





Cerise modèles de maladies
La maladie la plus importante chez les cerises est pourriture brunecausée par le champignon pathogène Monilia laxa. Une autre maladie importante, qui survient également dans des conditions d'humidité élevée, est la maladie du trou de tircausée par le champignon pathogène Wilsonmyces carpophilu et la maladie de tache foliaire du cerisiercausés par Blumeriella jaapii.
Accumulation de pluie
Une pluie intense emportera les pesticides résiduels sur les feuilles des vignes ou d'autres plantes. La résistance à la pluie des fongicides modernes s'est considérablement améliorée depuis 1980. Actuellement, on peut s'attendre à ce que les fongicides les plus modernes résistent à 30 mm de pluie s'ils ont eu la chance de sécher sur la feuille. Si la pluie commence immédiatement après la pulvérisation ou pendant la pulvérisation, la résistance à la pluie peut être largement réduite.
Avec les anciennes formulations de fongicides de contact, nous devons nous attendre à une dureté de la pluie inférieure à 12 mm. Comme nous y étions habitués dans les années 1970. Pour mouiller les feuilles dans un vignoble, il faut environ 2 mm de pluie. C'est pourquoi, dans cette mdoule, nous n'accumulons que les pluies de plus de 2 mm dans une période de mouillage des feuilles. Cela signifie qu'il pourrait y avoir au total 6 mm de pluie en un seul jour, mais que ce module n'en accumule aucune car les feuilles sont redevenues sèches avant qu'il ne pleuve 2 mm.
La pluie est accumulée pendant 3, 5 et 7 jours. Sur une période plus longue, la croissance des plantes est beaucoup plus importante pour l'effet des fongicides de contact que la résistance des composés à la pluie.
Modèle de la tavelure du fruit à noyau
La tavelure des fruits à noyau est induite par le champignon phytopathogène Cladosporium carpophilum. Le pathogène est présent sur les pêches, les nectarines, les abricots et les prunes, mais les pertes sont généralement plus importantes sur les pêches que sur les autres fruits.
La maladie affecte les rameaux, les feuilles et les fruits. Les dommages les plus graves résultent des infections des fruits.
Symptômes
Les lésions des fruits commencent par de petites taches rondes et verdâtres. Ces taches n'apparaissent généralement pas avant que le fruit soit à moitié développé, même si l'infection a eu lieu plus tôt dans la saison (environ six à sept semaines après la chute des pétales). Les lésions plus anciennes mesurent environ 1/4 de pouce de diamètre et prennent un aspect vert poussiéreux ou velouté. Les nombreuses lésions sont généralement regroupées près de l'extrémité de la tige du fruit (ce site est exposé au soleil). Des taches étendues peuvent entraîner des fissures dans les fruits, qui servent de points d'entrée à plusieurs champignons de pourriture des fruits. Les fruits peuvent également tomber prématurément ou ne pas être bien conservés.
Les feuilles peuvent également être infectées. De petites taches rondes, jaunâtres et vertes apparaissent sur la face inférieure de la feuille. Le tissu végétal peut se dessécher et tomber, laissant des trous de grenaille. En saison des pluies, les feuilles infectées tombent généralement tôt.
Sur les rameaux, les chancres commencent par de petites lésions rougeâtres sur la croissance de la saison en cours. Ces chancres s'étendent lentement et peuvent ne pas être visibles avant le milieu de l'été. Les petits chancres ont des bords irréguliers, mais ne provoquent pas de zones creuses sur l'écorce.
Sur les brindilles, les Le mycélium (ou conidies) hiberne sous forme de cellules sphériques brun foncé.. À partir du mycélium hivernant, des conidies sont produites au printemps, et ces dernières sont transportées vers les feuilles et les fruits. par le vent ou par la pluie. Les conditions qui favorisent le développement de la maladie sont des températures supérieures à 16°C pour la production de spores, plus de 10°C (optimal 22°C à 27°C) pour la germination des spores, et entre2°C et 35°C pour le développement de la maladie. La germination et la pénétration dans les tissus végétaux suivent peu après. Les inoculations et les infections se poursuivent jusqu'à environ un mois avant la maturité du fruit. Lorsque le champignon se développe sur le fruit, le mycélium se fixe étroitement à la surface entre les poils, formant un tapis de cellules courtes et dodues qui donnent naissance aux conidiophores et aux conidies. La chair de la pêche n'est pas pénétrée, mais le contact étroit du champignon avec les cellules externes permet l'absorption de la nutrition du fruit à travers les parois intactes. Il est évident que les cellules externes sont endommagées.
Chez FieldClimate, le risque d'une infection par Caldosporium carpophilum est déterminé par conditions humides au printemps et au début de l'été après la chute des pétales. La maladie est généralement plus grave chez zones basses, ombragées et humides avec faible mouvement d'air.
Dans FieldClimate, nous déterminons les infections dans une gamme de température de 7 à 24°C, avec une température optimale autour de 20°C.
Le modèle FieldClimate calcule en fonction de la durée d'humidité des feuilles et de la température un modèle de risque de Cladosporium carpohilum.
Trou de balle
Le trou de balle est causé par le champignon pathogène Wilsonmyces carpophilu (Coryneum blight).
Plus grave sur les abricots, mais se produit sur tous les fruits à noyau. Le champignon infecte les feuilles, les rameaux et les fruits.
Symptômes
Les feuilles infectées présentent de petites taches brunes avec des bords rougeâtres (environ 1 mm de diamètre), ces taches s'étendent à des lésions circulaires plus grandes (environ 3 mm de diamètre). Ces taches sèchent et tombent de la feuille, donnant un aspect de trou de balle. Les rameaux infectés présentent des marges brunes nettement découpées avec un centre négrotique, qui ne tombe pas, mais suinte de grandes quantités de gomme. Plus tard, la lignification des rameaux infectés est entravée et les lésions se transforment en chancres. Dans les cas graves, une défoliation prématurée de l'arbre peut en résulter.
Les fruits présentent d'abord de petites taches circulaires d'un violet profond. Au fur et à mesure que la maladie progresse, les symptômes diffèrent selon le type de fruit. Sur les abricots, les taches deviennent brunes, en relief et rugueuses, donnant au fruit une surface galeuse. Sur les pêches et les nectarines, les croûtes se transforment en profondes indentations.
Les fruits infectés présentent des taches de gomme et, dans les cas graves, des fissures dans la peau.
Le champignon shothole survit dans les bourgeons infectés. Il est capable d'infecter les feuilles, les tiges et les fruits au cours de l'année. temps froid et pluvieux périodes au printemps et en automne. Des périodes de pluie sont nécessaires pour infecter les organes sains des plantes.
Le champignon est capable de persister plusieurs années dans les chancres ou les bourgeons des rameaux infectés. Lorsque les conditions sont favorables, il peut continuer à se développer, même pendant l'hiver. Au printemps, les conidies sont projetées par la pluie sur les fleurs et les jeunes feuilles et les infectent. Dans les périodes défavorables (conditions sèches), les conidies sont encore viables pendant plusieurs mois. La pluie est nécessaire pour la dispersion et des conditions humides sont nécessaires pour la germination. Le champignon est capable de se développer au-dessus de 2°C.
La littérature :
- Adaskaveg JE, Ogawa JM, Butler EE (1990) Morphology and ontogeny of conidia in Wilsonomyces carpophilus, gen. nov., and comb. nov., causal
pathogène de la maladie du trou de balle des espèces de Prunuss. Mycotaxon 37, 275-290. - Ashkan M, Asadi P (1971) Shot hole of stone fruits in Iran. Iranian Journal of Plant Pathology 7, 39-63.Esfandiari E (1947) Les maladies des plantes cultivées et des arbres
fruitiers des régions subtropicales du nord de l'Iran. Entomologie et Phytopathologie Appliquées 5, 2. - Grove GG (2002) Influence de la température et de la période d'humidité sur l'infection du feuillage des cerises et des pêches par Wilsonomyces carpophilus. Canadian
Journal of Plant Pathology 24, 40-45. - Ogawa JM, Zehr EI, Bird GW, Ritchie DF, Uriu K, Uyemoto JK (1995) Compendium of stone fruit diseases. (The American Phytopathological
Society Press : St Paul, MN)
Oïdium
L'oïdium est une maladie courante sur de nombreux types de plantes. Plusieurs champignons de l'oïdium provoquent des maladies similaires sur des plantes différentes (comme le Podosphaera espèces sur les pommes et les fruits à noyau ; espèces de Sphaerotheca sur les baies et les fruits à noyau ; Erysiphe necator sur la vigne). Les champignons de l'oïdium nécessitent généralement conditions humides pour libérer les spores hivernantes et pour que ces spores germent et infectent les tissus végétaux. En revanche, aucune humidité n'est nécessaire pour que le champignon s'établisse et se développe après avoir infecté la plante. Les oïdiums favorisent normalement chaudClimats de type méditerranéen.
L'oïdium est facilement reconnaissable sur la plupart des plantes. par le mycélium poudreux blanc à gris. et de spores qui se forment sur les deux côtés des feuilles, des fleurs, des fruits et sur les pousses. Sur les fruits des arbres, une tache rugueuse et liégeuse se développe à l'endroit de l'infection.
Tous les champignons de l'oïdium ont besoin de tissus végétaux vivants pour se développer. Sur les plantes-hôtes vivaces à feuilles caduques telles que la vigne, le framboisier et les arbres fruitiers, l'oïdium survit à d'une saison à l'autre en les bourgeons infectés ou les organes de fructification appelés chasmothecia, qui résident sur l'écorce des cordons, des branches et des tiges.
La plupart des champignons de l'oïdium se développent sous la forme d'une fine couche de mycélium à la surface de la partie de la plante affectée. Les spores, qui sont le principal moyen de dispersion, constituent l'essentiel de la croissance poudreuse et sont produites en chaînes visibles à l'aide d'une loupe. En revanche, les spores du mildiou se développent sur des tiges ramifiées qui ressemblent à de petits arbres. De plus, les colonies de mildiou sont grises au lieu d'être blanches et se trouvent principalement sur la face inférieure des feuilles.
Les spores de l'oïdium sont transportées par vent aux plantes hôtes. Bien que les exigences en matière d'humidité pour la germination varient, de nombreuses espèces d'oïdium peuvent être présentes dans les plantes hôtes. germent et s'infectent en l'absence d'eau. En fait, les spores de certains champignons de l'oïdium sont tuées et la germination et la croissance mycélienne sont inhibées par l'eau à la surface des plantes. Températures modérées et ombre sont généralement les conditions les plus favorables au développement de l'oïdium, car les spores et le mycélium sont sensibles à la chaleur extrême et à la lumière directe du soleil.
Ce champignon passe l'hiver sous forme de mycéliums à l'intérieur des écailles de bourgeons, l'infection primaire se produit lorsque les feuilles émergent de ces bourgeons infectés. Les infections secondaires se produisent lorsque les conidies produites par les infections primaires et secondaires ultérieures sont soufflées ou éclaboussées par la pluie sur des tissus sensibles. Les fruits (avant le durcissement du noyau) et la croissance terminale succulente sont sensibles à l'infection.
Les températures moyennes minimales, optimales et maximales pour S. pannosa sont d'environ 5°, 24° et 24°C. Beaucoup plus de conidies sont formées dans l'air sec que dans l'air humide à toutes les températures (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli ; 1954).
L'oïdium est commun sous une humidité relative et des températures similaires à celles de l'oïdium du cerisier.
La littérature :
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954) : Les relations de température des oïdiums. HILGARDIA. A Journal of Agricultural Science publié par la California Agricultural Experiment Station. Université de Californie. Volume 22/Numéro 17.
Taphrina leaf curl
Peach leaf curl (pathogène fongique) : Taphrina deformans) est une maladie fongique qui peut causer une défoliation précoce grave et des pertes de récolte sur presque tous les cultivars de pêches et de nectarines.
Symptômes
Le symptôme le plus commun et le plus frappant de l'enroulement des feuilles se produit sur le feuilles (feuillage). Les feuilles infectées sont sévèrement déformées et présentent souvent une variété de couleurs (vert clair et jaune à des nuances de rouge et de pourpre). Le champignon provoque une prolifération rapide et aléatoire des cellules méristématiques sur les bords des feuilles, ce qui fait que les feuilles se plissent, se gondolent et s'enroulent (photo 2). Au fur et à mesure que ces feuilles infectées mûrissent, des asques nus contenant des ascospores de l'agent pathogène sont produits à la surface, leur donnant un aspect poussiéreux, après quoi les feuilles brunissent, se ratatinent et tombent de l'arbre.
Beaucoup de fruits infectés tombent rapidement et passent inaperçues ; celles qui restent peuvent devenir tordues à l'extrémité de la tige comme une petite courge jaune, tandis que d'autres deviennent rougeâtres à violettes et présentent des déformations "verruqueuses" en surface.
Cycle de la maladie
L'agent pathogène est communément présent presque partout où les pêches sont cultivées. Le champignon pathogène passe l'hiver sous forme de conidies (blastospores, spores "hyphales") dans des sites protégés de l'écorce et autour des bourgeons. Les infections primaires se produisent au début du printemps. Elle commence dès le gonflement des bourgeons jusqu'à l'apparition des premières feuilles sur les bourgeons. Les infections sur les jeunes feuilles de pêcher se produisent à des températures de 10°C à 21°C. Peu d'infections se produisent en dessous de 7°C. Les infections apparaissent principalement lorsque pluie laver les spores ayant hiverné dans les bourgeons et températures froides allonger la durée de développement des feuilles (elles sont exposées longtemps à l'agent pathogène avant d'être complètement développées et de pouvoir résister à la pénétration du champignon). Si températures après le gonflement des bourgeons sont chaud et les feuilles se développent rapidementles infections rarement s'établir, même en cas de pluies printanières. L'humidité due à la pluie (ou à d'autres facteurs) pendant plus de 12,5 heures est nécessaire pour l'infection des feuilles, mais seulement lorsque la température est inférieure à 16°C (61°F) pendant la période d'humidité. L'infection maximale se produit lorsque les arbres sont mouillés pendant 2 jours ou plus, ce qui est fréquent à l'ouest des Cascades. Bien qu'infectés, les symptômes peuvent ne pas apparaître si les températures restent supérieures à 21°C (69°F). Les fruits sont sensibles après la chute des pétales jusqu'à ce que la température de l'air reste supérieure à 19°C. Une pluie de 0,5 pouce et une humidité de 24 heures sont nécessaires pour l'infection des fruits.
Le risque d'un Taphrina defomans L'infection est calculée dans FieldClimate de deux manières :
- en utilisant les valeurs de température (ancien modèle).
- en utilisant l'accumulation de pluie pendant les dernières heures consécutives et la température pendant cette période. En outre, ce modèle inclut également le temps d'incubation (moment où les symptômes sont observés sur le terrain) en utilisant une température inférieure à 19°C pour les calculs.
Les deux modèles sont disponibles dans FieldClimate sous "Taphrina Leaf Curl disease".
Maladie des taches foliaires
La tache des feuilles du cerisier est causée par le champignon Blumeriella jaapii. La maladie réduit la floraison et affaiblit l'arbre. La tache des feuilles du cerisier défolie souvent l'arbre au milieu de l'été. La défoliation répétée rend l'arbre plus sensible aux dommages causés par l'hiver et peut éventuellement le tuer.
Symptômes
Les feuilles présentent de petites taches violettes à brunes avec des limites bien définies au début de l'été. Les feuilles jaunissent et tombent. En juillet, le centre des taches infectées tombe fréquemment, donnant un aspect de trou de balle.
Cause
Le champignon passe l'hiver sur les feuilles mortes. Au printemps, après un temps humide, des spores se forment et les vents les soufflent pour infecter les feuilles. Lorsque les températures sont favorables (16°C à 24°C), l'infection nécessite un délai de deux semaines. période humide de quelques heures seulement. Les feuilles sont sensibles lorsqu'elles sont complètement dépliées, ce qui se produit généralement au moment de la chute des pétales. Les infections initiales des feuilles forment des taches, et d'autres spores sont produites dans les taches. Celles-ci Les spores sont projetées par la pluie sur d'autres feuilles. et les infecter. La propagation secondaire et l'infection par les spores se poursuivent de manière répétée, à chaque fois qu'un temps chaud et humide se présente, jusqu'à la chute des feuilles en automne.
Cycle de vie
La tache des feuilles du cerisier est causée par le champignon ascomycète Blumeriella jaapii (anciennement connu sous le nom de Coccomyces hiemalis) anamorphe Phloeosporella padi (Lib.) Arx. Ce champignon passe l'hiver dans les feuilles mortes sur le sol comme sa principale forme de survie. Au printemps, des apothécies se développent sur ces feuilles. Les ascospores sont produites dans ces fructifications fongiques et sont éjectées de force. pendant les périodes pluvieuses pendant environ 6 à 8 semaines, à partir de la chute des pétales. Il s'agit du stade télémorphe ou sexuel, qui entraîne de nouvelles recombinaisons et variations génétiques pour la maladie. Cet agent pathogène affecte principalement et est le plus visible sur les feuilles des cerisiers. Les symptômes les plus évidents sont symptôme de cette maladie est le jaunissement des feuilles avant leur chute au début du printemps. Les feuilles infectées sont remplies de nombreux lésions noires qui couvrent une partie évidente de la feuille. Ces lésions se présentent d'abord sous la forme de petites taches violettes qui passent du rouge au brun puis au noir. La plupart des taches sont circulaires mais peuvent s'amalgamer pour former de grandes plaques mortes irrégulières. Ces taches ont tendance à s'agrandir et à se séparer des tissus sains. Enfin, elles tombent de la feuille en donnant un aspect de "trou de balle". Cet effet "shot hole" est plus fréquent sur les cerises acides que sur les cerises douces. Dans les cas plus graves et plus avancés, les lésions se retrouvent sur les fruits, les pétioles et les tiges. Les feuilles présentant des lésions tombent généralement au début de la saison de croissance et il est courant que les arbres gravement touchés soient défoliés au milieu de l'été. Les feuilles gravement infectées par la maladie deviennent très jaunes avant de tomber.
Si la maladie n'est pas contrôlée et qu'on la laisse persister pendant plusieurs années, elle peut avoir divers effets négatifs sur la production globale de cerises d'un arbre, principalement en raison du cycle de défoliation précoce et répétée. Ces effets comprennent des fruits nains et inégalement mûrs qui ont mauvais goût, des arbres plus sensibles aux blessures hivernales, la mort des pousses de fruits, des bourgeons fruitiers petits et faibles, une diminution de la taille et de la nouaison des fruits, et finalement la mort de l'arbre. Si la maladie a infecté les feuilles de manière assez importante au début du printemps, il est possible que les fruits ne parviennent pas à maturité cette année-là. Ces fruits resteront généralement de couleur claire, mous et très pauvres en solides solubles et en sucre.
Cycle de la maladie
Infection primaire
Ce champignon passe l'hiver dans les feuilles mortes sur le sol. Au début du printemps (vers la chute des pétales), des fructifications fongiques appelées apothécies se développent dans ces feuilles. Les spores (ascospores) sont produites dans les apothécies et sont libérées de force à partir de la chute des pétales. Ces ascospores sont propagées par le vent ou la pluie sur les nouvelles feuilles vertes et saines et servent ainsi d'inoculum primaire lors d'une nouvelle saison de croissance. Les ascospores peuvent adhérer à la feuille et, si un film d'eau et une température optimale sont présents, germer dans les heures qui suivent l'adhésion. Après la germination, les ascospores peuvent pénétrer dans la feuille par les stomates (ouvertures naturelles) sur la face inférieure de la feuille. Les ascospores ne peuvent pas pénétrer très efficacement dans les stomates immatures. Ces ascospores se développent dans les espaces intercellulaires de la feuille. Le temps d'incubation varie en fonction de la quantité d'humidité disponible et des températures. Les petites lésions violettes peuvent apparaître dans les 5 jours si les conditions sont humides et si la température reste stable entre 60 (15°C) et 68 degrés F (20°C). Cette période d'incubation peut toutefois durer jusqu'à 15 jours si les températures sont plus basses et les conditions plus sèches. En général, les feuilles deviennent moins sensibles à la maladie à mesure qu'elles vieillissent au cours de la saison de croissance. Le premier signe de la maladie peut se trouver sur les drageons proches du sol. On considère qu'il s'agit du cycle de reproduction primaire de la tache des cerises. Cependant, l'infection issue de ce cycle primaire est assez faible, mais elle est essentielle pour que l'agent pathogène puisse produire des spores secondaires.
Infection secondaire
Une fois que ces lésions de l'infection primaire se sont formées, il existe un potentiel d'infections secondaires. Les spores secondaires ou estivales, appelées conidies, se forment sur la face inférieure de la feuille à partir d'éruptions légèrement concaves appelées acervules. Ces conidies sont en forme de corne et très solubles dans l'eau. Les spores ont un aspect blanchâtre sur la face inférieure de la feuille. Elles sont se propagent par le vent ou la pluie lorsqu'elles sont ouvertes et peuvent provoquer des infections supplémentaires. Chacune de ces spores peut se multiplier et entraîner la production de milliers de conidies supplémentaires en un laps de temps relativement court. Toutes ces conidies contiennent exactement la même information génétique et ne sont que la réplication de l'ADN du champignon. À ce stade, l'infection fongique submerge la plante hôte et provoque la chute prématurée de ses feuilles. La litière de feuilles produite devient alors le site d'hivernage de ce champignon, et le cycle recommence au printemps suivant. Ces infections secondaires permettent à la maladie de se transformer très rapidement en épidémie.
Environnement
La maladie est généralement distribuée aux États-Unis, au Canada et dans diverses parties du monde où conditions humides se produire. Le champignon de la tache foliaire du cerisier préfère les conditions modérément humides, avec des températures supérieures à 60 degrés Fahrenheit (15°C). La plage de température optimale pour la propagation de ce champignon se situe entre 60 (15) et 68 degrés Fahrenheit (20°C). L'infection sérieuse d'un arbre se produit dans les années avec de nombreuses périodes de pluie et des étés plus frais. Très peu d'ascospores seront éjectées des apothécies si la température est inférieure à 46 degrés F (7°C). Cette maladie se développe dans les zones qui n'utilisent aucun type de pratique d'assainissement comme mesure préventive pour empêcher la propagation de la maladie. Lors de la sélection d'un site pour un verger, choisissez des zones qui ont un sol bien drainé, beaucoup de soleil et une bonne circulation d'air.
Modélisation de l'infection par Blumeriella jaapii
Le modèle de maturation des ascopsores : Sous des températures optimales, les ascopsores deviennent matures en 2 semaines. Les températures optimales sont supérieures à 16°C. Dans des conditions moins optimales, la période de décharge des ascospores peut prendre jusqu'à 6 semaines. Le modèle montre la période de libération possible des ascospores. Pendant cette période, nous devons nous attendre à des infections primaires de la maladie.
Le modèle de décharge de l'ascospore :
La décharge des ascospores dépend du mouillage des apothécies et du séchage qui suit. Elle est plus importante lorsque plus d'apothécies sont mouillées (pluie plus forte ou plus de pluie) et lorsque la température pendant et après la pluie est plus élevée. Elle débute à des températures de 16°C.
Le modèle d'infection de Blummeriella jaapii :
L'infection est prédite à l'aide du tableau d'Eisensmith et Jones (tableau 1), qui utilise les données suivantes durée d'humectation des feuilles et température de l'air données. Dans le modèle, une période de mouillage est initiée lorsque le capteur d'humidité de la feuille devient humide. La période de mouillage se termine lorsque l'humidité relative descend en dessous de 90% et que le capteur d'humidité de la feuille est sec. Cependant, si le capteur d'humidité de la feuille devient à nouveau humide dans les 8 heures suivant le moment où l'humidité relative est tombée en dessous de 90%, on considère qu'il s'agit de la même période d'humidification à laquelle on ajoute les heures de sécheresse. Une période d'humectation peut se traduire par l'absence d'infection ou par une infection par le pathogène de la tache des cerises.
Utilisation du modèle :
Le modèle de la tache foliaire du cerisier peut être utilisé comme guide pour déterminer le besoin de des pulvérisations éradicatrices si la couverture protectrice a été faible lors d'un épisode de pluie. Le modèle ne doit pas être utilisé comme un guide de routine pour les applications de pulvérisation éradicative - un programme de pulvérisation protectrice est l'approche la plus fiable pour la lutte contre la tache foliaire du cerisier. Le modèle est utile pour comparer l'activité d'infection de la tache foliaire prévue entre les années.
Modèle d'infection par les ascospores et les conidies basé sur :
- EISENSMITH, S.P. et A. L. Jones (1981) : Infection Model for timing fungicide applications to control cherry leaf spot. Plant Dis. 65
- Eisensmith, S. P. et A. L Jones. 1981. Un modèle pour détecter les périodes d'infection de Coccomyces hiemalis sur cerise acide. Phytopathology.71:728-732.
- Jones, A. L. et T. B. Sutton. 1996. Maladies des fruits des arbres dans l'est. Northcentral Regional Bulletin 45, Michigan State University.
- Shane, B. (2011) : A propos du rapport d'Enviro-weather sur les taches de feuilles de cerisier, voir ici.
Monilia spp.
La pourriture brune, causée par Monilia spp. (Monilia laxa, Monilia fructigena et Monilia fructicola) font partie des maladies les plus destructrices des fruits à noyau en Europe. L'impact économique est particulièrement important dans la production de cerises.
Symptômes
Les symptômes de la maladie de la pourriture brune sont le flétrissement de la fleur du cerisier ainsi que de l'extrémité verte des rameaux en raison de la pénétration du pathogène dans la fleur ouverte par le stigmate des pistils ou des anthères. Cela se traduit généralement par le flétrissement de toute la partie d'un rameau d'un an. Les feuilles commencent à pendre, puis deviennent brunes et rigides, mais ne tombent généralement pas sur le sol, elles restent sur l'arbre jusqu'au printemps de l'année suivante. Parfois, surtout dans des conditions humides, des gouttelettes de gomme sont visibles, qui sont des symptômes de la colonisation du champignon ainsi que des chancres établis. Les fruits infectés sont couverts de taches putréfiées, d'où apparaissent des sporodochies verruqueuses (hyphe) avec des conidies de la forme "été". De plus, à la fin de l'automne et en hiver, le champignon produit des sporodochies de la forme "hiver" sur les rameaux infectés. Avec le temps, les fruits sévèrement affectés se momifient. Le mycélium qui se développe dans ces momies s'agrège progressivement en sclérotes. Ces fruits restent sur l'arbre pendant l'hiver.
Température et durée de l'humidité sont des facteurs environnementaux importants, déterminant l'incidence de l'infection par M. laxa sur les fleurs de cerisier. Monilia laxa est bien adapté à la des températures relativement basses au printemps et provoquent des infections à des températures aussi basses que 5°C dans une très courte période de durée d'humidité. L'infection de la fleur active à travers le stimulus n'a pas besoin de beaucoup de temps. humidité des feuilles. Humidité des feuilles n'est nécessaire que pour la germination des conidies. Par conséquent, l'infection des jeunes fruits nécessite des périodes d'humidité foliaire plus longues. Pour infecter les jeunes fruits, un appressoria doit être formé et l'humidité libre est nécessaire pour créer la pression nécessaire à la formation de la cheville d'infection qui pénètre dans la cellule de l'épiderme. Avec la maturité des fruits, de petites cicatrices sur les fruits permettent à nouveau une infection sans appressoria et la durée d'humidification des feuilles devient plus courte.
Aucune sporulation n'est possible si l'humidité relative ne dépasse pas 85%. Le modèle M.laxa de fieldclimate.com calcule les infections faibles et sévères. Infections faibles sont modélisés pour les conditions permettant d'infecter les stades hautement sensibles comme les fleurs et les fruits blessés proches de la maturité. Pour infecter ces tissus, de très courtes périodes d'humidité des feuilles se sont avérées suffisantes. Plusieurs études, par exemple : " Phenological Analysis of Brown Rot Blossom Blight of Sweet Cherry Caused by Monilinia laxa" (L. Tamm, Chr. E. Minder, et W. Flickiger ; 1994) ou "Effects of wounding, fruit age and wetness duration on the development of cherry brown rot in the UK.". ( X.-M. Xu*, C. Bertone et A. Berrie ;2003) ont confirmé les faibles périodes d'humidité nécessaires aux infections. Plus loin, les fruits immatures sont plus résistants aux infections, mais à proximité de la maturité, ils deviennent plus sensibles et les conditions pour des infections sévères sont données.
Les modèles de dispersion printanière de Monilinia laxa conidies dans les abricotiers, les pêchers, les pruniers et les amandiers. Journal canadien de botanique (1974), 52 : 167-176
Modélisation du climat des champs : Il est probable que le temps nécessaire à l'infection pendant la floraison doive être raccourci. Par conséquent, le modèle réduit les infections dans la zone de 2000 à 4800 degrés-heure au-dessus de 5°C.
Thrips des fleurs de l'Ouest
Les thrips des fleurs de l'Ouest sont des insectes minuscules, d'environ 0,03 pouce de long, avec deux paires d'ailes frangées. L'adulte a trois formes de couleur qui varient en abondance selon la période de l'année. Il existe une forme pâle qui est blanche et jaune, à l'exception de légères taches ou imperfections brunes sur le dessus de l'abdomen ; une forme de couleur intermédiaire avec un thorax orange et un abdomen brun ; et une forme sombre qui est brun foncé. La forme intermédiaire est présente toute l'année, mais au printemps, la forme sombre prédomine tandis que la forme pâle est plus abondante à d'autres moments de l'année.
Les nymphes du premier stade sont opaques ou jaune clair, et deviennent jaune doré après la première mue. Le stade nymphal dure de 5 à 20 jours.
Dégâts
Les nymphes éclosent et se nourrissent en nombre sur les fruits, souvent sous le calice ou les parties de la fleur en train de sécher. Leur alimentation provoque des cicatrices à la surface du fruit. Ces cicatrices s'agrandissent au fur et à mesure que le fruit grandit, et peuvent provoquer une déformation du fruit. Les thrips peuvent également provoquer un argentage juste avant la maturité des fruits.
Bien qu'une certaine alimentation ait lieu sur les fleurs, peu de dommages sont causés jusqu'à la formation des fruits. Les thrips peuvent endommager les pousses terminales et en arrêter la croissance. Généralement, une ou deux petites feuilles mortes s'accrochent à la terminaison. Les bourgeons situés juste en dessous de la terminaison poussent, donnant à la branche un aspect touffu.
Gestion
Le thrips des fleurs de l'Ouest passe l'hiver sous forme d'adultes dans les mauvaises herbes, les graminées, la luzerne et d'autres hôtes, soit dans le sol du verger, soit à proximité. Au début du printemps, si les sites d'hivernage sont perturbés ou s'assèchent, les thrips migrent vers les arbres et les plantes en fleurs et déposent leurs œufs dans les parties tendres de la plante hôte, par exemple les pousses, les bourgeons et les parties florales.
Contrôle culturel
Les thrips sont souvent attirés par les mauvaises herbes qui fleurissent sur le sol du verger. Pour éviter d'attirer les thrips dans les arbres, ne pas défaire la culture de couverture lorsque les arbres sont en fleurs. Les terres ouvertes et envahies de mauvaises herbes adjacentes aux vergers doivent être éliminées le plus tôt possible pour empêcher le développement des thrips et la migration des adultes dans les vergers.
Méthodes biologiquement acceptables
Les contrôles culturels, la culture propre et les pulvérisations de la formulation Entrust de spinosad sont des outils acceptables sur le plan biologique.
Suivi et décisions de traitement
Commencez à surveiller les thrips lorsque les arbres commencent à fleurir. La surveillance des thrips s'effectue en examinant les fleurs des arbres en frappant une pousse de cinq à dix fleurs contre une carte jaune ou en recherchant les stades immatures dans les fleurs. Souvent, les nymphes ne sont pas délogées par la méthode des claques. Il faut donc disséquer les fleurs individuelles et les examiner à l'aide d'une loupe pour détecter les nymphes. Les nymphes de premier stade sont de couleur blanche et sont souvent difficiles à voir, alors assurez-vous de bien vérifier. Vérifiez au moins 50 arbres par verger à la recherche de nymphes. Au cours des printemps chauds, les adultes migrent souvent dans et hors d'un bloc sans être détectés, il est donc important de toujours échantillonner pour les nymphes.
Si deux thrips adultes ou plus sont présents ou si des nymphes sont trouvées, un traitement est justifié. Si un traitement est appliqué, faites-le avant que le calice ne se resserre autour de l'ovaire en développement. Si des nymphes sont trouvées sous l'enveloppe après qu'elle se soit resserrée autour du fruit, utilisez du méthomyl.
Source : Littérature de UC IPM Davis :
- K. R. Day, UC Cooperative Extension, comté de Tulare
- K.Tollerup, Programme UC IPM, Centre agricole de Kearney, Parlier.
Dans FieldClimate, nous utilisons les conditions suivantes pour le calcul du modèle de risque :
Modèle de risque pour le thrips des fleurs de l'ouest Frankliniella occidentalis (Pergande) sur la base des valeurs quotidiennes :
- Le risque augmente lorsque la température de l'air est supérieure à 18°C et inférieure à 32°C et que l'humidité relative est supérieure à 70% par 20%.
- Le risque diminue lorsque la température de l'air est supérieure à 32°C pendant plus de 8 heures ou lorsque la température de l'air est supérieure à 32°C pendant plus de 8 heures.
- Si l'humidité relative n'est jamais supérieure à 70% ou si toutes les températures sont inférieures à 15°C, réduire le risque de 10%.
Chancre bactérien
Le chancre bactérien est une maladie des tiges et des feuilles des Prunus, en particulier des prunes et des cerises, mais aussi des abricots, des pêches et des espèces ornementales de Prunus. Il provoque des plaques d'écorce morte et de petits trous dans les feuilles.
Les espèces P. syringae existe sous la forme d'un grand nombre de pathovars (abrégé en pv.), appelés ainsi parce que, bien que tous se ressemblent, ils ont des hôtes différents et spécifiques. Le pathovar morsprunorum est limité aux espèces de Prunus, pv. syringae a une gamme d'hôtes beaucoup plus large, mais les deux provoquent des symptômes similaires sur Prunus.
Les bactéries vivent en surface (épiphytes) sur les feuilles et, par temps humide au printemps ou au début de l'été, elles peuvent pénétrer par les pores des feuilles (stomates), provoquant des infections dans les jeunes feuilles. Au fur et à mesure que la feuille mûrit, ces infections cessent de se développer et apparaissent sous la forme de petites plaques de tissus morts. Lorsque la feuille se développe complètement, les tissus vivants se détachent de la plaque morte, qui tombe, laissant un "trou de balle".
Les chancres se développent lorsque les cellules bactériennes pénètrent par les blessures ou les cicatrices des feuilles au moment de leur chute. Les chancres restent plus ou moins dormants pendant l'été, lorsque les tissus sont résistants, et pendant l'automne et l'hiver, lorsque les températures sont basses. Au printemps, les infections se propagent rapidement, tuant l'écorce.
Symptômes
Vous pouvez observer les symptômes suivants :
X) Sur les tiges et les éperons : Des zones d'écorce mortes et enfoncées se développent au printemps et au début de l'été, souvent accompagnées d'un suintement gommeux. Si l'infection se propage tout autour de la branche, celle-ci meurt rapidement. Cependant, il faut noter que la production de gomme (gummose) à partir de l'écorce des espèces de Prunus est en fait assez courante, et en l'absence d'écorce morte et enfoncée, il est probable qu'elle résulte de causes autres que le chancre bactérien, par exemple des dommages physiques ou des stress environnementaux.
Sur les feuilles : De petites taches brunes apparaissent, souvent rondes, qui se détachent ensuite pour laisser des trous, comme si la feuille avait été touchée par des plombs de chasse, d'où le nom populaire de "trou de balle".
Lutte non chimique
Dans la mesure du possible, effectuez toutes les tailles en juillet ou en août, lorsque les tissus sont les plus résistants. C'est également le meilleur moment pour tailler afin de minimiser le risque d'infection par les spores du champignon responsable de la maladie de la feuille d'argent. Coupez toutes les zones chancreuses, taillez jusqu'au bois sain et peignez rapidement avec une peinture pour plaies afin de protéger la plaie contre toute réinfection.
Source : Société royale d'horticulture
Modèle FieldClimate
Modèle de risque basé sur les valeurs quotidiennes :
- si la tension de l'eau du sol pendant toute la journée est inférieure à 25, si la pluie est supérieure à 2 mm et si la durée d'humidité de la feuille est supérieure à 6 heures, le risque augmente de 20% autrement
- si la tension de l'eau du sol pendant toute la journée est supérieure à 40 le risque diminue de 20% ou
- si pas d'humidité foliaire toute la journée le risque diminue de 10% ou
- si aucune heure avec plus de 70% rel le risque humide diminue de 10%
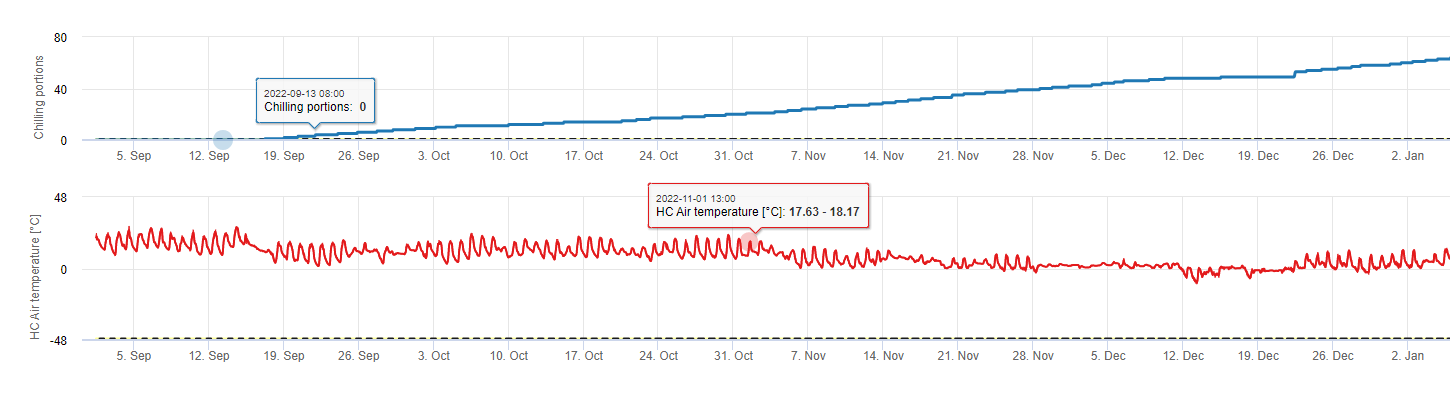
Portions réfrigérantes
Refroidissement
Les arbres fruitiers à noyau développent leurs bourgeons végétatifs et fructifères en été et, à l'approche de l'hiver, les bourgeons déjà développés entrent en dormance en réponse à la diminution de la durée du jour et aux températures plus fraîches. Ce stade de dormance ou de sommeil protège ces bourgeons du froid qui s'annonce. Une fois que les bourgeons sont entrés en dormance, ils seront tolérants à des températures bien inférieures au point de congélation et ne se développeront pas en réponse aux périodes de chaleur du milieu de l'hiver. Ces bourgeons restent en dormance jusqu'à ce qu'ils aient accumulé suffisamment d'unités de réfrigération (UC) de temps froid. Lorsque suffisamment de réfrigération s'accumule, les bourgeons sont prêts à se développer en réponse aux températures chaudes. Tant qu'il y a eu suffisamment d'UC, les bourgeons floraux et foliaires se développent normalement. Si les bourgeons ne reçoivent pas suffisamment de températures froides en hiver pour libérer complètement la dormance, les arbres développeront un ou plusieurs des symptômes physiologiques associés à un refroidissement insuffisant : 1) une foliation retardée, 2) une réduction de la nouaison et une augmentation du boutonnage et, 3) une réduction de la qualité des fruits.
Symptômes de refroidissement insuffisant
Foliation retardée :
Le retard de la foliation est un symptôme classique d'un refroidissement insuffisant. Un arbre peut avoir une petite touffe de feuilles près de l'extrémité des tiges et être dépourvu de feuilles sur 12 à 20 pouces sous l'extrémité. Les bourgeons inférieurs finiront par éclater, mais la foliation complète est considérablement retardée, la nouaison est réduite et l'arbre est affaibli. De plus, un drageonnage important des parties inférieures de l'arbre pose des problèmes de gestion, et le développement normal des bourgeons à fruits de l'année suivante peut être entravé.
Réduction de la nouaison et du boutonnage :
La floraison, en réponse à un refroidissement insuffisant, suit souvent le schéma observé pour le développement des feuilles. La floraison est retardée, prolongée et, en raison d'anomalies dans le développement du pistil et du pollen, la nouaison est réduite. Chez de nombreux cultivars de pêchers, les fleurs tombent avant ou autour de l'éclatement du fruit, mais chez d'autres, comme 'Jersey Queen' et 'Harvester', des boutons se forment. Les boutons sont le résultat de fleurs qui semblent avoir été fixées mais qui ne se transforment jamais en fruits de taille normale. Les fruits restent petits et difformes à mesure qu'ils mûrissent. Si vous ouvrez ces fruits, la graine est morte. Comme la formation de boutons n'est pas apparente en début de saison, les cultivateurs ne peuvent pas éclaircir les fruits anormaux et les boutons en développement servent de source de nourriture et de site d'hivernage pour les insectes et les maladies.
Réduction de la qualité des fruits :
Les effets d'un refroidissement insuffisant sur la qualité des fruits sont probablement ceux dont on parle le moins, mais ils semblent être très courants, surtout dans le centre et le sud du Texas. Les effets sur la croissance des feuilles et la nouaison sont spectaculaires, mais les effets d'un refroidissement insuffisant sur la qualité des fruits sont subtils et peuvent se produire alors que d'autres symptômes ne se manifestent pas. Un refroidissement insuffisant entraîne chez de nombreux cultivars un élargissement de la pointe et une réduction de la fermeté. De plus, la coloration du fond du fruit peut être plus verte que d'habitude, peut-être parce que le fruit perd de sa fermeté avant que la couleur du fond ne passe complètement du vert au jaune. L'ampleur de ces problèmes de qualité dépend du cultivar et du degré de carence en froid.
Modèles
Il existe plusieurs modèles utilisés pour calculer la réfrigération, chacun définissant ce qu'est une unité de réfrigération. Les trois modèles les plus courants sont le modèle du nombre d'heures en dessous de 45 degrés F (7°C), le modèle du nombre d'heures entre 32 et 45 degrés F (2 et 7°C) et le modèle de l'Utah. Les deux premiers modèles sont simples et définissent une unité de réfrigération comme une heure en dessous ou entre certaines températures. La méthode d'Utah est plus complexe car elle introduit le concept d'efficacité frigorifique relative et d'accumulation frigorifique négative (ou négation frigorifique).
Dans FieldClimate, nous utilisons le modèle de calcul des portions de réfrigération (CP). Les accumulations de froid sont calculées en tant que proportions de froid, en utilisant une gamme de température de 2 à 7°C. Les calculs des proportions de refroidissement se terminent après 96 heures de température égale ou supérieure à >15°C '(elle se maintient entre7 et 15°C)
Les calculs sont basés sur les travaux de Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276 : 165-174.
Équipement recommandé
Vérifiez quel jeu de capteurs est nécessaire pour surveiller les maladies potentielles de cette culture.