Biologie de Plasmopara viticola (Mildiou)
P. viticola est un parasite obligatoire. Cela signifie que les organes verts et frais de la vigne sont nécessaires à sa croissance. Pendant la période sans végétation, il persiste en formant des corps fructifères, appelés oospores. Les oospores des oomycètes peuvent survivre très longtemps dans le sol. C'est pourquoi on peut trouver le mildiou dans des endroits où les infections ne sont pas possibles chaque année. Au printemps, lorsque le sol est suffisamment humide et chaud, les oospores forment des macrosporanges qui peuvent libérer jusqu'à 200 zoospores dans l'eau libre. Les zoospores se déplacent vers les feuilles et les grappes sous l'effet du vent dans les gouttelettes d'eau. Elles possèdent deux flagelles et se déplacent dans un film d'eau sur la face inférieure des feuilles ou des grappes et des jeunes baies pour trouver un stoma qui leur permettra de pénétrer dans les tissus végétaux. Elles pénètrent et germent dans le stomate, dans lequel elles transfèrent tout leur plasma en moins d'une heure. Dans les études microscopiques, la découverte des stomates, l'enkystement et la germination dans les stomates sont terminés en 90 minutes.
P. viticola se développe dans l'espace intercellulaire et se nourrit d'haustorias pénétrant dans les cellules épidermiques et parenchymateuses. En fonction de la température et de l'humidité relative, il développe suffisamment de croissance intercellulaire avec suffisamment d'haustorias pour former un corps sous-matellaire qui remplit toute la zone sous-matellaire et qui soulève le tissu épidermique du tissu parenchymateux. Cela conduit au symptôme visible de la tache d'huile.
Les oomycètes sont sporulant en l'absence de lumière lorsque l'humidité relative est très élevé. Dans le cadre de P. viticola il n'y a pas de sporulation si les températures sont inférieures à 12°C et l'humidité relative inférieure à 95%. Les sporangiaphores sont formés par les vésicules de substomatelle et sortent du stomate. Les sporanges fraîchement formés sont collants et ne peuvent être retirés des sporangiaphores qu'avec de l'eau. Lors de la diminution de l'humidité relative, les sporanges deviennent secs et peuvent être enlevés par le vent.
Les sporanges libèrent jusqu'à 20 zoospores dans l'eau libre. Ces zoospores doivent être distribués par le vent dans des gouttelettes d'eau pour atteindre les feuilles fraîches, ou les sporanges peuvent être distribués par la pluie ou le vent lui-même. Le processus d'infection primaire et secondaire est le même.
En raison de la grande importance du stade sexuel pour l'hibernation du pathogène, nous pouvons supposer que les types d'accouplement conviennent à tous les vignobles où le mildiou de la vigne est présent. La formation des zoospores a lieu sur les feuilles plus âgées pendant l'été et au début de l'automne.
La primo-infection en détail
Les infections provenant des oospores sont appelées infections primaires. Ce terme est trompeur car plusieurs infections primaires peuvent se produire au début de l'été. Dans les événements épidémiologiques, l'infection primaire ne joue pas un rôle important s'il y a suffisamment de taches d'huile dans les vignobles et si le potentiel d'infection des spores d'été (sporanges) dépasse celui des oospores.
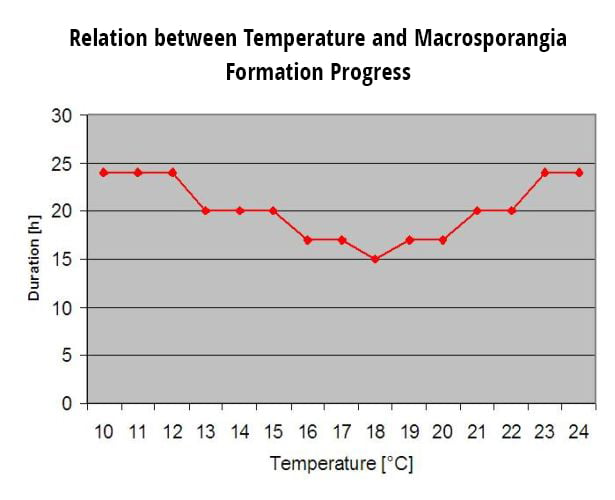
Les oospores hivernantes se transforment en macrosporanges lorsque l'humidité relative est suffisante pendant environ 24 heures. Les macrosporanges libèrent leurs zoospores dans l'eau libre et les fortes pluies (comme lors d'un orage). Les spores sont transportées vers les feuilles de la vigne et/ou les pousses vertes. Il s'agit donc d'infections primaires, ont besoin d'une pluie plus longue La première pluie provoque la saturation en eau des feuilles tombées au sol et les fortes pluies des jours suivants provoquent la libération des zoospores. La première pluie provoque la saturation en eau des feuilles tombées au sol et les fortes pluies des jours suivants provoquent la libération par les macrosporanges de leurs zoospores, qui atteignent les feuilles ou les sarments de la vigne. Pour que l'infection réussisse, une période d'humidité des feuilles suffisamment longue est nécessaire pour permettre aux zoospores d'atteindre les stomates des feuilles ou des pousses et de les infecter (l'illustration ci-dessus montre le cycle de développement du mildiou de la vigne (Plasmopara viticola).
Le modèle de l'infection primaire du mildiou vérifie d'abord si le temps est propice au développement des macrosporanges. C'est le cas tant que les feuilles sont humides ou que l'humidité relative après la pluie ne tombe pas en dessous de 70%. En fonction de la température, les macrosporanges peuvent se développer en 16 à 24 heures. Si des macrosporanges matures sont disponibles, cela sera indiqué graphiquement dans l'affichage de l'infection primaire du mildiou. Si des macrosporanges sont présents, une forte pluie peut disséminer leurs zoospores. Une pluie continue de 5 mm est interprétée comme une forte pluie et les zoospores sont disséminées, une infection primaire est alors possible si l'humidité des feuilles a duré suffisamment longtemps.
 Infections secondaires
Infections secondaires
 Infections secondaires
Infections secondairesInfections secondaires de Plasmopara viticola ne sont possibles que si des taches d'huile déjà mûres existent dans votre vignoble ou à proximité. Les taches d'huile sont matures lorsqu'elles sont capables de sporuler (produire des sporanges). Les sporanges ne sont produits que par nuit. La lumière du soleil inhibe la sporulation. Les sporanges sont produits s'il y a plus chaud que 12°C et le humidité relative est très élevé. Le taux de production de sporanges augmente avec la température jusqu'à 24°C. La température optimale pour la sporulation sur les cépages européens (Vitis vinifera) est d'environ 24°C. Si les températures dépassent 29°C, aucune sporulation ne peut avoir lieu. Dans notre modèle, nous vérifions si humidité supérieure à 95% se produit pendant la nuit. Si cette condition dure pendant une température horaire cumulée de plus de 50°C, la sporulation est terminée et de nouveaux sporanges de Plasmopara existent dans le vignoble. Par exemple, les heures à 50°C correspondent à 4 heures à 13°C ou à 3 heures à 17°C.
Sporanges de Plasmopara viticola ont une durée de vie limitée. Plus il fait chaud et plus l'air est sec, plus ils meurent rapidement (dans ng.fieldclimate.com, nous remettons à 0 lorsque l'humidité relative est inférieure à 50%). Ils meurent définitivement au cours de la prochaine période de rosée ou d'humidité des feuilles, qui est trop courte pour l'infection. L'aptitude des sporanges plus anciens est donc très limitée. Notre modèle suppose que la durée de vie des sporanges est limitée à un jour.
Pour provoquer de nouvelles infections, les sporanges doivent être disséminés. Il y a deux façons de distribuer les spores : S'il pleut immédiatement après la formation des sporanges, ceux-ci se répandent avec les éclaboussures de la pluie. Si les feuilles de la vigne restent humides assez longtemps, un niveau élevé de spores se propage. Plasmopara viticola Les infections ont lieu. Si le lendemain matin commence sans pluie et avec une humidité décroissante, les sporanges desséchés se détachent de leurs branches. Le moindre mouvement d'air les transporte vers les feuilles saines. S'il ne pleut pas rapidement, elles mourront.
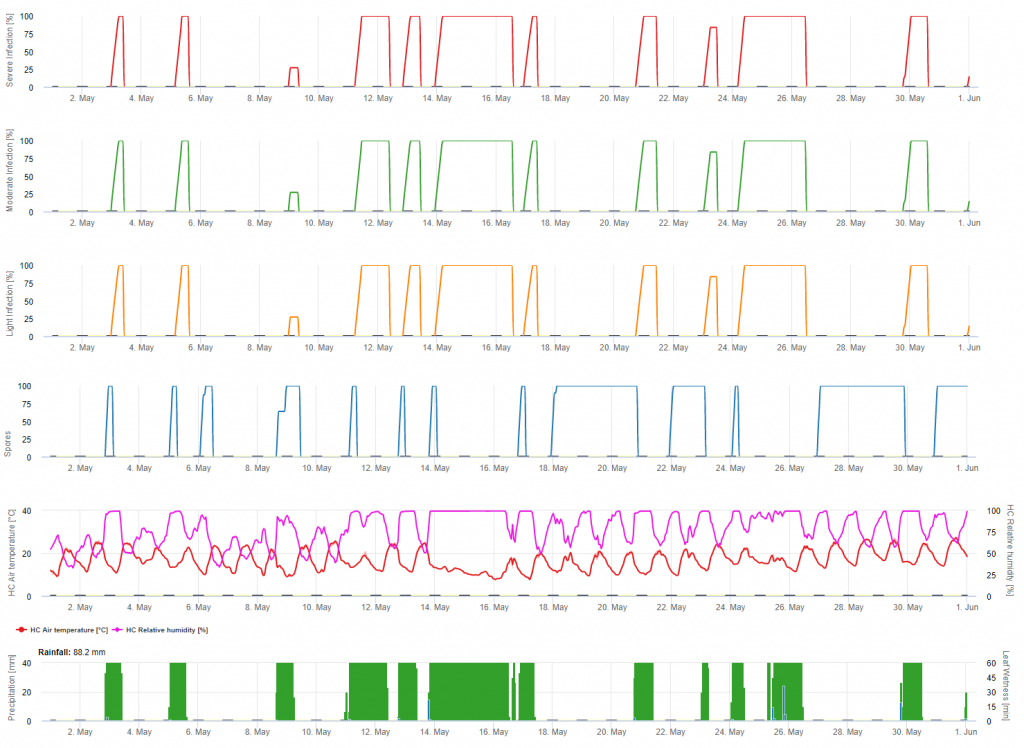
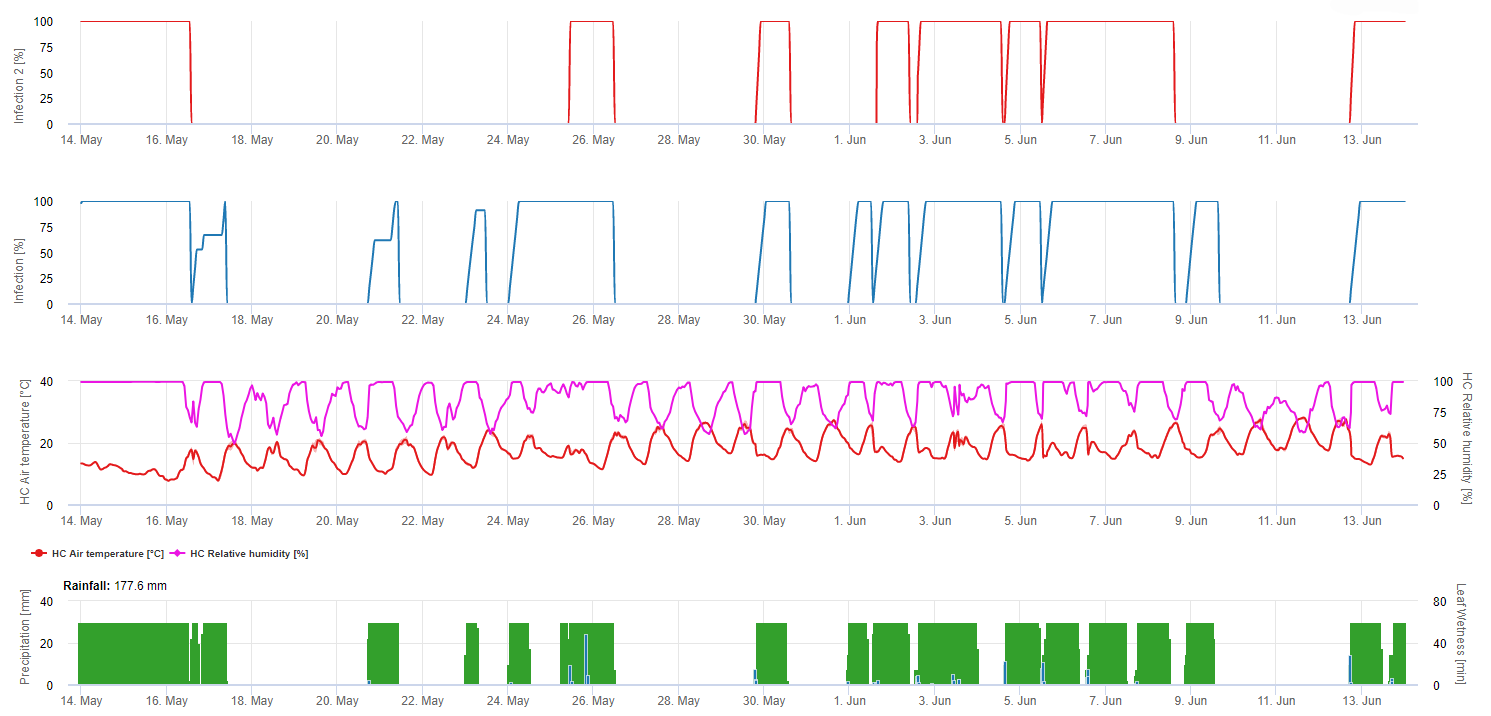
Mildiou Infection primaire
Les infections sont représentées après le développement des macrospores (orange, cinquième graphique) dans trois classes de gravité différentes (faible : orange, modérée : verte, sévère : rouge) en fonction de la quantité de pluie et donc de la distribution des spores dans le vignoble. Si l'infection par 100% est déterminée, les conditions optimales ont été réunies pour pénétrer dans le tissu végétal (protection curative de la plante) et la courbe d'incubation (premier graphique) commence à augmenter. Si l'incubation se termine (100%) et qu'aucune protection phytosanitaire n'a été prise en compte, les symptômes (taches d'huile) auraient dû être vérifiés sur le terrain.
1. La courbe de progression de l'infection et, après une infection complète, la période d'incubation qui en découle (temps écoulé entre l'infection et les symptômes visibles sur le terrain).
2. Une courbe de progression de l'infection est déterminée lorsque des macrosporanges ou des sporanges sont présents et que les feuilles sont humides. Elle augmente en fonction de l'humidité des feuilles et de la température. Si la courbe de progression de l'infection atteint 100%, on peut considérer que l'infection est terminée. La courbe de progression de l'incubation correspondant à l'infection spécifique commence à être calculée au début de l'infection. Si une infection ne peut pas être achevée, la courbe d'incubation sera également arrêtée.
3. Les conditions météorologiques sont affichées sur la même page que les résultats du modèle de maladie. Vous pouvez donc vérifier d'un seul coup d'œil la température, l'humidité relative, la pluie et l'humidité des feuilles pendant l'infection.
4. Le modèle indique les dates d'infection pour les infections primaires et secondaires du mildiou de la vigne. Il s'agit là de l'information la plus importante. Les infections qui n'ont pas encore été couvertes par des fongicides préventifs ou curatifs peuvent être désastreuses pour le rendement/la qualité. Des informations supplémentaires sur la gravité de l'infection (faible/moyenne/sévère) pourraient être utiles pour décider de la stratégie de protection des plantes en fonction de l'historique du vignoble (quantité d'inoculum), de la variété et de l'expérience des années précédentes.
Stratégies de protection des plantes :
- Les fongicides prophylactiques et systémiques ou curatifs sont largement utilisés pour lutter contre le mildiou. Les produits chimiques prophylactiques sont appliqués avant, mais aussi près que possible d'un épisode d'infection dans la période de plus grande sensibilité de l'hôte, entre une longueur de pousse de 10 cm et des baies de la taille d'un pois.
- Les produits chimiques prophylactiques peuvent également être utilisés en pulvérisant le plus tôt possible avant un événement météorologique prévu de plus de 2 mm de pluie (pour le cercle primaire).
- Un fongicide curatif doit être appliqué dès que possible après un épisode d'infection (infection par 100%) et avant l'apparition de taches d'huile. Veillez à prévenir le développement de la résistance aux fongicides.
- La surveillance de la présence du mildiou (taches d'huile) dans le vignoble doit être effectuée régulièrement et prendre en compte au moins 200 pieds de vigne. On considère qu'il y a un risque si l'on observe plus de deux taches d'huile pour 50 pieds de vigne.
Le modèle se base sur les publications de MUELLER-THURGAU, ARENS, MUELLER et SLEUMER, BLAESER, HERZOG, GEHMANN et de nombreux autres travailleurs liés au réseau de recherche sur l'épidémiologie du mildiou et de l'oïdium de la vigne depuis le début du dix-neuvième siècle.
Biologie de Uncinula necator
L'oïdium de la vigne est causé par le champignon Uncinula necator. Il s'agit d'un parasite obligatoire, ce qui signifie qu'il a besoin de tissus verts pour se nourrir. U. necator a fait son entrée en Europe au XIXe siècle et s'est répandue dans tous les pays viticoles jusqu'à la fin de ce siècle. Il a des effets très importants sur le rendement et peut entraîner la défoliation des vignes s'il n'y a pas de contrôle chimique. Toutes les variétés de vignes européennes sont sensibles à ce pathogène. La sélection de la résistance à ce pathogène est assez récente et les résultats nécessiteront des années de travail.
U. necator doit surmonter la période sans végétation dans la plupart des zones de culture de la vigne. Pour ce faire, le pathogène utilise deux mécanismes : a) survivre dans ses organes de fructification, les cleistothèques, ou b) survivre aux hivers en étant encapsulé dans les bourgeons dormants où il trouve des tissus vivants pendant tout l'hiver. La présence d'un seul ou des deux modes d'hibernation dans un vignoble dépend de l'historique de la maladie dans ce vignoble. Les bourgeons ne peuvent être infectés que tant qu'ils ne sont pas fermés et entourés d'un anneau de tissu sénescent. Ce n'est le cas que pendant quelques semaines après la formation des bourgeons. En situation de viticulture ordinaire, les vignes sont taillées en branches de moins de 14 bourgeons ou en coursons. Les bourgeons restants ne peuvent être infectés que jusqu'à la floraison. D'un autre côté, on peut trouver des sarments du stade BBCH 15 à 19. Ces pousses sont des pousses partiellement ou totalement couvertes de mildiou et formées à partir de bourgeons infectés latents. Ces pousses sont une source d'inoculum précoce et très puissante. Elles ne sont généralement pas réparties à l'intérieur d'un vignoble. Cela conduit à des taches avec une incidence précoce et élevée de la maladie à l'intérieur des vignobles et à des taches avec une maladie faible ou modérée au début de la saison. Les pousses en drapeau ne représentent qu'un seul type d'accouplement en raison de leur histoire végétative. Cela peut conduire à des vignobles où l'on ne trouve que peu ou pas de cleistothèques en raison de la dominance d'un seul type d'accouplement.
Lorsque les cleistothèces sont le seul inoculum, on constate une hibernation continue des cleistothèces qui ne se transforment en pousses drapeaux que les années où la maladie se développe très tôt. La formation des cleistothèces est bien visible en raison de la coloration jaune du tissu sexuel combiné. On peut la trouver dans les vignobles non pulvérisés à partir du milieu de l'été. Dans les vignobles pulvérisés, l'incidence de la maladie est plus longue à se développer, ce qui permet aux types d'accouplement de se rencontrer. La propagation de la Uncinula necator pendant l'été est principalement influencée par la température.
Les températures optimales pour l'oïdium se situent entre 21°C et 32°C. Plus le nombre d'heures quotidiennes dans cette plage de températures est élevé, plus le risque d'oïdium est important. La formation des conidies, l'infection et la vitesse d'incubation nécessitent des plages de températures similaires. Étant donné que U. necator ne nécessite pas d'eau libre pour l'infection et pas d'humidité relative élevée pour la formation des conidies, il n'y a pas d'impact utilisable du paramètre d'humidité directe sur le pathogène.
La présence d'humidité libre interagit avec U. necator par l'intermédiaire de son hyperparasite commun Ampelomyces quisqualis. Cet hyperparasite fongique se développe à l'intérieur des hyphes du champignon de l'oïdium. Il transforme les conidies et les cleistothèces en ses propres pycnides. Il a besoin d'humidité libre ou d'une humidité relative très élevée pour infecter et se développer avec de fines hyphes à croissance rapide sur les feuilles pour trouver des lésions d'oïdium. Fortement infecté U. necator Les lésions prennent une couleur marron très foncée. En période de pluie, elles sont recouvertes d'un exsudat de spores blanches provenant des pycnides de l'hyper parasite. Ampelomyces quisqualis est également disponible en tant que produit commercial pour lutter contre l'oïdium.
Travaux pratiques
Nous avons mis en œuvre différents modèles concernant le processus d'infection dans FieldClimate.
a) le champignon hiverne principalement sous la forme d'un mycélium. Au printemps, le mycélium commence à se développer et construit des conidiophores avec des conidies (il n'y a donc pas d'infection primaire).
b) si le champignon se développe en automne sur les baies, la forme sexuée peut être présente (construction de cleistothèces) et donc, au printemps, la libération d'ascospores peut conduire à une infection primaire (utiliser : modèle POWDERY MILDEW/ASCOSPORE INFECTION).
Modèle de risque californien (par Thomas Gubler)
Le modèle d'évaluation du risque de l'oïdium de l'UC Davis (modèle Gubler-Thomas) est basé sur des études épidémiologiques en laboratoire et sur le terrain de l'oïdium de la vigne en Californie.
Indice d'évaluation des risques :
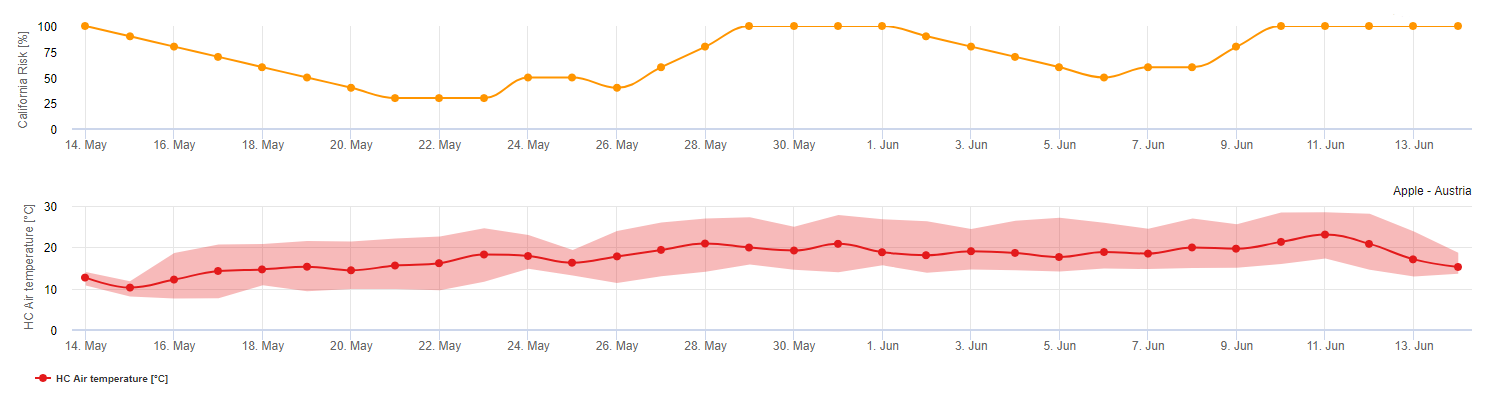
Après la libération et la germination des ascospores (utiliser pour ce modèle ASCOSPORE INFECTION), le développement et la reproduction de l'oïdium sont principalement influencés par les températures ambiantes. Le modèle UC Davis évalue les températures ambiantes et le risque de développement de l'oïdium à l'aide d'un indice de 0 à 100 points.
Pour déclencher l'indice d'évaluation du risque, après le débourrement et après l'apparition de la maladie, il faut trois jours consécutifs avec un minimum de six heures consécutives de températures comprises entre 21 et 30 °C pour que l'épidémie d'oïdium soit déclenchée. Tout au long du calcul de l'indice, pour chaque journée avec 6 heures consécutives ou plus entre 21 et 32 °C, l'indice augmente de 20 points. Le modèle fluctue ensuite entre 0 et 100 points d'indice en fonction de la durée des températures journalières. L'indice gagne 20 points pour chaque jour qui satisfait à l'exigence de six heures consécutives de températures comprises entre 21 et 32 °C, et il perd 10 points pour chaque jour qui ne satisfait pas à l'exigence de six heures, ou si, à tout moment de la journée, la température dépasse 32 °C ou est inférieure à 21 °C.
Interprétation de l'indice :
Un indice faible de 0 à 30 indique que le pathogène ne se reproduit pas. Les producteurs peuvent envisager de reporter les applications de fongicides pendant les périodes prolongées où l'indice est faible. Un indice de 40-50 est considéré comme modéré et implique un taux de reproduction de l'oïdium d'environ 15 jours. Des valeurs d'indice de 60-100 indiquent que le pathogène se reproduit rapidement (tous les 5 jours) et que le risque d'épidémie est élevé.
Modèle de risque Pessl Instruments
La dispersion des Uncinula necator pendant l'été est influencée principalement par température. L'antagoniste de U. necator appelé Ampelomyces quisqualis est principalement favorisé par l'humidité libre. Les températures optimales pour l'oïdium se situent entre 21°C et 32°C. Plus le nombre d'heures quotidiennes pendant lesquelles la température se situe dans cette fourchette est élevé, plus le risque d'apparition de l'oïdium est important (voir modèle Gubler). Les journées avec plus de 6 heures de température dans cette fourchette augmentent le risque d'environ 20 points.. Les jours où la température ne se situe pas dans cette fourchette optimale réduisent le risque d'environ 10 points. Il s'agit des jours où la température est inférieure à 21°C toute la journée ou des jours où la température dépasse 32°C pendant 6 heures. En revanche, les jours où la température est inférieure à 21°C toute la journée ou dépasse 32°C pendant 6 heures. humidité de la feuille longue Les périodes de sécheresse conduisent à l'établissement d'un champignon antagoniste. A. quisqualis. Ces mesures conduiront à une diminution de U. necator.
Pour ce modèle, nous recommandons de suivre ces règles : Si le risque d'oïdium est inférieur à 20 points, l'intervalle de pulvérisation peut être prolongé. Entre 20 et 60 points, l'intervalle de pulvérisation normal est valable. Si le risque est supérieur à 60 points, il convient de réduire l'intervalle de pulvérisation.
Le risque commence : avec 3 jours avec un nombre d'heures égal ou supérieur à 6 : 21°C <= Température 60 Points
Le risque augmente : Avec chaque jour ayant une durée égale ou supérieure à 6 heures : 21°C <= Température +20 Points
Le risque diminue : Chaque jour où la température n'atteint pas 21°C ou chaque jour avec 6 heures de : 32°C -10 Points
Modèle de troisième risque : Inoculum primaire d'ascospores
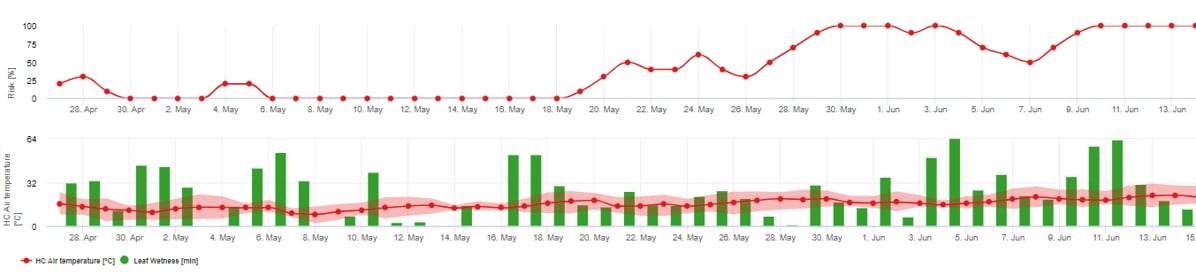
Nous recommandons d'utiliser ce modèle lorsque les ascospores sont formées pendant les périodes froides de l'hiver (pour éviter l'infection par le mycélium ("Flag shoots")).
L'infection des ascospores prédit la libération des ascospores et les infections initiales, qui sont basées sur les températures moyennes pendant les périodes d'humidité prolongée des feuilles.
Pour le calcul en fieldclimate.com environ 2,5 mm de précipitations sont nécessaires pour libérer les ascospores, suivies de au moins 8 à 12 heures d'humidité des feuilles après la libération des ascospores à des températures comprises entre 10 -15°C pour l'infection. Une fois que l'infection a eu lieu, le modèle passe à une phase d'évaluation du risque de maladie (utiliser le modèle Gubler/Risk) et se base entièrement sur les effets de la température sur le taux de reproduction de l'agent pathogène.
Utilisation pratique
Dans ce modèle, nous affichons l'infection par Uncinula necator dans une ligne comprise entre 0 et 100%.
Nous présentons deux modèles différents dans FieldClimate:
1. Les infections d'ascospores possibles selon les modèles proposés par David Gadoury et Doug Gubler sont affichées dans le graphique d'infection d'ascospores. Si la courbe de progression du modèle d'infection atteint 100%, il est possible qu'il y ait une infection par ascospores. 3 à 5 jours après cet événement, les premiers symptômes de l'oïdium peuvent être visibles sur les feuilles proches de l'ancien cordon.
2. Le risque d'oïdium selon le modèle californien et le modèle pour les zones à climat modéré est représenté dans le graphique ci-dessus. Les périodes à risque doivent être couvertes par des applications de fongicides. Dans les périodes où le risque est supérieur à 60 points, il est recommandé d'effectuer des pulvérisations plus fréquentes, en particulier autour et peu après la floraison.
La pourriture noire de la vigne touche de nombreux viticulteurs aux États-Unis et dans la plupart des autres régions chaudes et arides du monde. Pour gérer la maladie, il est important de comprendre son cycle de vie et les conditions environnementales. Une fois que l'infection a eu lieu, différentes méthodes sont disponibles pour contrôler la maladie.
Biologie
L'agent pathogène de la pourriture noire de la vigne passe l'hiver dans de nombreuses parties de la vigne et peut également passer l'hiver sur le sol, en particulier dans les momies. En outre, le pathogène peut passer l'hiver pendant au moins deux ans dans les lésions des pousses infectées qui sont conservées sous forme de sarments ou d'éperons.
Une fois, les pluies peuvent libérer les spores hivernantes (ascospores) qui se forment dans les momies sur le sol et peuvent être dispersées par le vent. Certaines momies sur le sol peuvent avoir une décharge importante d'ascospores qui commence environ 2 à 3 semaines après le débourrement des bourgeons et arrive à maturité 1 à 2 semaines après le début de la floraison. Un deuxième type de spore (conidies) peut également se former dans les lésions de la canne ou dans les momies qui sont restées dans le "treillis, et celles-ci sont dispersées sur de courtes distances (de quelques centimètres à quelques pieds) par les gouttes de pluie". L'infection se produit lorsque l'un ou l'autre des types de spores se pose sur le tissu vert du raisin et que le tissu reste humide pendant une "durée suffisante, qui dépend de la température". La période pendant laquelle ces spores hivernantes peuvent provoquer une infection dépend de la source. S'il y a une source d'infection importante, l'infection s'installera rapidement. En présence d'humidité, ces ascospores germent lentement, en 36 à 48 heures, mais finissent par pénétrer dans les jeunes feuilles et les pédicelles des fruits. Les infections deviennent visibles après 8 à 25 jours. Lorsque le temps est humide, les ascospores sont produites et libérées pendant tout le printemps et l'été, ce qui permet une infection primaire continue. (Wilcox, Wayne F. "Black rot Guignardia bidwellii." Fiche d'identification de la maladie n° 102GFSG-D4. 2003. Cornell. 24 oct. 2010).
Le champignon de la pourriture noire a donc besoin d'un temps chaud pour se développer de manière optimale ; le temps frais ralentit sa croissance. Une période de 2 à 3 jours de pluie, de bruine ou de brouillard est également nécessaire pour l'infection". (Ries, Stephen M. "IPM : Reports on Plant Diseases : Black Rot of Grape". Integrated Pest Management at the University of Illinois. Déc. 1999. 24 oct. 2010). Les gouttes de pluie transmettent ces spores en les déplaçant vers différentes parties de la plante, en particulier les jeunes feuilles sensibles. En présence d'eau, les conidies germent en 10 à 15 heures et pénètrent dans les jeunes tissus. De nouvelles infections de pourriture noire se poursuivent jusqu'à la fin du printemps et en été pendant les périodes prolongées de temps chaud et pluvieux. Les conidies sont capables de germer et de provoquer une infection plusieurs mois après leur formation. Au mois d'août, les pycnides se transforment en un stade d'hivernage qui, à son tour, donne naissance à des pseudothèces à l'intérieur desquels les spores de printemps (ascospores) sont produites. Des recherches ont montré que les ascospores sont une source importante d'infections primaires au printemps. Au printemps, par temps humide, les "pycnides" des tissus infectés absorbent l'eau et les conidies sont expulsées. Les conidies sont projetées au hasard par la pluie et peuvent infecter n'importe quel jeune tissu en moins de 12 heures à des températures comprises entre 60 et 90 degrés. (Ellis, Michael A. "Fact sheet Agricultural and Natural Resources : Grape Black Rot". Département de pathologie végétale. Université de l'Ohio State Extension. 2008).
Utilisation pratique
Pour notre modèle, nous nous basons sur la littérature publiée par Spotts (Spotts. R. A. 1977. Effet de la durée d'humectation des feuilles et de la température sur l'infectivité du Guignardia). En discutant avec Daniel Molitor, nous avons modifié le modèle original de Spotts pour en faire un modèle séparant les infections légères, modérées et sévères, semblable à la conception des modèles de tavelure du pommier. (Daniel Molitor : Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Dissertation Justus-Liebig-Universität Gießen, 2009).
Une infection répondant aux "critères de Spotts" est considérée comme une infection légère. Les infections répondant aux "critères de Spotts" par 150% sont considérées comme modérées et les infections répondant aux "critères de Spotts" par 200% sont considérées comme sévères.
Le modèle indique les dates d'infection pour les infections légères, modérées et sévères. Lorsque 100% d'une infection est remplie, les conditions pour l'émission de spores, la germination et l'infection du tissu végétal sont données. La gravité de l'infection dépend de la période de température optimale et de la période d'humidité du champignon. En fonction de la pression d'infection, les infections doivent être couvertes de manière préventive ou une pulvérisation curative doit être appliquée peu de temps après l'infection. Dans le climat semi-aride modéré de la Moselle ou de la plupart des régions viticoles autrichiennes, il n'est pas recommandé de procéder à une pulvérisation après avoir constaté une infection légère.
Lobesia botrana (Totricidae, Olethreutinae) a été décrite pour la première fois en Autriche, mais elle est originaire du sud de l'Italie et a immigré à travers l'Europe, l'Afrique du Nord et de l'Ouest, le Moyen-Orient et la Russie orientale. Il a été plus récemment introduit au Japon, et en 2008 il a été signalé pour la première fois au Chili.
Hôtes
Raisin (Vitis vinifera) et le laurier-sauce (Daphne gnidium) sont les hôtes préférés, mais il a également été signalé sur des mûres (Rubus fruticosus), la groseille à maquereau (Ribes sp.), groseilles noires et rouges (Ribes nigurm), olive (Olea europaea), cerise (Prunus avium), élaguer (Prunus domestica), le kaki (Diospyrus kakis), le kiwi (Actinidia chinensis), la grenade (Punica granatum), l'œillet (Dianthus spp.), et un certain nombre d'autres hôtes sauvages.
Dégâts
En mai et juin, les larves de la première génération tissent leur toile et se nourrissent des grappes de fleurs. Les larves de la deuxième génération (juillet-août) se nourrissent des baies vertes. Les jeunes larves pénètrent dans les baies et les creusent, laissant la peau et les graines. Les larves de troisième génération (août-septembre) causent les dégâts les plus importants en s'enroulant et en se nourrissant à l'intérieur des baies et des grappes, qui deviennent contaminées par les excréments. En outre, les dommages causés aux baies par l'alimentation favorisent l'infection par Botrytis et d'autres champignons secondaires tels que Aspergillus, Alternaria, Rhizopus, Cladosporium, et Penicillium.
Biologie et cycle de vie
Le papillon adulte mesure environ 6 à 8 mm de long, pour une envergure de 11 à 13 mm. La femelle est légèrement plus grande. Les mâles et les femelles ont des ailes similaires à motifs en mosaïque. La première paire d'ailes (ailes antérieures) est de couleur crème fauve, tachetée de gris-bleu, de brun et de noir. La deuxième paire d'ailes est grise avec une bordure frangée. Au repos, les ailes sont maintenues en forme de cloche sur l'abdomen. Les femelles pondent leurs œufs séparément. Les œufs sont elliptiques et plats, d'un diamètre d'environ 0,6 à 0,8 mm. Ces œufs en forme de lentilles sont visibles et d'un blanc crème irisé, virant au jaune lorsque l'embryon se développe, puis au noir lorsque la tête de la larve en développement se forme. Il existe cinq stades larvaires. Le premier stade mesure environ 1 mm et le dernier environ 12-15 mm. Les larves ont une tête noire, puis elles deviennent brun jaunâtre et leur corps prend la couleur de leur alimentation (contenu des intestins). Le cinquième et dernier stade larvaire tisse un cocon de silicone grisâtre pour se nymphoser. La chrysalide mesure environ 4 à 9 mm de long.
Dans notre zone climatique, la tordeuse de la vigne a deux à trois générations par an. Quatre générations sont signalées dans les régions plus chaudes d'Espagne, de Grèce et d'Égypte. Le stade nymphal passe l'hiver à l'intérieur des cocons de silicium (diapause) sous l'écorce ou dans les fissures du sol ou dans des endroits cachés et protégés. Lorsque la température de l'air augmente au-dessus de 10°C pendant environ 10 à 12 jours, les adultes éclosent. Les premiers mâles sont trouvés et après une semaine, les femelles éclosent. Le premier vol des mâles peut donc commencer dès le débourrement et se poursuivre pendant 4 à 5 semaines. Les activités de vol ont lieu lorsque le soleil se lève et que les températures avoisinent les 12°C. Pendant le vol, les insectes s'accouplent et la majorité des femelles ne s'accouplent qu'une seule fois. Les activités de ponte commencent peu après l'accouplement (environ un à deux jours). Les femelles déposent les œufs individuellement sur les fleurs ou à proximité. Une femelle pond environ 100 œufs et la durée de vie adulte est de 1 à 3 semaines en fonction des conditions climatiques.
Au printemps, l'éclosion des œufs prend 10 jours ou plus, tandis qu'en été, les œufs éclosent en 3 ou 4 jours, en fonction des conditions de température. Les larves de la première génération s'enroulent sur les fleurs et s'en nourrissent, elles peuvent pénétrer dans le pédoncule et provoquer le dessèchement de la grappe. Le développement larvaire dure de 20 à 30 jours selon les conditions climatiques. Elles se nymphosent ensuite sous l'écorce, dans le sol ou sur la fleur et les adultes émergent à nouveau 6 à 14 jours plus tard.
Les femelles des deuxième et troisième vols pondent des œufs individuellement, directement sur les baies ombragées. Peu après l'émergence de la larve, celle-ci pénètre dans la baie et la creuse pour se nourrir. Une seule grappe peut être infestée par plusieurs larves. Les toiles, les excréments et les infections fongiques peuvent entraîner une contamination étendue de la grappe.
Le seuil inférieur de développement est d'environ 10 °C et le seuil supérieur de développement est de 30 °C. Les conditions optimales de développement se situent entre 26 et 29°C et 40 à 70% d'humidité. Si la durée du jour diminue et qu'il fait plus froid, le stade de la diapause commence. Les stades larvaires meurent lorsque les températures tombent en dessous de 8°C, mais la nymphe en diapause peut résister même aux hivers froids de l'Europe du Nord.
La première génération est plus courte que les générations d'été. Les œufs éclosent au bout d'environ 66 degrés-jours Celsius (DDC). Les larves qui se nourrissent de grappes de fleurs se développent plus rapidement que celles qui se nourrissent de baies de raisin plus tard dans la saison, ce qui influe sur le temps de génération. Les pupes non diapausantes ont besoin d'environ 130 DJ pour se développer. Les femelles adultes peuvent pondre des œufs environ 61 jours après l'émergence.
Les estimations de DD pour une génération varient considérablement dans la littérature, de 427 DDC à 577 DDC pour la première génération à 482 DDC à 577 DDC pour les générations suivantes.
Suivi et gestion
Les mâles sont attirés par les phéromones, qui sont utilisées pour surveiller les vols des mâles. Avant le débourrement, placez des pièges avec des L. botrana des leurres placés en hauteur (phéromone iScout). Placer au moins un piège par tranche de 30 acres ou par bloc de vignoble s'il est plus petit. Changer les appâts selon les recommandations du fabricant. Vérifier les pièges chaque semaine, en notant le nombre de papillons capturés et en retirant les papillons piégés du fond du piège collant. Tracer les captures hebdomadaires pour déterminer le début et le pic des vols de mâles dans chaque génération. Poursuivre la surveillance à l'aide de pièges jusqu'à l'apogée du troisième vol.
Les applications d'insecticides doivent être programmées pour l'émergence des larves, c'est pourquoi la surveillance de la ponte et la détermination de l'éclosion des œufs sont essentielles à la gestion de ce ravageur. Pour la première génération, la ponte doit être surveillée du pic jusqu'à la fin du vol. Recherchez les œufs sur le pédoncule de 100 grappes, en sélectionnant une grappe par vigne. Noter le stade de la majorité des œufs trouvés. Les œufs sont blancs lorsqu'ils viennent d'être pondus, ils deviennent jaunes puis noirs lorsque les larves sont proches de l'émergence. Un chorion translucide indique que la larve a émergé. Après l'éclosion de l'œuf, recherchez des toiles sur les parties de la fleur. Ouvrez les toiles et recherchez les dégâts causés par l'alimentation et les larves.
Commencer la surveillance des œufs de deuxième et troisième générations sur les baies une semaine après que les premiers papillons du vol concerné ont été capturés dans les pièges. Poursuivre la surveillance des œufs chaque semaine jusqu'à une semaine après le pic de vol. Inspecter 100 grappes, en en sélectionnant une par vigne. Continuer à surveiller les grappes pour détecter les dommages causés par l'alimentation (trous ou baies creuses), les toiles et la présence de larves.
Dans les pays où L. botrana est établie, les mesures de lutte sont ciblées sur la deuxième génération. Cela est dû en partie à l'émergence prolongée de la première génération et à la réinfestation possible par les vignobles voisins non traités. Toutefois, le traitement de la première génération est recommandé si les populations sont importantes ou si les traitements sont effectués à l'échelle d'une zone. Les insecticides sont moins efficaces après la fermeture des grappes. Plusieurs insecticides à risque réduit sont homologués pour être utilisés dans les vignes afin de lutter contre les larves de tortricidés. Il s'agit notamment de régulateurs de croissance des insectes, de spinosynes et de Bacillus thuringiensis.
La confusion sexuelle est étudiée en Europe depuis plusieurs années. Elle s'est avérée plus efficace lorsque les populations de tordeuses de la vigne sont faibles et lorsqu'elle est appliquée sur de grandes surfaces de plus de 10 acres ou sur l'ensemble du territoire.
De nombreux prédateurs et parasitoïdes sont signalés dans la littérature européenne. Parmi les parasitoïdes, on trouve 4 espèces de mouches tachinides et près de 100 espèces de guêpes parasites dans les familles des ichneumonides, des braconides, des ptéromalides et des chalicidés. Les parasites qui ont le plus d'impact sont ceux qui s'attaquent à la chrysalide d'hivernage. En Espagne, il s'agit des ptéromalidés Dibrachys affinis et D. cavusqui provoqueraient jusqu'à 70% la mortalité des nymphes, tandis qu'en Italie, les ichneumonides Dicaelotus inflexus et Capteur Campoplex sont les plus importants.
La littérature :
- Briere JF, Pracros P. 1998. Comparaison des modèles de croissance en fonction de la température avec le développement de Lobesia botrana (Lepidoptera : Tortricidae). Environ. Entomol. 27 : 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. dans las comarcas vitícolas valencianas. Bol. Serv. Plagas 7 : 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). In Los parasitos de la vid, estrategias de proteccion razonada. Madrid, Espagne. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33 : 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. Etude de la relation entre les captures de Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) dans les pièges à phéromones sexuelles et l'accumulation de degrés-jours dans les vignobles de Sherry (sud-ouest de l'Espagne). J. Appl. Ent. 125 : 9-14.
- Gabel B, Mocko V. 1986. Une simulation fonctionnelle de la pyrale de la vigne Lobesia botrana Den. Et Schiff. (Lep., Torticidae). J. Appl. Ent. 101 : 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. Prévision de l'activité de vol de Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) dans le sud-ouest de l'Espagne. J. Appl. Entomol. 133 : 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. Lutte contre Sparganothis pilleriana Schiff. et Lobesia botrana (Den. & Schiff). dans les vignobles allemands à l'aide de la confusion sexuelle par phéromone. Bull. IOBC/WPRS 25 : 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera : Tortricidae) : influence de l'infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thèse N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. Daphne gnidium, a possible native host plant of the European grapevine moth Lobesia botrana, stimates its oviposition. Un changement d'hôte est-il pertinent ? Chemoecol. 16 : 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. Attraction de la pyrale de la vigne vers des parties de plantes hôtes et non-hôtes dans la soufflerie : effets de la phénologie de la plante, du sexe et du statut d'accouplement. Entomol. Exp. Appl. 122 : 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Ent. 125 : 515-518.
- Moreau J, Benrey B, Thiéry. 2006. Le cépage affecte les performances larvaires et les performances reproductives des femelles de la pyrale de la vigne Lobesia botrana (Lepidoptera : Tortricidae). Bull. Entomol. Res. 96 : 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. Effets du méthoxyfénozide sur les œufs, les larves et les adultes de Lobesia botrana (Den & Schiff) (Lepidoptera : Torticidae).
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. In Les ravageurs de la vigne. Féret, Bordeaux. pp. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. La relation entre la dispersion et la survie des larves de Lobesia botrana et leur densité dans les inflorescences de vigne. Entomol. Exp. Appl. 84 : 109-114.
- Xuéreb A, Thiéry D. 2006. Le parasitisme larvaire naturel de Lobesia botrana (Lepidoptera : Torticidae) varie-t-il selon l'année, la génération, la densité de l'hôte et le cultivar de vigne ? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). In Lepidotteri dei fruttiferi e della vite. Milan, Italie. Pp. 85-88.
Modélisation de la pyrale de la vigne dans la plate-forme FieldClimate
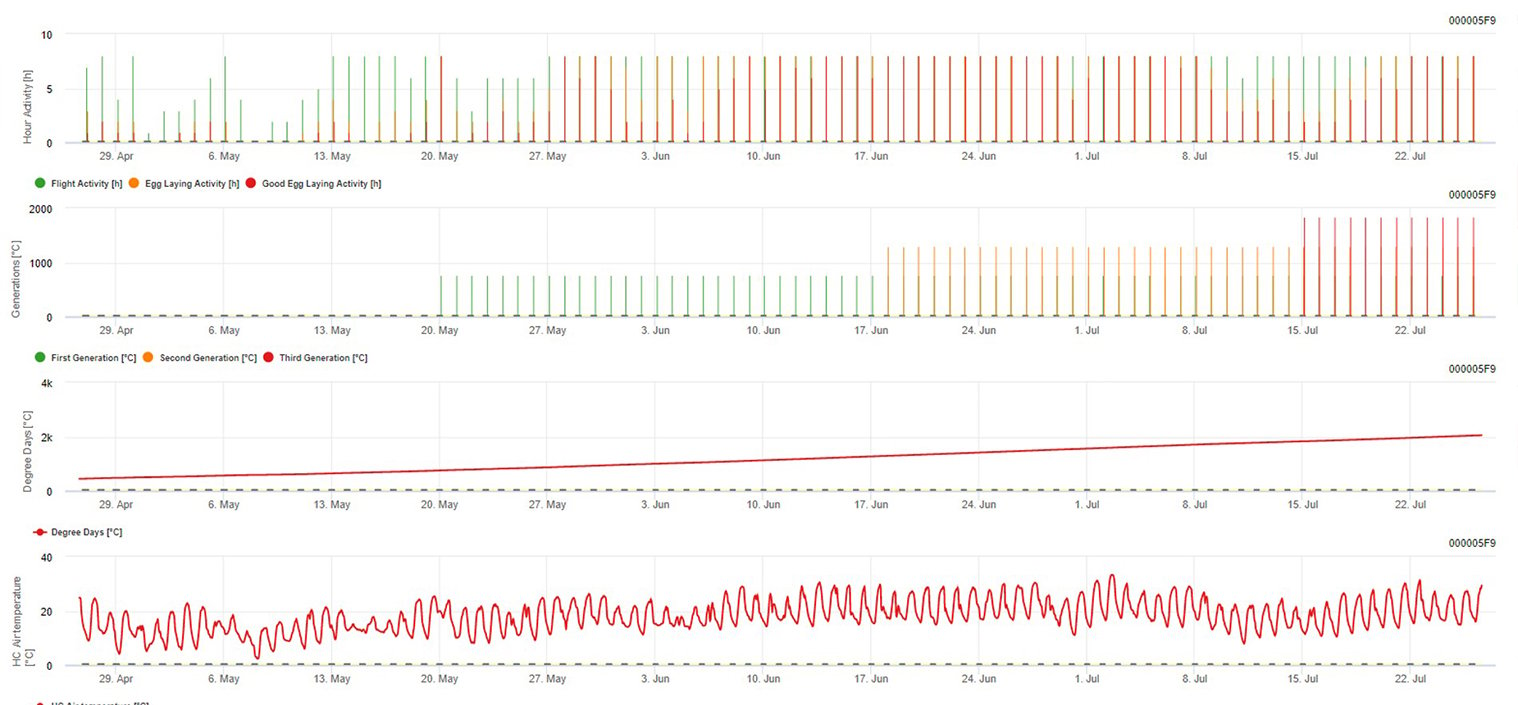
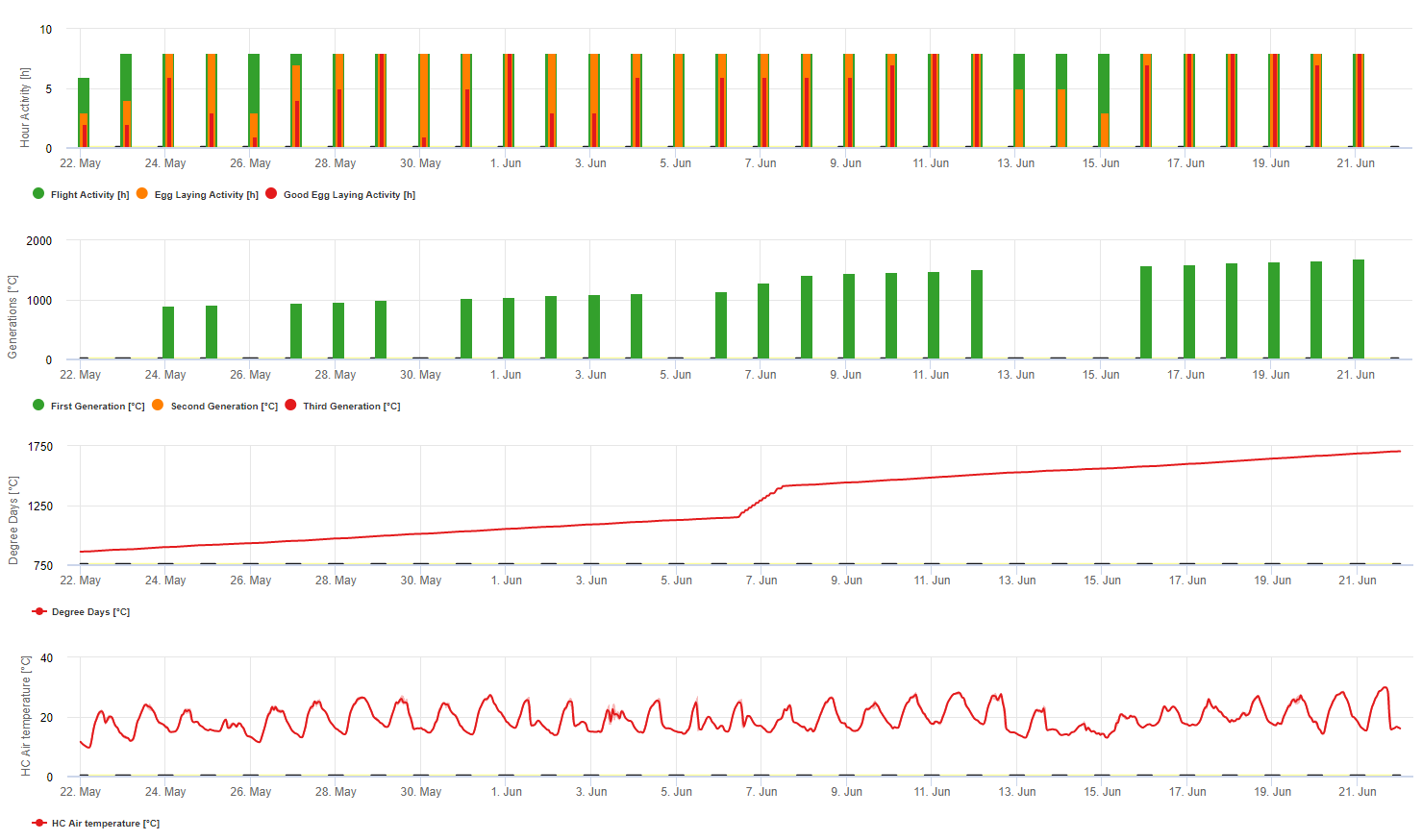
Capteur nécessaire : Température de l'air
Données horaires Sortie : activité de vol, activité de ponte, première génération, deuxième génération et troisième génération, degrés-jours cumulés
Pour l'activité de vol, les œufs et les bonnes activités de ponte : nous vérifions les données des 8 dernières heures (entre 19h00 et 03h00). Le seuil d'activité de vol est >12°C, celui de l'activité de ponte > 15°C et celui d'une bonne activité de ponte > 17°C. Si toutes les conditions d'activité de ponte sont remplies, nous générons la première génération (= degrés-jours cumulés). La première génération + 530 DJ est la deuxième génération (si elle n'a pas été prévue en raison des conditions précédentes). La deuxième génération + 530 DJ est la troisième génération.
Le graphique montre l'apparition de la première génération (bonnes conditions pour les trois activités de vol, de ponte et de bonne ponte) avec environ 469 degrés-jours accumulés. Le vol de la deuxième génération commence à la mi-juin avec environ 1010 degrés-jours accumulés et de bonnes conditions de température pour la ponte.
Botrytis cinerea est un champignon nécrotrophe qui affecte de nombreuses espèces végétales, bien que ses hôtes les plus remarquables soient les raisins. En viticulture, il est communément appelé pourriture de la grappe botrytis ; en horticulture, il est généralement appelé moisissure grise.
Le champignon donne lieu à deux types d'infections différentes sur les raisins. La première, la pourriture grise, est le résultat de conditions constamment humides ou mouillées, et entraîne généralement la perte des grappes affectées. La seconde, la pourriture noble, se produit lorsque des conditions plus sèches succèdent à des conditions plus humides, et peut donner naissance à des vins de dessert doux et distinctifs, tels que le Sauternes ou l'Aszú de Tokaj. Le nom de l'espèce Botrytis cinerea est dérivé du latin pour "raisins comme des cendres" ; bien que poétique, le "raisins" fait référence à l'agglutination des spores fongiques sur leurs conidiophores, et "cendres" fait simplement référence à la couleur grisâtre des masses de spores. Le champignon est généralement désigné par son nom anamorphe (forme asexuée) car la phase sexuelle est rarement observée. Le téléomorphe (forme sexuelle) est un ascomycète, Botryotinia cinerea.
Biologie de B. cinerea
Dans les fruits et les raisins, les infections florales suivies d'une latence ont un impact important sur l'épidémiologie de la moisissure grise. Plusieurs voies d'infection de la fleur au fruit peuvent être distinguées. Dans le raisin, les kiwis et les framboises, les infections par le stylet dans l'ovule ont été postulées. Dans l'ovule, l'agent pathogène reste latent, ce qui semble être le résultat d'une stratégie de défense préformée de l'hôte (similaire à la teneur en resvératrol des jeunes baies de raisin). Dans le raisin, les kiwis et les fraises, les infections par les étamines, les pétales ou les septales ont été jugées importantes. Dans le raisin, des études ont montré que B. cinerea pouvait infecter les étamines et se développer basipétalement pour infecter le réceptacle, puis se développer de manière systémique vers le pédicelle et les tissus vasculaires des baies.
Un projet de recherche de 6 ans dans la région du Cap a montré que les baies de raisin peuvent être infectées par le stoma et les lentilles du pédicelle et à un niveau inférieur du racheis. Les infections du pédicelle sont également possibles pendant la période de floraison. Plus tard, ce tissu augmente sa résistance contre B. cinerea infections.
D'autres voies d'infection postulent la croissance saprophyte de l'agent pathogène sur les débris floraux et l'infection ultérieure des baies lorsque la sensibilité augmente avec la maturation ou par des dégâts causés par les insectes ou la grêle sur les baies. Une autre thèse est celle de l'accumulation des conidies dans le fruit pendant l'été et l'infection des baies sensibles plus tard dans la saison. L'infection condidienne des fruits en cours de maturation est possible à partir de n'importe quelle source d'inoculum. Il est très probable qu'un faible nombre de baies infectées latentes se forment, qui présentent une sporulation extensive lorsque la sensibilité des baies augmente avec la maturité. On peut supposer que les baies deviennent sensibles à partir d'une teneur en sucre de 6%.
Dans le cas du kiwi, nous avons un impact important des conditions de cueillette sur l'apparition de B. cinerea. Les fruits cueillis avec une surface humide peuvent être infectés par B. cinerea aux microlésions provoquées par les doigts des cueilleurs.
Dans le contrôle pratique de B. cinerea nous devons séparer deux périodes d'infection importantes : La floraison et la sénescence. 1) Pendant la floraison, les jeunes fruits sont sensibles et l'infection est suivie d'une période de latence. 2) L'infection des fruits mûrs (sénescents) entraîne des symptômes sans période de latence. L'importance de l'infection pendant la floraison des raisins peut varier d'une saison à l'autre et d'une région à l'autre. Pour les fruits dont la durée de conservation est limitée (raisins de table, kiwis ou fraises), les symptômes apparaissent lorsqu'ils sont stockés dans des conditions réfrigérées dans des magasins ou des entrepôts. Une lutte chimique contre les raisins de cuve montrant une bonne résistance à la B. cinerea pendant la floraison ne donnera aucun résultat économique. Par conséquent, toutes les conditions de risque et de probabilité d'infection, la sensibilité du fruit et la durée de conservation, les conditions de stockage doivent être prises en considération dans la décision d'une demande contre les virus de la grippe aviaire. Botrytis cinerea pendant la floraison.
Dans les fruits à noyau, l'infection par B. cinerea se produit surtout pendant la floraison. A cette époque, les traitements contre Monilina spp. sont prises en compte, ce qui a également pour effet d'infecter le Botrytis cinerea l'infection.
Modèle de B. cinerea et l'utilisation pratique
Capteurs nécessaires : Humidité des feuilles, température, humidité relative
Botrytis cinerea Botrytis cinerea est un parasite fakultatif. Il se développe également sur le matériel végétal mort. Pour cette raison, il est toujours présent dans les vignobles et les vergers. Botrytis cinerea est lié au climat humide. Pour s'infecter, il a besoin d'une humidité relative très élevée ou de la présence d'eau libre (capteur : humidité des feuilles, humidité relative). Le champignon est incapable d'infecter le matériel végétal adulte sain par les spores. L'infection a lieu sur les jeunes pousses de la vigne pendant les longues périodes humides ou sur les pousses endommagées par les tempêtes de grêle.
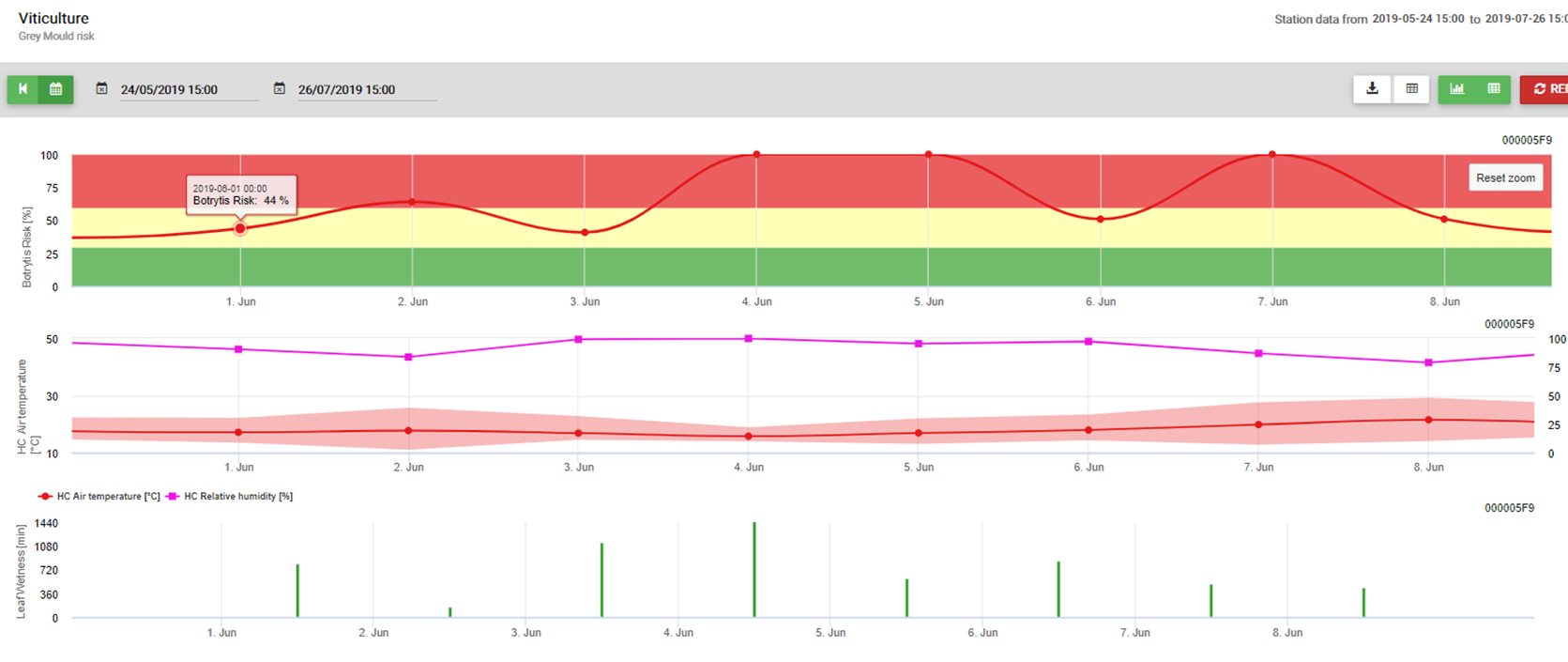
Le modèle utilise la corrélation suivante entre la durée d'humidité des feuilles et la température pour calculer le risque d'infection.
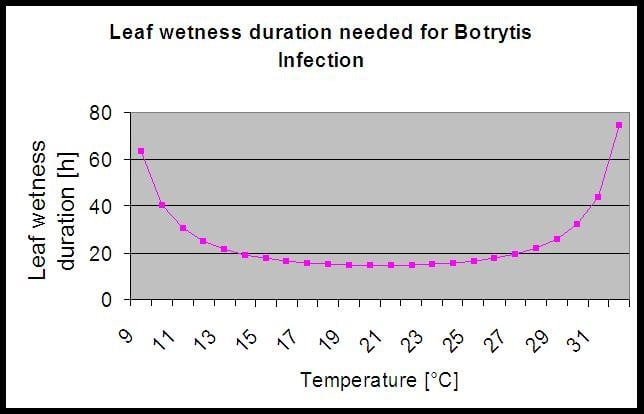
Le site Botrytis Le modèle de risque donne une valeur de risque de 0 à 100%. Cette valeur indique la pression de B. cinerea à ce moment-là. Si nous avons une valeur de 100%, cela signifie qu'il y a eu plusieurs fois une période d'humidité suffisamment longue pour infecter les tissus sensibles (nous calculons ce que l'on appelle les "points humides" (tableau entre l'humidité des feuilles, la température avec un maximum initial de 38400 points (début de la saison, qui affiche un risque de 30%). Après cette période, chaque période humide avec environ 4000 points humides (tableau) augmente le risque de 10% ou, à l'inverse, chaque période sèche réduit le risque de 1/5 de la valeur précédente.
Une demande contre B. cinerea dépend du fruit et de l'objectif de production.
L'anthracnose, la tache noire, la pourriture de l'œil d'oiseau, la tache de l'œil d'oiseau sont présentes dans le monde entier. La maladie réduit la qualité et le rendement des fruits, affaiblit la vigne par la destruction importante des nouvelles pousses et des feuilles. La plupart des espèces de Vitis sont sensibles à l'anthracnose.
Pendant les saisons humides de croissance de la vigne, la maladie est bien établie, elle provoque des dégâts sur les cultivars très sensibles après les pluies de début de saison.
Une fois établie dans les vignobles, la maladie peut être très difficile à gérer. Cependant, en Europe, la maladie est presque négligeable en raison des fongicides à base de composés de cuivre, de dithiocarbamates, de phtalimides pour lutter contre le mildiou et du manque de précipitations dans les climats arides.
Symptômes
La maladie attaque les parties aériennes et succulentes de la vigne, y compris les jeunes pousses, les feuilles, les pétioles, les vrilles et les grappes ; les lésions sur les pousses et les baies sont les plus courantes. Les premiers symptômes apparaissent sous la forme de petites taches circulaires isolées qui deviennent brunâtres, enfoncées avec un centre gris et des bords sombres, ronds ou anguleux. Les symptômes ressemblent parfois à des blessures causées par la grêle. Sur les feuilles, de nombreuses petites taches brunes apparaissent, dont le centre est gris, le centre nécrosé tombe généralement, créant un aspect de "trou de tir". Les jeunes feuilles sont plus sensibles aux infections. Sur les baies, de petites taches circulaires brun rougeâtre se développent et s'enfoncent légèrement. Le centre devient gris et est entouré de marges brun rougeâtre à noires (œil d'oiseau).
Cycle de vie de l'agent pathogène
La maladie est causée par l'agent pathogène Elsinöe ampelina (de Bary) Cisaillement.
Les cannes infectées sont la principale source de la maladie. Les sclérotes ou le mycélium qui survivent dans les lésions deviennent actifs au printemps et produisent des conidies dans des conditions humides (pluie ou rosée pendant 24 heures) à une température comprise entre 2°C et 40°C. Les conidies sont projetées par la pluie vers de nouveaux tissus. Elles germent et produisent de nouvelles infections. Le temps chaud réduit le temps d'humidification nécessaire à l'infection initiale et la période d'incubation avant que les symptômes ne soient visibles sur les feuilles. La germination optimale des spores se produit à 25-30°C, avec un minimum de 3 à 4 heures d'humidification des feuilles. La durée d'incubation est également la plus courte dans cette fourchette de température (3-4 jours dans des conditions idéales d'humidité). En cas d'infection à une température d'environ 10°C, il faut environ 14 jours pour que les symptômes de la maladie soient visibles sur les feuilles.
Les conidies ou les ascospores formées sur les baies infectées qui passent l'hiver sur la vigne ou le sol peuvent également provoquer des infections primaires. Les spores infectent les nouvelles feuilles, les pousses, les vrilles, les jeunes baies et produisent des lésions dans des conditions humides. Ces conidies servent d'inoculum secondaire et sont responsables de nouvelles infections au cours de la saison. Les conidies sont délogées par les gouttes de pluie et dispersées dans le vignoble.
Le développement épidémique est dû à la sensibilité des tissus de la vigne, à la dispersion des conidies et aux conditions météorologiques. Le facteur le plus important est la fréquence des périodes de pluie.
Modélisation dans Fieldclimate
Développement des spores pendant l'hiver : Température : 2-40C°, humidité relative supérieure à 90% ou humidité des feuilles - lorsque les spores sont développées (100%) et que les conditions d'humidité des feuilles et de température sont toujours présentes, l'infection commence à être calculée. Le développement des spores (humidité relative inférieure à 50%) et l'infection cessent d'être calculés lorsque l'humidité est trop faible. La sévérité de l'infection dépend des conditions humides (événement pluvieux).