





La fusariose des épis ou la gale des petits grains est causée par le champignon Fusarium graminearum (Schwabe), bien que les Pays-Bas et d'autres régions d'Europe centrale fassent état d'une baisse de la consommation d'eau. F. culmorum comme l'espèce la plus répandue (Snidjers, 1989). En Pologne, F. culmorum, F. graminearum et F. nivale ont montré des niveaux de virulence modérés à sévères similaires, tandis que les F. avenaceum s'est avéré moyennement ou modérément virulent. Cependant, dans plusieurs études visant à identifier l'organisme responsable, jusqu'à 18 Fusarium spp. ont été isolés et identifiés (Mihuta-Grimm et Foster, 1989 ; Reis, 1985).
Article de L. GilchristH.J. Dubin La tavelure est présente dans les régions chaudes et humides où la floraison coïncide avec les périodes de pluie. L'incidence de cette maladie a augmenté au cours des dix dernières années pour diverses raisons. La raison la plus importante est peut-être l'augmentation de la superficie où le blé est cultivé en rotation avec du maïs ou d'autres céréales. D'autres raisons sont les changements dans le système de culture à des fins de protection du sol et les changements dans la culture du blé, qui passe des zones traditionnelles à des zones plus humides et non traditionnelles (Gilchrist et al., 1997).
La gale fusarienne entraîne de graves pertes de production dans le monde entier, qui peuvent atteindre 50 pour cent. De même, au Paraguay, les conditions climatiques de 1972 et 1975 ont favorisé les épidémies de Fusarium et de Septoria qui, ensemble, ont causé des pertes allant jusqu'à 70 pour cent (Viedma, 1989). La Pologne, les Pays-Bas, le Royaume-Uni, l'ex-Tchécoslovaquie, la Fédération de Russie, la France et l'Autriche sont quelques-uns des pays européens qui signalent des cas de tavelure. Des régions du Canada, des Etats-Unis, du Mexique, du Guatemala, du Brésil, de l'Equateur, de l'Uruguay et de l'Argentine dans les Amériques sont fortement touchées par Fusarium (Ireta et Gilchrist, 1994).
La tavelure peut causer des dommages importants au niveau du rendement et de la qualité, ainsi que des toxicoses chez les animaux et les humains (Ireta et Gilchrist, 1994 ; Baht et al., 1989 ; Luo, 1988 ; Snidjers, 1989 ; Marasas et al., 1988). Les dégâts causés par la tavelure aux Etats-Unis ont été estimés à plus de 1,23 milliard de dollars US en 1993 et à 1,23 milliard de dollars US en 1994. En Chine, on estime que la tavelure peut affecter jusqu'à 7 millions d'hectares et que 2,5 millions de tonnes de céréales peuvent être perdues au cours des années épidémiques. Des maladies liées à la mycotoxine fusarienne ont été signalées chez l'homme en Chine, en Inde et au Japon, tandis que des maladies animales ont été signalées dans de nombreuses régions du monde (Dubin et al., 1997).
Aujourd'hui, il existe des réglementations mondiales pour les mycotoxines (Van Egmond et Dekker, 1995). Cependant, dans de nombreux pays, les réglementations ne sont pas appliquées et de nombreuses personnes, en particulier dans les zones rurales, consomment les céréales sans aucun contrôle, soit sous forme de grains de céréales, soit sous forme indirecte, sous forme de viande provenant d'animaux nourris avec des grains contaminés. Le tableau 16.1 présente les principaux effets sur les porcs et les volailles induits par les toxines les plus importantes produites par certaines céréales. Fusarium espèces.
Les conditions climatiques humides et chaudes entre l'épiaison et la maturité augmentent la gravité de la tavelure. Le point d'entrée de F. graminearum est l'épi, en particulier les organes floraux. Cela affecte la formation des graines et le remplissage des grains. Les épillets infectés perdent rapidement leur chlorophylle et prennent une couleur pâle. Plus tard, ils prennent une couleur rose ou pêche, en particulier à la base et sur les bords des glumes (planche 41). Si les conditions environnementales restent favorables, l'infection progresse vers les épillets adjacents et, dans certains cas, peut infecter l'ensemble de l'épi, y compris le rachis et son pédoncule. Lorsque l'infection est sévère, les grains endommagés sont couverts de mycéliums et prennent l'aspect d'une masse cotonneuse rose. Si la maladie est modérée, le grain peut être ratatiné, de faible poids et de couleur blanchâtre (grains de pierre tombale).
Principaux effets toxiques sur les porcs et les volailles induits par les toxines les plus importantes produites par certaines espèces de Fusarium
| Mycotoxine | Signes cliniques |
| Zéaralénone | Vulve rouge et gonflée ; prolapsus vaginal chez les porcs ; les porcelets allaités peuvent présenter une hypertrophie de la vulve ; problèmes de fertilité. |
| Vomitoxine (désoxynivalénol, DON) | Diminution de la consommation d'aliments et de la prise de poids chez les porcs ayant reçu >2 mg/kg d'aliments ; vomissements ; refus de s'alimenter à des concentrations très élevées de DON (>20 mg/kg d'aliments)a |
| Autres trichothécènes Toxine T-2 Toxine HT-2 Diacétoxyescirpénol > | Plus toxique que le DON ; réduction de la consommation d'aliments ; vomissements ; irritation cutanée et gastro-intestinale ; neurotoxicité ; progéniture anormale ; sensibilité accrue aux maladies ; hémorragies. |
| Ocharatoxine | Affecte principalement les tubules proximaux des reins chez les porcs et les volailles ; les reins sont grossièrement élargis et pâles ; foie gras chez les volailles. |
amg/kg = parties par million (ppm).
Source : Trenholm et al : Trenholm et al, 1984.
Fusarium graminearum (planche 42) peut attaquer les plantes de blé à tous les stades de croissance, provoquant des maladies des semis, des tiges et des racines. Les infections primaires peuvent provenir des ascospores ou des macroconidies déposées sur les glumes et les anthères extrudées. Des températures de 10° à 30°C et une humidité relative supérieure à 95% pendant 40 à 60 heures sont généralement suffisantes pour que les macroconidies infectent avec succès les épis (Ireta, 1989).
Fusarium graminearum est l'un des rares Fusarium qui produit des périthèces en conditions de terrain (Planche 43, Planche 44). Les périthèces représentent le stade sexuel du champignon, Gibberella zeae, et sont produits sur les glumes de blé. Les périthèces jouent un rôle important dans la survie du pathogène d'une année sur l'autre (Khonga et Sutton, 1988) et coexistent avec les mycéliums dans les résidus de la culture précédente pour constituer la source initiale d'inoculum pour la tavelure. Des recherches effectuées en Chine ont montré que la température la plus basse pour la production de périthèces était de 7 à 10°C, et la plus appropriée de 15 à 20°C. Le taux d'humidité du sol le plus bas pour la production de périthèces était de 50 à 60 %, et le plus approprié de 70 à 80 % (Wang, 1997).
Les facteurs qui déterminent le développement de la maladie sont le climat, les niveaux d'inoculum et le stade de croissance du blé. Les épidémies de gale du blé dépendent principalement de la quantité d'inoculum primaire, plutôt que de l'inoculum secondaire. L'invasion se produit principalement pendant la période de floraison. L'infection secondaire par les conidies (planche 45) se produit après l'apparition des épis malades dans le champ.
Les résidus de culture et les pratiques culturales jouent un rôle important dans la préservation de la biodiversité. F. graminearum et, par conséquent, des épidémies. Les infections sur blé semé dans un champ de résidus de maïs peuvent être deux ou trois fois plus graves (Teich et Nelson, 1984). En général, le pathogène n'était saprophyte que sur les chaumes de riz dans les zones de rotation riz-blé et sur les tiges de maïs dans les zones de plantation sèche en Chine (Wang, 1997). Si les résidus sont labourés, la survie des périthèces diminue et réduit la source primaire d'inoculum (Reis, 1989).
Fusarium graminearum est un parasite facultatif et est pathogène sur de nombreuses autres graminées, y compris des adventices communes et des cultures céréalières (seigle, riz, orge et triticale). Si les pratiques telles que la rotation avec des cultures non hôtes ou la gestion des résidus de culture ne sont pas efficaces seules, elles peuvent, en combinaison, réduire la source d'inoculum primaire. La lutte contre la maladie repose effectivement sur une gestion intégrée, comprenant des pratiques agronomiques appropriées, l'utilisation de cultivars résistants ou tolérants et des applications chimiques.
Biologie de la fusariose des épis
Pathogènes occasionnels
La fusariose est causée par des espèces fongiques du genre Fusarium. L'espèce la plus courante est Fusarium graminearum (stade sexuel - Gibberella zeae). Ce champignon est le même que celui qui est fréquemment associé à la pourriture des tiges de maïs. Un autre champignon Fusarium L'espèce responsable de la fusariose est Fusarium culmorum. Les deux F. graminearum et F. culmorum peut également provoquer la pourriture des racines des petites céréales. Sur l'orge, deux autres Fusarium espèces, F. poae et F. avenaceumpeut également provoquer une brûlure des grains.
Survie et propagation
Le champignon persiste et se multiplie sur les résidus de culture infectés de petites céréales et de maïs. Par temps humide, les spores des champignons sont soufflées par le vent ou projetées sur les épis des cultures céréalières. Les spores peuvent provenir de l'intérieur d'une culture ou être soufflées depuis les cultures environnantes, parfois sur de longues distances. Les cultures de blé et de blé dur sont susceptibles d'être infectées depuis la période de floraison (pollinisation) jusqu'au stade de la pâte dure du développement du grain. Les spores du champignon responsable peuvent se déposer sur les anthères exposées au moment de la floraison et se développer ensuite dans les grains, les glumes ou d'autres parties de l'épi. Pour l'orge de printemps, qui fleurit lorsque l'épi est dans la botte, l'infection est plus fréquente après la période de floraison, une fois que l'épi traverse la gaine foliaire. L'infection dans l'une ou l'autre culture peut se poursuivre jusqu'à une date proche de la maturité du grain si les conditions environnementales sont favorables à l'organisme (ou aux organismes).
Les conditions les plus favorables à l'infection sont des périodes prolongées (48 à 72 heures) d'humidité élevée et de températures chaudes (75 à 85 degrés Fahrenheit (24°C à 30°C)). Cependant, l'infection peut se produire à des températures plus fraîches lorsque l'humidité élevée persiste pendant plus de 72 heures. Les infections précoces peuvent produire des spores transportées par l'air, qui sont responsables de la propagation secondaire de la maladie, en particulier si la culture présente une floraison inégale en raison de talles tardives.
Étant donné que le développement de la fusariose dépend de conditions environnementales favorables depuis la floraison (émergence des épis chez l'orge) jusqu'au développement des grains, l'apparition et la gravité de la maladie varient d'une année à l'autre. Une combinaison de facteurs peut conduire aux pertes de rendement et de qualité les plus sévères : un inoculum abondant, des périodes prolongées ou répétées d'humidité et de forte hygrométrie pendant la floraison (émergence des épis chez l'orge) jusqu'au développement des grains, et l'utilisation d'un cultivar très sensible.
Modèle d'infection par la fusariose de l'épi
Les agents fongiques appartenant au complexe Fusarium Head Blight sur le blé sont connus pour être favorisés par des températures chaudes de 20 °C à 30 °C et de longues périodes d'humidité. Des périodes d'humidité foliaire de plusieurs jours conduisent à des symptômes visibles précoces. Mais les symptômes peuvent apparaître après une période de latence plus longue si l'infection est suivie d'une période d'humidité des feuilles de 18 heures ou même moins. Des symptômes peuvent également être observés après une inoculation artificielle à des températures de 15°C.
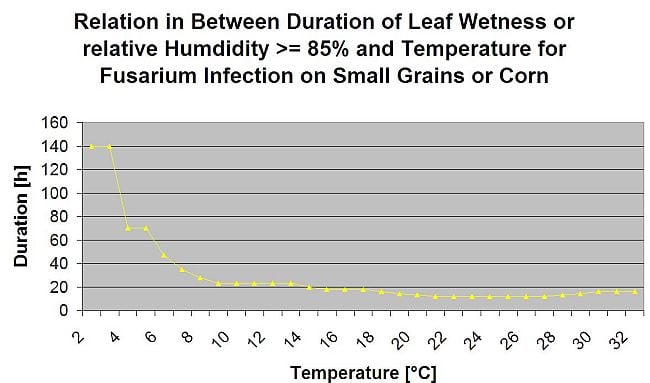

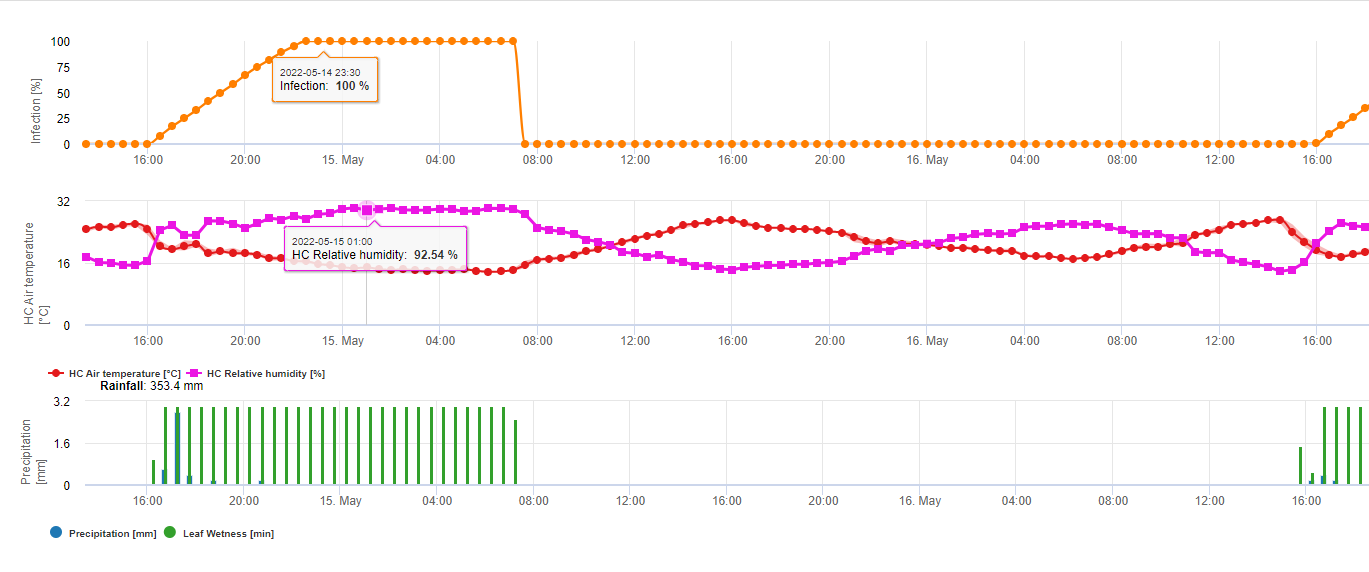
En résumant toutes les différentes combinaisons de températures et d'humidité que nous avons trouvées dans la littérature, nous avons décidé de signaler les infections par la fusariose de l'épi si la température et la période d'humidité des feuilles ou les périodes avec plus de 85% d'humidité relative dépassent les valeurs visualisées dans le graphique suivant.
Les infections sont déclenchées par une pluie de 2 mm. On peut supposer qu'il s'agit d'une infection par la fusariose de l'épi si la valeur de la progression de l'infection atteint 100%. Le calcul de cette valeur de progression suit la relation entre la durée des conditions humides et la température affichée dans le graphique ci-dessus.
Ce modèle est utilisé pour visualiser les jours d'infection et les conditions climatiques pendant le climat. La connaissance par les cultivateurs du stade de développement des différentes variétés de blé permet de décider de l'application d'une pulvérisation curative immédiatement après l'infection.
Alerte aux mycotoxines du fusarium
Les essais d'infection avec des périodes prolongées d'humidité foliaire de la fusariose de l'épi ont conduit à des teneurs élevées en mycotoxines. D'après ces informations, une période d'humidité foliaire de 48 heures ou plus au cours des stades 61 et 69 est considérée comme présentant un risque élevé de mycotoxines. Les expériences faites au cours des analyses de DON dans le blé cultivé commercialement ont montré que des périodes d'humidité foliaire suffisamment longues pour l'infection après une infection initiale dans les stades 61 à 69 peuvent également augmenter les valeurs de DON. En cas de périodes d'humidité foliaire plus longues, les mycotoxines peuvent augmenter jusqu'au stade 85. FieldClimate accumule un chiffre de risque proportionnel à la progression de l'infection pour chaque période d'infection réussie au cours de la période qui a été sélectionnée pour être appropriée pour ces calculs. 6 périodes d'infection tout juste terminées conduiraient à un risque de 100%. Normalement, la période d'humidité des feuilles menant à une infection par le fusarium est plus longue que le minimum nécessaire. Par conséquent, la plupart des infections par le fusarium entraîneront une augmentation du risque de plus de 17%. La valeur de risque indiquant une situation problématique en matière de mycotoxines dépend de l'historique du champ. Le blé cultivé après un maïs non travaillé ou le blé non travaillé après un maïs non travaillé ne présente qu'un faible risque s'il n'est pas pulvérisé dans des conditions optimales. Dans le cas du blé non pulvérisé, il faut s'attendre à une augmentation des valeurs de DON dès 35% de risque. Le blé qui suit une culture autre que le maïs ou le blé sans travail du sol peut présenter un risque plus élevé de 50%. Si le blé suit une culture de maïs ou de blé avec travail du sol, le risque accru peut atteindre 70%. Le blé de première année doit être testé pour le DON si le risque dépasse 100%.
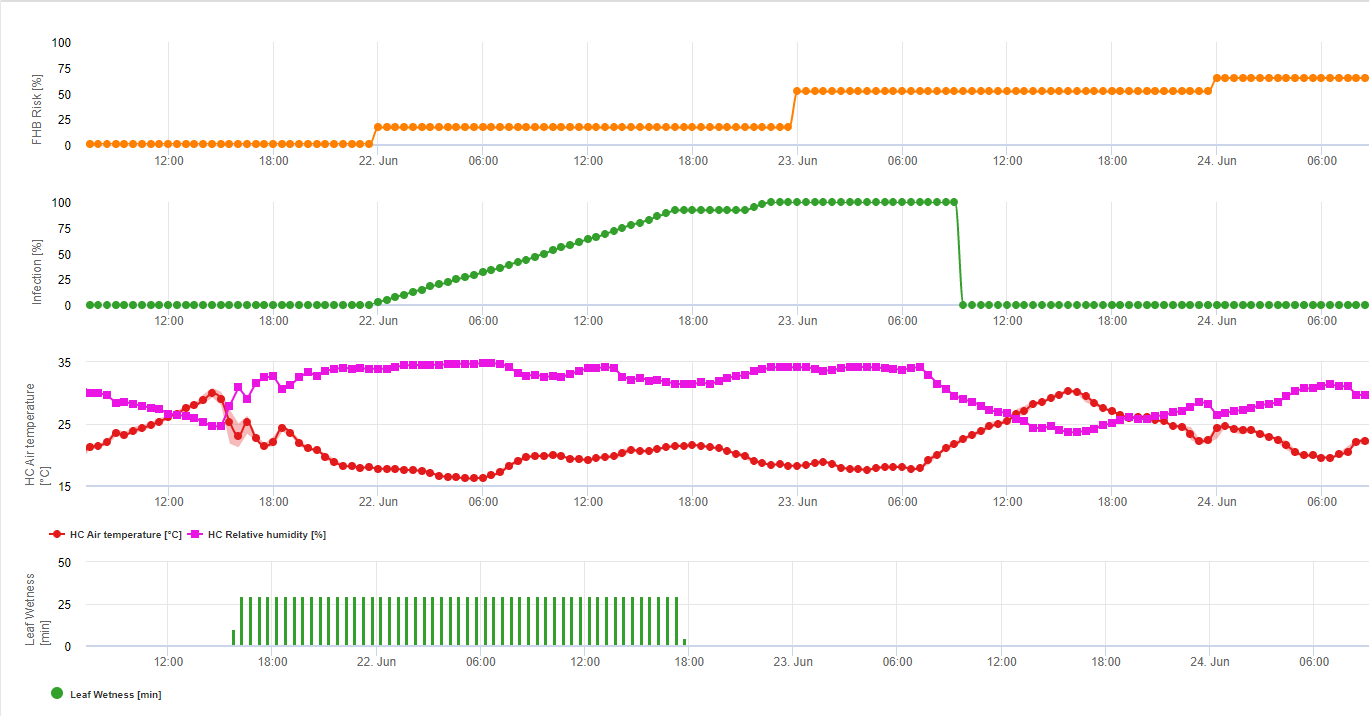
1) Modèle de risque pour la fusariose de l'épi qui détermine les périodes à risque pour une infection. Lorsque l'infection 100% (ligne verte) est atteinte, le risque (ligne bleue) est très élevé et les conditions pour le champignon ont été favorables à l'infection. En fonction de la méthode d'application (curative, préventive), la période à risque est indiquée par la ligne bleue.
2) Fusariose de l'épi : Dans ce modèle, l'infection de la fusariose est calculée en fonction des précipitations (2 mm nécessaires), de l'humidité relative (supérieure à 85%) ou de l'humidité des feuilles, de la température pendant le processus. Si l'infection atteint 100%, les conditions optimales pour le champignon pathogène sont réunies. Le modèle calcule ensuite le risque de mycotoxine FHB.
Il existe deux principales maladies du blé dues à la septoriose. Il s'agit de la tache Septoria tritici, provoquée par le champignon Septoria tritici (téléomorphe : Mycophaerella graminicola) et Septoria nodorum blotch, causée par le champignon Septoria nodorum (téléomorphe : Leptosphaeria nodorum). Ces deux maladies entraînent de graves pertes de rendement allant de 31 à 53 pour cent (Eyal, 1981 ; Babadoost et Herbert, 1984 ; Polley et Thomas, 1991). Dans le monde entier, plus de 50 millions d'hectares de blé, poussant principalement dans les régions à forte pluviométrie, sont touchés. Au cours des 25 dernières années, ces maladies ont augmenté et sont devenues un facteur limitant majeur de la production de blé dans certaines régions. En cas d'épidémies graves, les grains des cultivars de blé sensibles sont ratatinés et impropres à la mouture. Les épidémies de Septoria tritici et de Septoria nodorum sont associées à des conditions climatiques favorables (pluies fréquentes et températures modérées), à des pratiques culturales spécifiques, à la disponibilité d'inoculum et à la présence de cultivars de blé sensibles (Eyal et al., 1987).
Septoria spp. Biologie
Suite à Erick De Wolf, Septoria Tritici Blotch, Kansas State University, avril 2008 Septoria tritici est causée par un champignon, l'acarien, qui est à l'origine de la tache des feuilles. Septoria tritici. Elle est présente dans toutes les zones de culture du blé du monde et constitue un problème sérieux dans de nombreuses régions. La tache Septoria tritici est la plus dommageable lorsque la maladie attaque les feuilles supérieures et les épis des variétés sensibles en fin de saison.
Symptômes
Septoria tritici Les symptômes de la tache noire apparaissent pour la première fois à l'automne. Les premiers symptômes sont de petites taches jaunes sur les feuilles. Ces lésions deviennent souvent légèrement bronzées en vieillissant, et les fructifications fongiques peuvent être observées dans les lésions sur les tiges. Les lésions sont de forme irrégulière et varient d'elliptiques à longues et étroites (figure 1). Les lésions contiennent de petites taches rondes et noires qui sont les fructifications du champignon. Les fructifications noires ressemblent à des grains de poivre noir et peuvent généralement être vues sans l'aide d'une loupe. La maladie commence sur les feuilles inférieures et progresse progressivement jusqu'à la feuille étendard. Les gaines des feuilles sont également susceptibles d'être attaquées. Les années humides, le champignon de la tache mouchetée peut se déplacer sur les têtes et provoquer des lésions brunes sur les glumes et les aulnes, connues sous le nom de tache des glumes. Ces lésions deviennent souvent légèrement bronzées en vieillissant et les fructifications fongiques sont souvent visibles dans les lésions sur les aulnes.
La phase de la tache des glumes peut entraîner une perte de rendement importante, mais la relation entre la gravité de la maladie et la perte de rendement n'est pas bien comprise. Septoria tritici peut être confondue avec d'autres maladies foliaires qui présentent des symptômes très similaires : la tache bronzée et la tache de l'épinette. Stagonspora nodorum par exemple. Il est fréquent que les plantes soient infectées par plusieurs de ces maladies foliaires, et il peut être nécessaire de procéder à des examens de laboratoire pour diagnostiquer avec précision les maladies les plus répandues. Un examen en laboratoire est presque toujours nécessaire pour déterminer la cause de la tache des glumes. La connaissance de l'espèce n'est pas importante pour les décisions de pulvérisation, car les trois maladies réagissent de la même manière aux fongicides. Cependant, le fait de savoir quelles sont les maladies les plus répandues est un élément important de la sélection des variétés, car différents gènes contrôlent la résistance aux maladies.
La manière la plus fiable de distinguer la tache septoriale des autres maladies est la présence de fructifications noires. Le champignon responsable de la tache bronzée ne produit pas ce type de structure reproductive. En revanche, dans des conditions humides, le champignon responsable de la tache Stagonospora nodorum produit des fructifications brun clair. Outre la différence de couleur, ces structures sont également plus petites que celles produites par Septoria tritici.
Cycle de vie
Septoria tritici survit pendant l'été sur les résidus d'une culture de blé précédente et commence les infections à l'automne. Il existe des preuves que le champignon est capable de survivre en association avec d'autres graminées hôtes et des semences de blé. Ces sources du champignon sont probablement les plus importantes lorsque les résidus de blé sont absents. Indépendamment des pratiques de rotation ou de gestion des résidus, il y a généralement suffisamment d'inoculum pour déclencher des infections à l'automne. Septoria tritici est favorisée par un temps frais et humide. La température optimale se situe entre 16 et 21 °C ; cependant, des infections peuvent se produire pendant les mois d'hiver à des températures aussi basses que 5 °C. L'infection nécessite au moins 6 heures d'humidité des feuilles, et jusqu'à 48 heures d'humidité pour une infection maximale. Une fois que l'infection a eu lieu, le champignon met 21 à 28 jours pour développer les fructifications noires caractéristiques et produire une nouvelle génération de spores. Les spores produites dans ces corps fructifères sont exsudées en masses collantes et il faut de la pluie pour les projeter sur les feuilles supérieures et les têtes.
Infection par Septoria tritici
Pycnidiospores de S. tritici germent dans l'eau libre à partir des deux extrémités de la spore ou des cellules intercalaires (Weber, 1922). La germination des spores ne commence qu'environ 12 heures après le contact avec la feuille. Les tubes germinatifs se développent au hasard sur la surface de la feuille. Weber (1922) n'a observé qu'une pénétration directe entre les cellules épidermiques, mais d'autres ont conclu que la pénétration à travers les stomates ouverts et fermés est le principal moyen de pénétration de l'hôte (Benedict, 1971 ; Cohen et Eyal, 1993 ; Hilu et Bever, 1957). Kema et al. (1996) n'ont observé que la pénétration stomatique. Les hyphes qui se développent à travers les stomates se resserrent jusqu'à un diamètre d'environ 1 μm, puis s'élargissent après avoir atteint la cavité sous-matale.
Les hyphes se développent parallèlement à la surface de la feuille sous les cellules épidermiques, puis à travers le mésophylle jusqu'aux cellules de l'épiderme inférieur, mais pas dans l'épiderme. Il n'y a pas de formation d'haustoria et la croissance des hyphes est limitée par les cellules du sclérenchyme autour des faisceaux vasculaires, sauf lorsque les hyphes sont très denses. Les faisceaux vasculaires ne sont pas envahis. Les hyphes se développent de manière intercellulaire le long des parois cellulaires à travers le mésophylle, se ramifiant au niveau d'un septum ou du milieu d'une cellule. Aucun symptôme macroscopique n'apparaît pendant environ 9 jours, à l'exception d'une cellule morte occasionnelle, mais les cellules du mésophylle meurent rapidement après 11 jours. Les pycnides se développent dans des chambres sous-matricielles. Les hyphes se développent rarement dans les cellules de l'hôte (Hilu et Bever, 1957 ; Kema et al, 1996 ; Weber, 1922).
L'infection ne réussit qu'après au moins 20 heures d'humidité élevée. Seules quelques mouchetures brunes se développent si les feuilles restent humides pendant 5-10 heures après le dépôt des spores (Holmes et Colhoun, 1974) ou jusqu'à 24 heures (Kema et al., 1996). Les relations hôte-parasite sont les mêmes sur les blés résistants ou sensibles. La germination des spores à la surface des feuilles est la même quelle que soit la sensibilité. Le nombre de pénétrations réussies est à peu près le même, mais la croissance des hyphes est plus rapide chez les cultivars sensibles, ce qui entraîne un plus grand nombre de lésions. Les hyphes s'étendent au-delà de la zone nécrosée chez tous les cultivars. Une toxine peut jouer un rôle dans la pathogenèse (Cohen et Eyal, 1993 ; Hilu et Bever, 1957). En revanche, la colonisation a été fortement réduite sur une lignée résistante (Kema et al., 1996).
Stagonospora (Septoria) et Septoria pathogènes des céréales : Le processus d'infection
B.M. Cunfer, Département de pathologie végétale, Université de Géorgie, Griffin, GA
Le processus d'infection a fait l'objet des études les plus approfondies pour Stagonospora (Septoria) nodorum et Septoria tritici. Une étude approfondie sur Septoria passerinii est disponible. Presque toutes les informations rapportées concernent l'infection par les pycnidiospores. Cependant, le processus d'infection pour d'autres formes de spores est assez similaire. Les informations présentées concernent principalement l'infection des feuilles dans des conditions optimales. Certaines études ont été réalisées avec des plants intacts, tandis que d'autres ont été réalisées avec des feuilles détachées. Infection du coléoptile et de la plantule de blé par des S. nodorum a été décrite en détail par Baker (1971) et analysée par Cunfer (1983). Bien qu'aucune comparaison précise n'ait été faite, il semble que le processus d'infection présente de nombreuses similitudes dans chaque système hôte-parasite et soit typique de nombreux pathogènes nécrotrophes. Les informations sur les facteurs influençant le développement des symptômes et l'expression de la maladie sont exclues mais ont été examinées par d'autres auteurs (Eyal et al., 1987 ; King et al., 1983 ; Shipton et al., 1971). Un résumé des facteurs affectant la longévité des spores à la surface des feuilles est inclus.
Rôle des cirrus et survie des spores sur la surface de la feuille Les informations les plus détaillées sur la fonction du cirrus enveloppant les pycnidiospores exsudées par le pycnidium sont pour S. nodorum. Le cirrus est un gel composé d'éléments protéiques et saccharidiques. Sa composition et sa fonction sont similaires à celles d'autres champignons des Sphaeropsidales (Fournet, 1969 ; Fournet et al., 1970 ; Griffiths et Peverett, 1980). Les rôles principaux des composants du cirrus sont la protection des pycnidiospores contre la dessiccation et la prévention de la germination prématurée.
Le cirrus protège les pycnidiospores de telle sorte que certaines restent viables pendant au moins 28 jours (Fournet, 1969). Lorsque le cirrus est dilué avec de l'eau, si la concentration de la solution de cirrus est >20%, moins de 10% de pycnidiospores germent. À une concentration plus faible, les composants fournissent des nutriments qui stimulent la germination des spores et l'allongement des tubes germinatifs. La longueur des tubes germinatifs augmente jusqu'à une concentration de 15% cirrus, puis diminue modérément à des concentrations plus élevées (Harrower, 1976). Brennan et al. (1986) ont rapporté une germination plus importante dans un cirrus dilué. Les composants du cirrus réduisent la germination à 10-60% d'humidité relative. Une fois que les spores sont dispersées, les effets stimulants du liquide de cirrus sont probablement négligeables (Griffiths et Peverett, 1980).
À une humidité relative de 35-45%, les spores de S. tritici dans les cirres sont restées viables pendant au moins 60 jours (Gough et Lee, 1985). Les composants du cirrus peuvent agir comme un inhibiteur de la germination des spores, ou le potentiel osmotique élevé du cirrus peut empêcher la germination. Les pycnidiospores de S. nodorum n'ont pas survécu pendant 24 heures à une humidité relative supérieure à 80% à 20 C. Les spores ont survécu deux semaines ou plus à une humidité relative <10% (Griffiths et Peverett, 1980). Lorsque le fluide de cirrus de S. nodorum diluées dans l'eau, environ deux tiers des pycnidiospores ont perdu leur viabilité en 8 heures, et après 30 heures à la lumière du jour, seules 5% ont germé. Lorsque les spores sont conservées dans l'obscurité, 40% restent viables après 30 heures (Brennan et al., 1986).
Conidies sèches de S. nodorumLes pycnidiospores de S. nodorum, à l'ombre et en plein soleil, ont survécu à l'extérieur pendant au moins 56 heures (Fernandes et Hendrix, 1986a). La germination des pycnidiospores de S. nodorum a été inhibée par des UV-B continus (280-320 nm), alors que la germination des pycnidiospores de S. nodorum a été inhibée par des UV-B continus (280-320 nm). S. tritici ne l'était pas. L'extension des tubes germinatifs sous UV-B continu a été inhibée pour les deux champignons, par rapport à l'obscurité (Rasanayagam et al., 1995).
Infection par Septoria nodorum
Le processus de pénétration de l'hôte et le développement des S. nodorum La pénétration des pycnidiospores dans la feuille a été examinée en détail par plusieurs chercheurs (Baker et Smith, 1978, Bird et Ride, 1981, Karjalainen et Lounatmaa, 1986 ; Keon et Hargreaves, 1984 ; Straley, 1979 ; Weber, 1922). Les pycnidiospores ont tendance à se loger dans les dépressions entre deux cellules épidermiques, et de nombreuses tentatives de pénétration des feuilles commencent par là. Les spores germent à la surface des feuilles en réponse à l'humidité libre (Fernandes et Hendrix, 1986b). Elles commencent à germer 2-3 heures après le dépôt, et après 8 heures la germination peut atteindre 90%. La pénétration des feuilles commence environ 10 heures après le dépôt des spores (Bird et Ride, 1981 ; Brönnimann et al., 1972 ; Holmes et Colhoun, 1974).
Au début de la germination, le tube germinatif est entouré d'une matière amorphe qui s'attache à la feuille. Les tubes germinatifs qui poussent à partir des deux extrémités d'une spore et des cellules intercalaires ont tendance à se développer le long des dépressions entre les cellules et sont souvent orientés le long de l'axe long de la feuille (O'Reilly et Downes, 1986). Les hyphes provenant de spores qui ne sont pas dans des dépressions poussent de manière aléatoire avec des ramifications occasionnelles (Straley, 1979). Un appressorium se forme avec une tige d'infection qui pénètre la cuticule et les parois périclinales des cellules épidermiques directement dans le lumen cellulaire, ce qui entraîne la mort rapide des cellules.
De nombreuses pénétrations sont d'abord subcuticulaires ou la croissance latérale d'un hyphe se produit à l'intérieur de la paroi cellulaire avant la croissance dans le cytoplasme (Bird et Ride, 1981 ; O'Reilly et Downes, 1986). La pénétration à travers les stomates ouverts ou fermés se produit également et peut être plus rapide que la pénétration directe (Harrower, 1976 ; Jenkins, 1978 ; O'Reilly et Downes, 1986 ; Straley, 1979). Les tubes germinatifs se ramifient au niveau des stomates et des jonctions de cellules épidermiques. La pénétration d'un tube germinatif dans un stomate peut se produire sans formation d'un appressorium. La pénétration se fait parfois à travers les trichomes (Straley, 1979). Apparemment, la plupart des tentatives de pénétration échouent, des papilles denses se formant dans les cellules au site de la tentative de pénétration (Karjalainen et Lounatmaa, 1986 ; Bird et Ride, 1981).
Après la pénétration, les cellules épidermiques meurent rapidement et se lignifient, et les hyphes se développent dans le mésophylle. Les cellules du mésophylle se déforment et du matériel lignifié se dépose à l'extérieur de certaines cellules, qui s'effondrent alors. La lignification se produit avant que les hyphes n'atteignent la cellule. Le processus est le même dans tous les cultivars mais se développe plus lentement dans les cultivars résistants. Les hyphes se développent de manière intercellulaire entre les cellules épidermiques, puis dans le mésophylle. Lorsque le mésophylle est pénétré, la détérioration des chloroplastes commence en 6-9 jours (Karjalainen et Lounatmaa, 1986).
Cependant, le taux de photosynthèse commence à diminuer un jour après l'infection et avant que les symptômes ne soient visibles (Krupinsky et al, 1973). Le sclérenchyme entourant les faisceaux vasculaires empêche l'infection des tissus vasculaires. Les faisceaux vasculaires bloquent la dissémination des hyphes à travers le mésophylle, sauf lorsque le tissu sclérenchyme est jeune et pas complètement formé (Baker et Smith, 1978).
Stagonospora nodorum libère une large gamme d'enzymes dégradant les parois cellulaires, y compris l'amylase, la pectine méthyl estérase, les polygalacturonases, les xylanases et la cellulase in vitro et pendant l'infection des feuilles de blé (Baker, 1969 ; Lehtinen, 1993 ; Magro, 1984). Les informations relatives à la dégradation des parois cellulaires par les enzymes sont en accord avec les observations histologiques. Ces enzymes peuvent agir en conjonction avec les toxines. La sensibilité aux enzymes peut être liée à la résistance et au taux de colonisation fongique (Magro, 1984). Comme de nombreux nécrotrophes, les pathogènes Septoria et Stagonospora produisent des composés phytotoxiques in vitro. La détérioration et la mort des cellules avant la croissance des hyphes dans le tissu mésophylle (Bird et Ride, 1981) sont compatibles avec la production de toxines. Cependant, le rôle définitif des toxines dans le processus d'infection et leur relation avec la résistance de l'hôte n'ont pas été établis (Bethenod et al, 1982 ; Bousquet et al, 1980 ; Essad et Bousquet, 1981 ; King et al, 1983). Les différences de gamme de plantes-hôtes entre les souches de S. nodorum peut être liée à la production de toxines (Bousquet et Kollmann, 1998). L'initiation de la germination des spores et le pourcentage de spores germées ne sont pas influencés par la sensibilité de l'hôte (Bird et Ride, 1981 ; Morgan 1974 ; Straley, 1979 ; Straley et Scharen, 1979 ; Baker et Smith, 1978).
Bird et Ride (1981) signalent que l'extension des tubes germinatifs à la surface des feuilles est plus lente sur les cultivars résistants que sur les cultivars sensibles. Ce mécanisme, qui s'exprime au moins 48 heures après le dépôt des spores, indique une résistance à l'élongation des tubes germinatifs avant la pénétration. Il y a eu moins de pénétrations réussies chez les cultivars résistants, et la pénétration s'est déroulée plus lentement sur les cultivars résistants (Baker et Smith, 1978 ; Bird et Ride, 1981). La lignification a été proposée pour limiter l'infection dans les cultivars résistants et sensibles, mais d'autres facteurs ont ralenti le développement du champignon dans les lignées résistantes. Dans les lignées sensibles, les hyphes à croissance plus rapide peuvent échapper à la lignification des cellules hôtes. Quatre jours après l'inoculation de l'orge avec un isolat de S. nodorumLes hyphes de pénétration se développent à travers la cuticule et parfois dans les couches externes de cellulose des parois cellulaires épidermiques. Des papilles épaisses se sont déposées sous les hyphes de pénétration et les cellules n'ont pas été pénétrées (Keon et Hargreaves, 1984).
Infection par Septoria passerinii
Green et Dickson (1957) présentent une description détaillée du processus d'infection du S. passerinii sur l'orge. Le processus d'infection est similaire à celui de la S. tritici. Comme pour S. tritici, le temps nécessaire à la pénétration des feuilles est considérablement plus long que pour S. tritici. S. nodorum. Les tubes germinatifs se ramifient et se développent sur la surface de la feuille au hasard, mais parfois le long des dépressions entre les cellules épidermiques. La pénétration des feuilles se fait presque exclusivement par les stomates. Les hyphes de germination se gonflent et, si la pénétration échoue, les hyphes continuent à s'allonger. Il n'y a pas de pénétration 48 heures après le dépôt des spores. Après 72 heures, les tubes germinatifs s'épaississent au-dessus des stomates, se développent entre les cellules de garde et sur les faces des cellules accessoires et dans les cavités sous-matales. La pénétration directe entre les cellules épidermiques n'est que rarement observée.
La germination des spores et la pénétration de l'hôte sont les mêmes sur les cultivars résistants et sensibles. L'extension des hyphes à l'intérieur des feuilles est beaucoup moins importante sur les cultivars résistants et des papilles sont observées sur de nombreuses parois cellulaires, mais pas sur toutes. Les hyphes se développent sous l'épiderme d'un stomate à l'autre, mais ne pénètrent pas entre les cellules épidermiques. Le mésophylle est colonisé, mais aucun haustoria ne se forme. Après la nécrose des cellules du mésophylle, les cellules de l'épiderme s'effondrent. Le développement mycélien dans la feuille est peu abondant et généralement bloqué par les faisceaux vasculaires. Dans les feuilles plus jeunes, si la gaine vasculaire est moins développée, les hyphes passent entre le faisceau et l'épiderme. Les pycnides se forment dans les cavités sous-matales, principalement sur la face supérieure des feuilles (Green et Dickson, 1957).
Facteurs affectant la longévité des spores Parmi les pathogènes Stagonospora et Septoria des céréales, des informations définitives sur le processus d'infection n'ont été rapportées que pour le Stagonospora et le Septoria. S. nodorum, S. triticiet S. passerinii. Comme beaucoup d'autres pathogènes nécrotrophes, aucun de ces deux groupes de pathogènes ne provoque de réaction hypersensible. Une différence significative dans le processus d'infection entre les pathogènes Septoria et Stagonospora est que la germination et la pénétration des spores se déroulent beaucoup plus rapidement pour les Septoria et les Stagonospora. S. nodorum que pour S. tritici et S. passerinii. Cela a une influence significative sur l'épidémiologie des maladies.
Les agents pathogènes de la septoriose pénètrent dans la plante principalement par les stomates, tandis que les agents pathogènes de la S. nodorum pénètre à la fois directement et à travers les stomates. S. nodorum pénètre et tue rapidement les cellules de l'épiderme, mais S. tritici et S. passerinii ne tuent pas les cellules épidermiques avant que les hyphes ne se soient ramifiées à travers le mésophylle de la feuille et qu'une nécrose rapide ne commence. Les études histologiques de la croissance fongique après la pénétration de l'hôte correspondent aux données générées par les études épidémiologiques de la résistance de l'hôte. La résistance ralentit le taux de colonisation de l'hôte mais n'a pas d'effet appréciable sur le processus de développement des lésions.
Les mécanismes contrôlant la réponse de l'hôte, qu'ils soient liés aux enzymes et aux toxines ou à d'autres métabolites libérés par les pathogènes au cours de l'infection, ne sont pas encore clairs. Il existe peu d'informations sur l'infection par les ascospores. Le processus d'infection est probablement très similaire à celui des pycnidiospores. Les ascospores de Phaeosphaeria nodorum germent dans une large gamme de températures, et leurs tubes germinatifs pénètrent directement dans la feuille. Cependant, selon Rapilly et al. (1973), les ascospores, contrairement aux pycnidiospores, ne germent pas dans l'eau libre.
Modèle d'infection par Septoria spp.
Les infections par la septoriose sont possibles à des températures basses, alors que les températures inférieures à 7°C peuvent ne pas conduire à une infection dans les 2 jours. La température optimale de la maladie est atteinte dans une zone de 16 à 21°C. Les infections sont possibles pendant une période d'humidité relative élevée ou d'humidification des feuilles de 14 heures ou plus. Pour répondre à ces conditions, nous avons décidé de séparer les modèles pour les infections faibles, modérées et sévères. Les infections faibles peuvent être données s'il est possible pour le pathogène d'infecter le tissu de l'hôte. Cela signifie que les infections faibles peuvent avoir lieu si les températures sont minimales et si les périodes d'humidité des feuilles sont d'une durée critique. Une infection modérée a lieu dans des conditions où la plupart des essais d'infection donnent des résultats raisonnables et les infections sévères ont lieu dans des conditions où le pathogène a des conditions optimales pour l'infection.
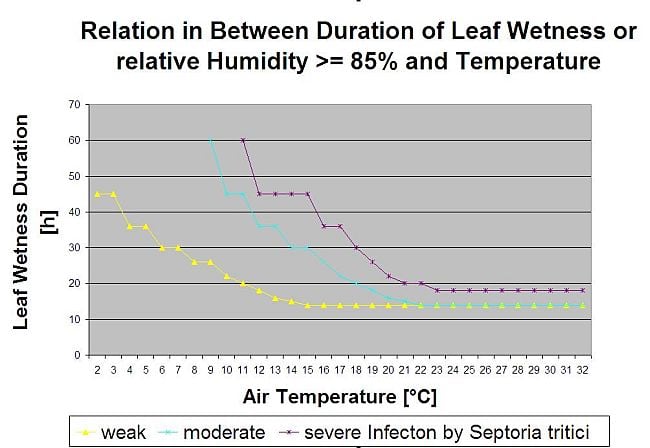

Début de l'infection après une pluie de 0,5 mm. Nous avons décidé de ne pas utiliser de modèle pour la formation des pycnides. Les conditions nécessaires à la formation des pycnides sont supposées être une période où l'humidité relative est supérieure à 85%. La durée de vie des pycnides est estimée à 24 heures. Dans tous les climats où Septoria tritici a une chance d'infecter, nous trouverons 2 heures remplissant ces conditions presque tous les jours autour du lever du soleil.
Évaluation de la gravité de l'infection
Pour pouvoir évaluer la pression d'infection de Septoria tritici entre le stade 10 (première feuille à travers le coléoptile) et le stade 32 (nœud deux au moins 2 cm au-dessus du nœud 1) et entre 32 et 51 (début de l'épiaison), nous devons évaluer la sévérité des infections en fonction des conditions climatiques. Cette évaluation se fait sur une échelle de 1 à 5. Une sévérité de 1 est donnée si la condition pour une faible infection est remplie et qu'il a plu moins de 5 mm, sinon la valeur de sévérité correspondante sera 2. Une gravité de 3 est attribuée si une infection modérée est remplie et qu'il a plu moins de 5 mm. S'il a plu plus de 5 mm au cours d'une infection modérée ou moins de 5 mm au cours d'une infection sévère, la gravité est de 4.
Une infection sévère avec plus de 5 mm de pluie correspond à une valeur de sévérité de 5.
Septoria tritici évaluation de la pression de la maladie
Le climat n'est qu'un des facteurs qui déterminent la pression de la maladie dans le champ. Les deux autres facteurs sont l'historique du champ et la sensibilité de la variété cultivée. Si nous pouvons accumuler les valeurs de sévérité de la maladie du stade 10 au stade 32 jusqu'à une valeur de 4, nous pouvons nous attendre à une faible pression de la maladie par le climat. Si cette valeur atteint 6, on peut s'attendre à une pression modérée de la maladie et si elle atteint 10, on peut s'attendre à une forte pression de la maladie de la part du climat. La connaissance de la sensibilité de la variété et de l'historique du champ nous amènera à pulvériser ou non en cas de pression faible ou modérée de la maladie dans cette situation. Une valeur cumulée de 10 peut de toute façon conduire à une pulvérisation au stade 32. La décision de pulvériser à un stade plus avancé dépend davantage du climat printanier. Si nous pouvons accumuler les valeurs de sévérité depuis le stade 10 jusqu'à une valeur de 6, nous pouvons nous attendre à une faible pression de la maladie. Si cette valeur atteint 10, on peut s'attendre à une pression modérée de la maladie et si cette valeur atteint 15, on peut s'attendre à une forte pression de la maladie en raison de la situation climatique.
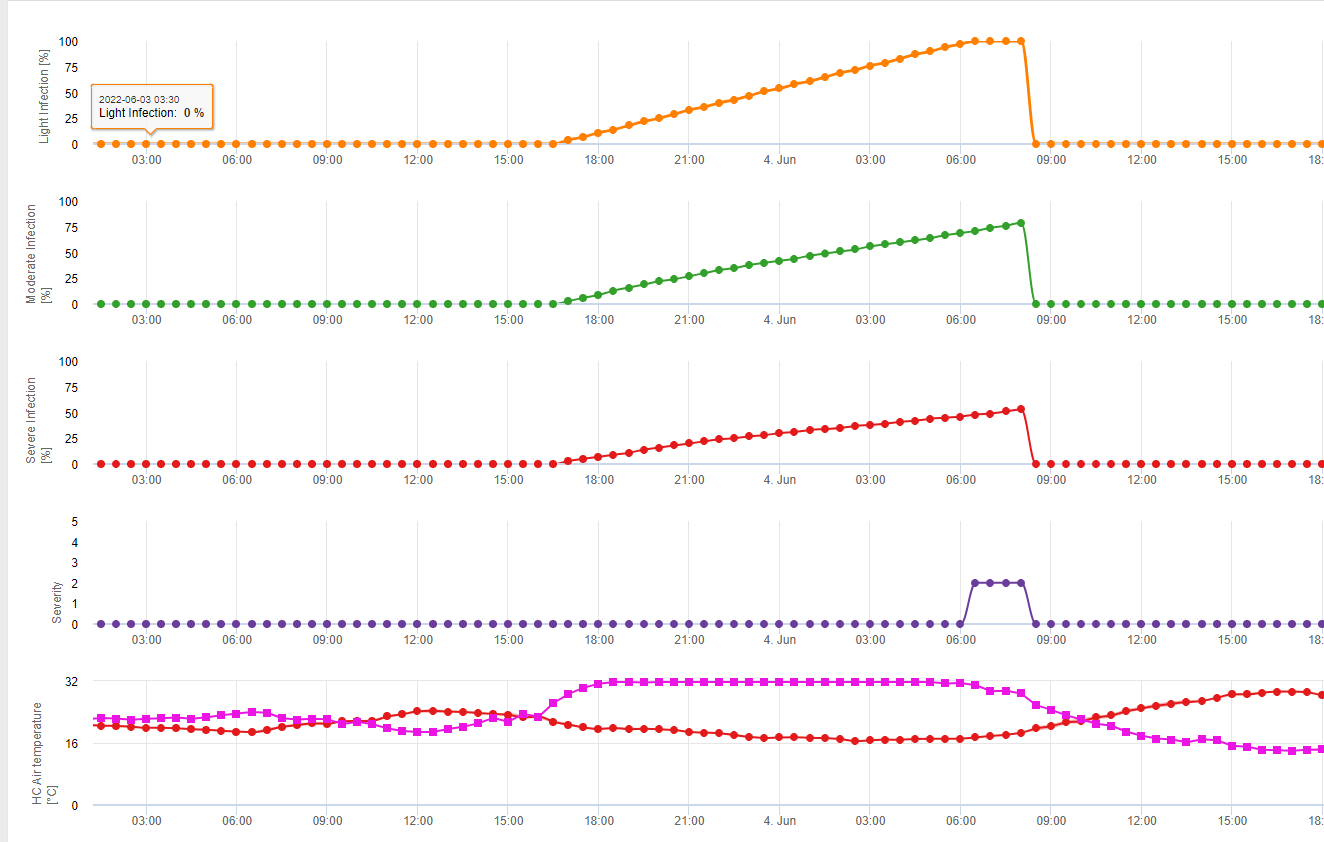
Dans FieldClimate, nous montrons le Septoria tritici Sévérité avec les trois différentes sévérités d'infection dans un graphique (voir ci-dessus). En raison des précipitations et des longues périodes d'humectation des feuilles, les conditions d'une infection sévère par le S. tritici ont été en plein champ les 14 et 16 mai. Les niveaux de gravité atteignent la valeur maximale de 5 le 14 mai, ce qui signifie que le risque d'infection est désormais élevé.
La biologie de l'infection de Stagonospora nodorum diffère dans une certaine mesure de celle de S. tritici, mais cette différence n'est pas suffisamment importante pour justifier un modèle distinct. C'est pourquoi nous suggérons d'utiliser ce modèle pour l'ensemble des maladies de Stagnospora et de Septoria dans les céréales, y compris S. passerinii. S. tritici et S. passerinii ont tendance à avoir besoin de périodes d'humectation plus longues que les S. nodorum. Dans les régions où l'anticyclone de S. nodorum Les infections classées comme faibles, avec une valeur de gravité de 2, doivent être traitées plus sérieusement que dans d'autres domaines.
Pour Septoria nodorum Un modèle de risque est présenté dans le document FieldClimate (voir ci-dessus). Un risque élevé a été déterminé le 17 juin et le 7 juillet (100%). En fonction du stade de la plante sensible à l'infection, des mesures de protection des plantes doivent être prises en compte si le risque atteint 80% (voir également les prévisions météorologiques, la protection des plantes). Si le risque est de 100% et qu'une infection a déjà été détectée, des mesures phytosanitaires systémiques (application curative) doivent être prises pour protéger la plante.