt, apró, kör alakú foltok formájában jelennek meg, amelyek barnássá, beesetté válnak, szürke középponttal és sötét, kerek vagy szögletes peremmel. Néha a tünetek jégeső okozta sérülésnek tűnnek. A leveleken számos apró, barna folt jelenik meg, amelyek középen szürkék, a nekrotikus középpont általában kihullik, "lövészlyukszerű" megjelenést keltve. A fiatal levelek fogékonyabbak a fertőzésekre. A bogyókon apró vörösesbarna, kör alakú foltok alakulnak ki, amelyek kissé beesnek. A központ szürkévé válik, és vörösesbarna vagy fekete szegélyek veszik körül (madárszem).
A kórokozó életciklusa
A betegséget a kórokozó okozza Elsinöe ampelina (de Bary) Shear.
A betegség fő forrása a fertőzött vesszők. A sérülésekben túlélő szkleróciumok vagy micéliumok tavasszal aktiválódnak, és nedves körülmények között (eső vagy 24 órán át tartó harmat), 2°C-40°C közötti hőmérsékleten konídiumokat termelnek. A konídiumokat az eső az új szövetekbe juttatja. Ezek kicsíráznak és új fertőzéseket hoznak létre. A meleg időjárás csökkenti a kezdeti fertőzéshez szükséges nedvesedési időt és a lappangási időt, mielőtt a tünetek a leveleken láthatóvá válnak. A spórák optimális csírázása 25-30°C-on következik be, legalább 3-4 óra szükséges levélnedvesség mellett. A lappangási idő is ebben a hőmérsékleti tartományban a legrövidebb (3-4 nap ideális nedves körülmények között). A 10°C körüli hőmérsékleten történő fertőzések esetén a betegség tünetei körülbelül 14 nap alatt válnak láthatóvá a leveleken.
A fertőzött bogyókon képződött konídiumok vagy aszkospórák, amelyek a szőlőn vagy a padlón telelnek át, szintén okozhatnak elsődleges fertőzéseket. A spórák az új leveleket, hajtásokat, indákat, fiatal bogyókat fertőzik, és nedves körülmények között elváltozásokat okoznak. Ezek a konídiumok másodlagos inokulumként szolgálnak, és a szezonban további fertőzésekért felelősek. A konídiumok az esőcseppek hatására elmozdulnak és szétszóródnak a szőlőben.
A járvány kialakulását a szőlőszövet fogékonysága, a konídiumok terjedése és az uralkodó időjárási viszonyok okozzák. A legfontosabb tényező az esős időszakok gyakorisága.
Modellezés Fieldclimate-ban
Túlélő spórák fejlődése: Hőmérséklet: 2-40C°, relatív páratartalom 90% felett vagy levélnedvesség - amikor a spórák kifejlődtek (100%) és még mindig a levélnedvesség és a hőmérséklet feltételei mellett kezdődik a fertőzés kiszámítása. A spórák fejlődése (relatív páratartalom 50% alatt) és a fertőzés leáll, ha a páratartalom túl alacsony. A fertőzés súlyossága a nedves körülményektől (esőesemény) függ.