Conditions :
- le matin, lorsque le soleil se lève et que l'humidité relative diminue, les températures optimales se situent entre 20°C et 32°C - un bon vol est indiqué.
- Si les températures ne sont pas dans la fourchette optimale (trop froides/chaudes) ou si elles sont trop humides (humidité des feuilles), le risque diminue.
- La sortie est le risque quotidien.
Ainsi, des températures optimales et des humidités relatives en baisse durant la matinée indiquent une bonne journée de vol. Lorsqu'il fait humide pendant la nuit et que les températures sont trop basses, c'est mauvais pour la propagation. Il en va de même lorsqu'il fait chaud et humide pendant la journée.
L'oïdium est une maladie courante sur de nombreux types de plantes. Plusieurs champignons de l'oïdium provoquent des maladies similaires sur des plantes différentes (comme le Podosphaera espèces sur les pommes et les fruits à noyau ; espèces de Sphaerotheca sur les baies et les fruits à noyau ; Erysiphe necator sur la vigne). Les champignons de l'oïdium nécessitent généralement conditions humides pour libérer les spores hivernantes et pour que ces spores germent et infectent les tissus végétaux. En revanche, aucune humidité n'est nécessaire pour que le champignon s'établisse et se développe après avoir infecté la plante. Les oïdiums favorisent normalement chaudClimats de type méditerranéen.
L'oïdium est facilement reconnaissable sur la plupart des plantes. par le mycélium poudreux blanc à gris. et de spores qui se forment sur les deux côtés des feuilles, des fleurs, des fruits et sur les pousses. Sur les fruits des arbres, une tache rugueuse et liégeuse se développe à l'endroit de l'infection.

Tous les champignons de l'oïdium ont besoin de tissus végétaux vivants pour se développer. Sur les plantes-hôtes vivaces à feuilles caduques telles que la vigne, le framboisier et les arbres fruitiers, l'oïdium survit à d'une saison à l'autre en les bourgeons infectés ou les organes de fructification appelés chasmothecia, qui résident sur l'écorce des cordons, des branches et des tiges.
La plupart des champignons de l'oïdium se développent sous la forme d'une fine couche de mycélium à la surface de la partie de la plante affectée. Les spores, qui sont le principal moyen de dispersion, constituent l'essentiel de la croissance poudreuse et sont produites en chaînes visibles à l'aide d'une loupe. En revanche, les spores du mildiou se développent sur des tiges ramifiées qui ressemblent à de petits arbres. De plus, les colonies de mildiou sont grises au lieu d'être blanches et se trouvent principalement sur la face inférieure des feuilles.
Les spores de l'oïdium sont transportées par vent aux plantes hôtes. Bien que les exigences en matière d'humidité pour la germination varient, de nombreuses espèces d'oïdium peuvent être présentes dans les plantes hôtes. germent et s'infectent en l'absence d'eau. En fait, les spores de certains champignons de l'oïdium sont tuées et la germination et la croissance mycélienne sont inhibées par l'eau à la surface des plantes. Températures modérées et ombre sont généralement les conditions les plus favorables au développement de l'oïdium, car les spores et le mycélium sont sensibles à la chaleur extrême et à la lumière directe du soleil.
Ce champignon passe l'hiver sous forme de mycéliums à l'intérieur des écailles de bourgeons, l'infection primaire se produit lorsque les feuilles émergent de ces bourgeons infectés. Les infections secondaires se produisent lorsque les conidies produites par les infections primaires et secondaires ultérieures sont soufflées ou éclaboussées par la pluie sur des tissus sensibles. Les fruits (avant le durcissement du noyau) et la croissance terminale succulente sont sensibles à l'infection.
Les températures moyennes minimales, optimales et maximales pour S. pannosa sont d'environ 5°, 24° et 24°C. Beaucoup plus de conidies sont formées dans l'air sec que dans l'air humide à toutes les températures (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli ; 1954).
L'oïdium est commun sous une humidité relative et des températures similaires à celles de l'oïdium du cerisier.
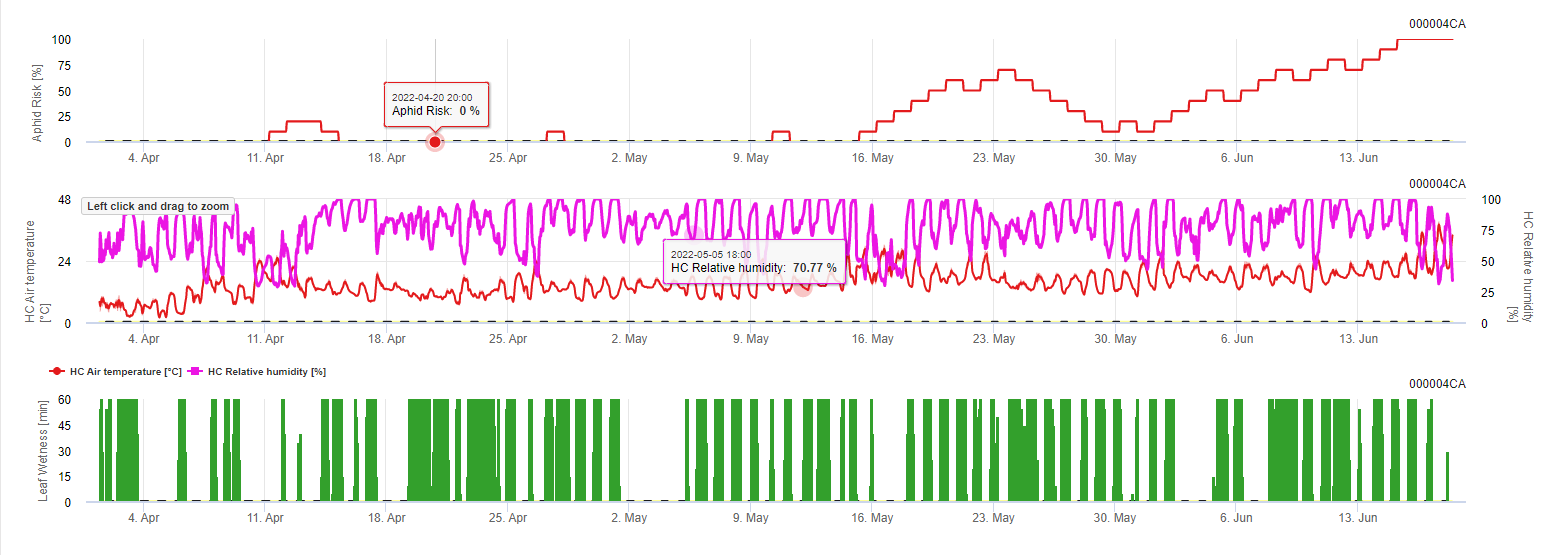
Modèle Fieldclimate : La maladie fongique est modélisée par les facteurs température et durée d'humidité des feuilles. Par exemple, le 11 mai, la période d'humidité des feuilles sous des températures modérées a favorisé le développement de la maladie et un risque de 100% a pu être déterminé.
La littérature :
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954) : Les relations de température des oïdiums. HILGARDIA. A Journal of Agricultural Science publié par la California Agricultural Experiment Station. Université de Californie. Volume 22/Numéro 17.
Refroidissement
Les arbres fruitiers à noyau développent leurs bourgeons végétatifs et fructifères en été et, à l'approche de l'hiver, les bourgeons déjà développés entrent en dormance en réponse à la diminution de la durée du jour et aux températures plus fraîches. Ce stade de dormance ou de sommeil protège ces bourgeons du froid qui s'annonce. Une fois que les bourgeons sont entrés en dormance, ils seront tolérants à des températures bien inférieures au point de congélation et ne se développeront pas en réponse aux périodes de chaleur du milieu de l'hiver. Ces bourgeons restent en dormance jusqu'à ce qu'ils aient accumulé suffisamment d'unités de réfrigération (UC) de temps froid. Lorsque suffisamment de réfrigération s'accumule, les bourgeons sont prêts à se développer en réponse aux températures chaudes. Tant qu'il y a eu suffisamment d'UC, les bourgeons floraux et foliaires se développent normalement. Si les bourgeons ne reçoivent pas suffisamment de températures froides en hiver pour libérer complètement la dormance, les arbres développeront un ou plusieurs des symptômes physiologiques associés à un refroidissement insuffisant : 1) une foliation retardée, 2) une réduction de la nouaison et une augmentation du boutonnage et, 3) une réduction de la qualité des fruits.
Symptômes de refroidissement insuffisant
Foliation retardée :
Le retard de la foliation est un symptôme classique d'un refroidissement insuffisant. Un arbre peut avoir une petite touffe de feuilles près de l'extrémité des tiges et être dépourvu de feuilles sur 12 à 20 pouces sous l'extrémité. Les bourgeons inférieurs finiront par éclater, mais la foliation complète est considérablement retardée, la nouaison est réduite et l'arbre est affaibli. De plus, un drageonnage important des parties inférieures de l'arbre pose des problèmes de gestion, et le développement normal des bourgeons à fruits de l'année suivante peut être entravé.
Réduction de la nouaison et du boutonnage :
La floraison, en réponse à un refroidissement insuffisant, suit souvent le schéma observé pour le développement des feuilles. La floraison est retardée, prolongée et, en raison d'anomalies dans le développement du pistil et du pollen, la nouaison est réduite. Chez de nombreux cultivars de pêchers, les fleurs tombent avant ou autour de l'éclatement du fruit, mais chez d'autres, comme 'Jersey Queen' et 'Harvester', des boutons se forment. Les boutons sont le résultat de fleurs qui semblent avoir été fixées mais qui ne se transforment jamais en fruits de taille normale. Les fruits restent petits et difformes à mesure qu'ils mûrissent. Si vous ouvrez ces fruits, la graine est morte. Comme la formation de boutons n'est pas apparente en début de saison, les cultivateurs ne peuvent pas éclaircir les fruits anormaux et les boutons en développement servent de source de nourriture et de site d'hivernage pour les insectes et les maladies.
Réduction de la qualité des fruits :
Les effets d'un refroidissement insuffisant sur la qualité des fruits sont probablement ceux dont on parle le moins, mais ils semblent être très courants, surtout dans le centre et le sud du Texas. Les effets sur la croissance des feuilles et la nouaison sont spectaculaires, mais les effets d'un refroidissement insuffisant sur la qualité des fruits sont subtils et peuvent se produire alors que d'autres symptômes ne se manifestent pas. Un refroidissement insuffisant entraîne chez de nombreux cultivars un élargissement de la pointe et une réduction de la fermeté. De plus, la coloration du fond du fruit peut être plus verte que d'habitude, peut-être parce que le fruit perd de sa fermeté avant que la couleur du fond ne passe complètement du vert au jaune. L'ampleur de ces problèmes de qualité dépend du cultivar et du degré de carence en froid.
Modèles
Il existe plusieurs modèles utilisés pour calculer la réfrigération, chacun définissant ce qu'est une unité de réfrigération. Les trois modèles les plus courants sont le modèle du nombre d'heures en dessous de 45 degrés F (7°C), le modèle du nombre d'heures entre 32 et 45 degrés F (2 et 7°C) et le modèle de l'Utah. Les deux premiers modèles sont simples et définissent une unité de réfrigération comme une heure en dessous ou entre certaines températures. La méthode d'Utah est plus complexe car elle introduit le concept d'efficacité frigorifique relative et d'accumulation frigorifique négative (ou négation frigorifique).
Dans fieldClimate.com, nous utilisons les éléments suivants modèle de calcul des portions de réfrigération (CP). Les accumulations de froid sont calculées en tant que proportions de froid, en utilisant une gamme de température de 2 à 7°C. Les calculs des proportions de refroidissement se terminent après 96 heures de température égale ou supérieure à >15°C '(elle se maintient entre7 et 15°C)
Les calculs sont basés sur les travaux de Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276 : 165-174.