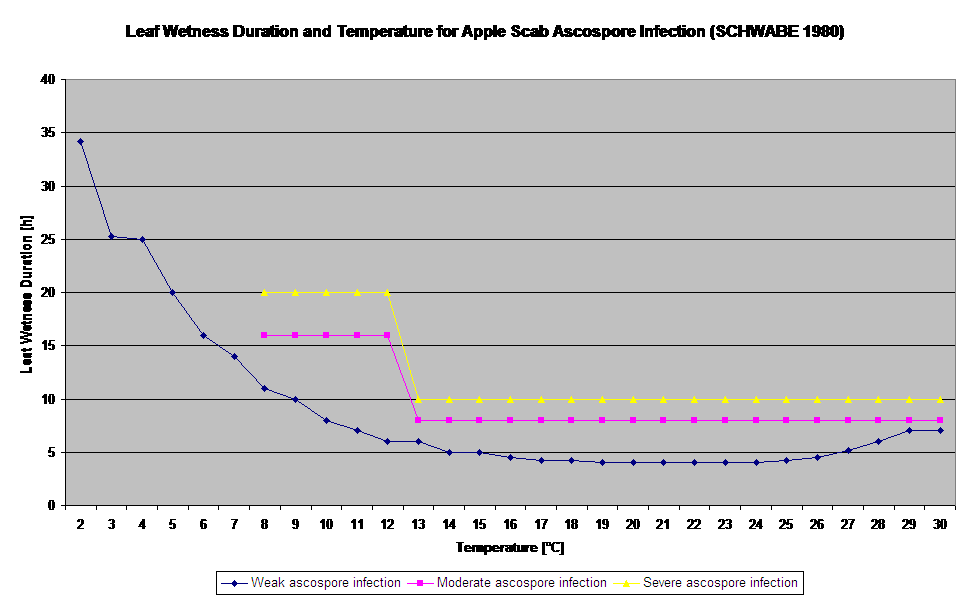
La germination et la pénétration des ascospores de Venturia inaequalis sur les feuilles ou les fruits dépendent des périodes d'humidification des feuilles (feuilles humides ou humidité relative >80% (maintient le niveau d'infection jusqu'à moins de 80% h.r.). et de la température de l'air (supérieure à 2°C). Des températures plus élevées conduisent, dans des conditions humides, à une pénétration plus rapide, tandis qu'à des températures plus basses, des périodes plus longues d'humidité des feuilles sont nécessaires. Cette relation a été publiée pour la première fois par MILLS et LAPLANTE (1945). Notre calcul est effectué sur la base des publications de SCHWABE (1980). SCHWABE a décrit la sévérité des infections de la tavelure en fonction de la température. Ces observations sont en concordance avec nos propres observations.
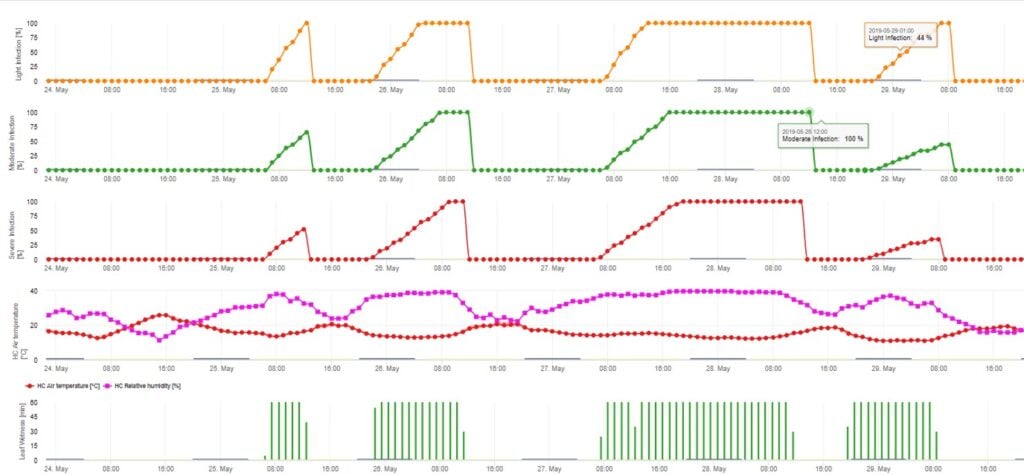
Le modèle d'infection par les ascospores calcule la progression de l'infection, en fonction de l'humidité et de la température des feuilles, pour les infections faibles, modérées et graves. La répartition de la prédiction en trois "classes de gravité" est très traditionnelle pour les modèles de tavelure du pommier. L'évolution de l'infection est affichée sous forme de graphique pour les valeurs horaires. Une infection a déjà eu lieu dès que la courbe de progression d'une infection faible atteint 100%. Une infection modérée ou sévère est terminée si les courbes de progression pour ces classes de gravité atteignent (100%). Souvent, dans la pratique ou dans la réalité, une infection atteint 70% ou 80% tôt le matin et en fonction de la météo actuelle, vous serez en mesure de savoir si cette infection va se terminer ou non. L'affichage des valeurs maximales quotidiennes de la courbe du processus d'infection donne un aperçu des dates d'infection prévues pour la saison.
Utilisation principale du modèle
Pour le marché, il est absolument important d'avoir des pommes exemptes de maladies pour obtenir un bon prix. Par conséquent, les producteurs ont tendance à n'avoir aucune tolérance à la tavelure dans leurs vergers. Le modèle doit être mis en œuvre dans le cadre d'un programme d'application curatif (application de pulvérisations contenant des composés de cyprodinil ou de pyriméthanil avant la floraison) lorsque le modèle indique une infection (qu'elle soit faible ou grave). Dans la période de floraison et de post-floraison, les infections sévères sont les plus fréquentes et toutes les périodes humides ont été couvertes par des pulvérisations protectrices. Par la suite, les tissus végétaux se développent très rapidement, et une protection peut ne fonctionner que pendant 4 jours. Dans ce cas, une pulvérisation curative après une période d'infection incomplète peut également être nécessaire. Mais dans ce cas, il est important de savoir si l'infection a été faible ou sévère et s'il y avait un nombre faible ou élevé d'ascospores prêtes à être infectées.
Littérature
- Schwabe WFS, 1980. Wetting and temperature requirements for apple leaf infection by Venturia inaequalis in South Africa. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Weather favouring apple scab infection in South Africa. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Biologie, épidémiologie et gestion de la tavelure du pommier. St. Paul, MN : Am. Phytopathol. Soc. : 545.
Infections secondaires par conidies
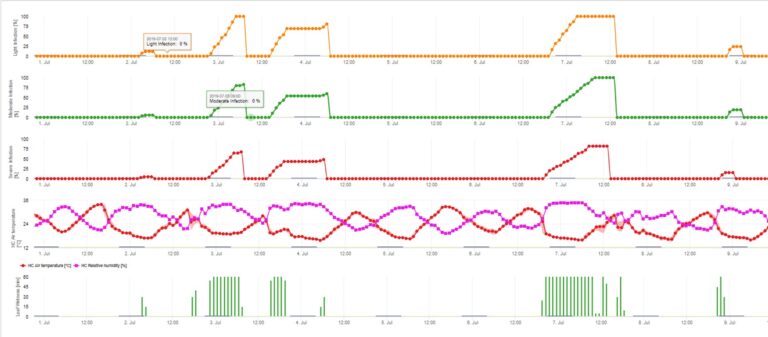
Les infections de la tavelure du pommier par les conidies (asexuées) dépendent des mêmes facteurs que les infections causées par les ascospores. La différence est que les conidies peuvent être libérées à la lumière et à l'obscurité et que les infections conidiennes ont lieu principalement en été.
Selon les régimes de température de l'air, les conidies de Venturia inaequalis ont besoin de périodes d'humidité foliaire plus ou moins longues pour germer et pénétrer dans les feuilles ou les fruits du pommier. Cette relation a été publiée pour la première fois par MILLS et LAPLANTE (1945). Nos calculs sont effectués sur la base des publications de SCHWABE (1980). Alors que Mills et LAPLANTE (1945) ont seulement supposé que les conidies ont besoin d'une certaine période d'humidité foliaire, SCHWABE (1980) a également inclus l'importance de la température dans le processus d'infection.
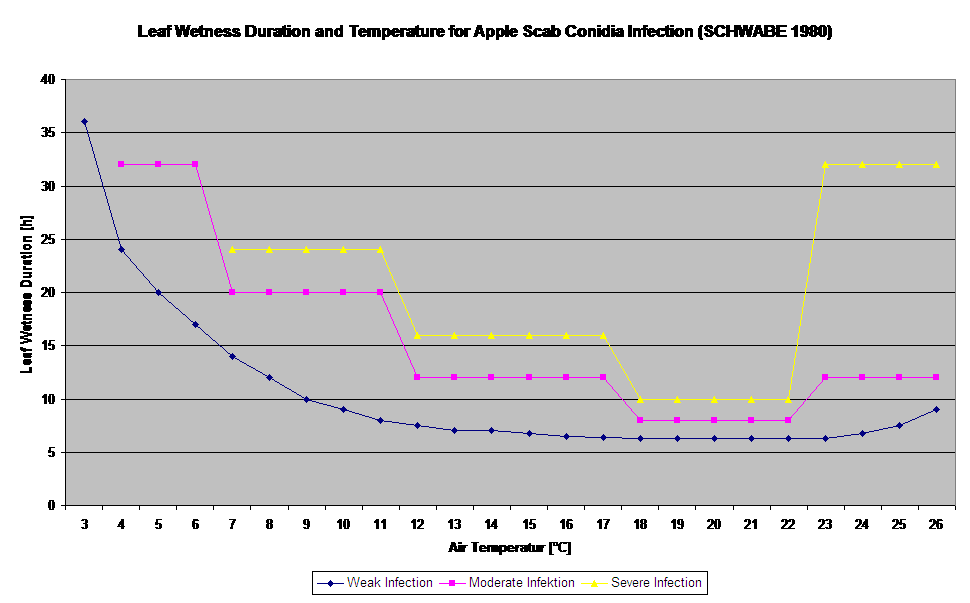
Le modèle d'infection par les conidies calcule la progression de l'infection en fonction de l'humidité et de la température des feuilles pour les infections faibles, modérées et graves. La répartition de la prédiction en "trois classes de gravité" est très classique pour les modèles de tavelure du pommier. La progression de l'infection est affichée sous forme de graphique pour les valeurs horaires. Une infection a déjà eu lieu dès que la courbe de progression d'une infection faible atteint 100%. Une infection modérée ou sévère est terminée si les courbes de progression pour ces classes de gravité atteignent 100%. Souvent, dans la pratique ou la réalité, une infection atteint 70% ou 80% tôt le matin et, en combinaison avec la météo actuelle, vous serez en mesure de savoir si cette infection va se terminer ou non. L'affichage des valeurs maximales quotidiennes de la courbe de progression de l'infection donne une vue d'ensemble des dates d'infection prévues pour la saison.
La littérature :
- Infection à Venturia inaequalis, en indiquant les dates d'infection des ascospores et des conidies (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Biologie, épidémiologie et gestion de la tavelure du pommier. St. Paul, MN : Am. Phytopathol. Soc. : 545.
Présentation graphique du modèle
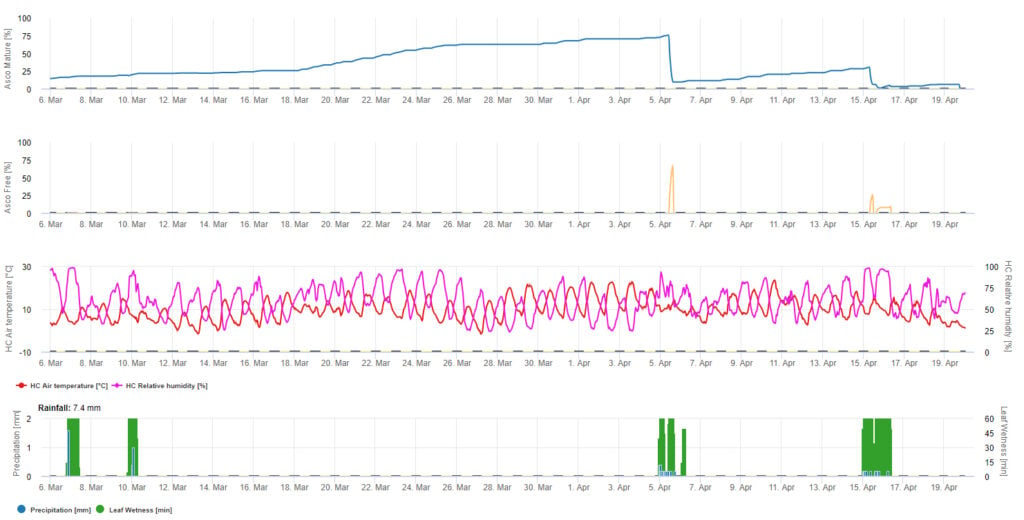
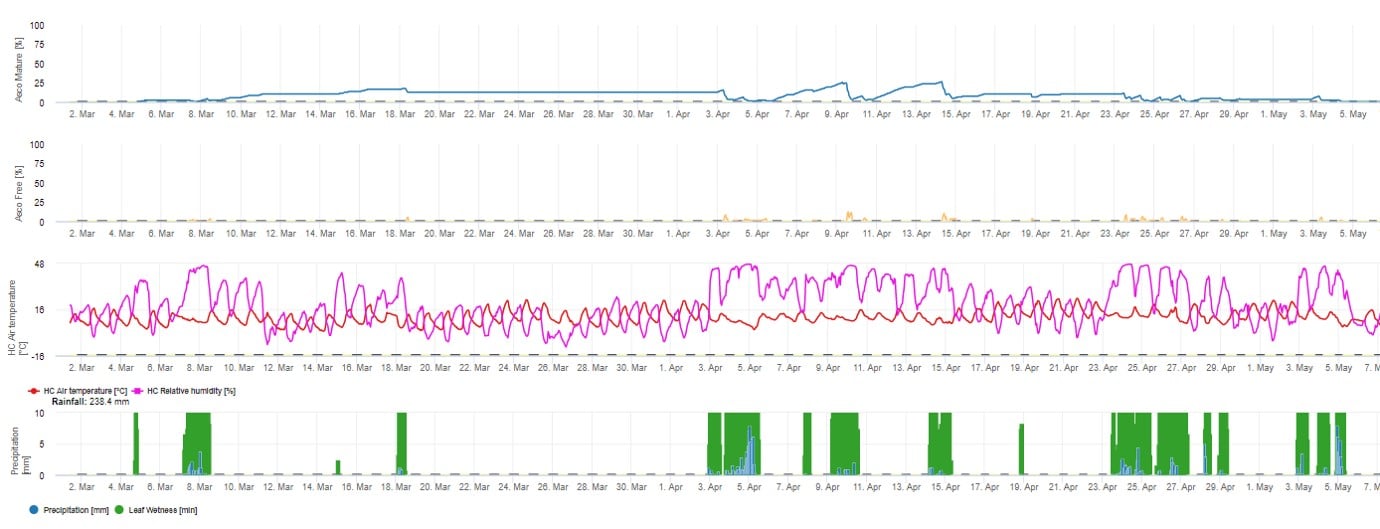
Pour la présentation graphique du modèle de maturation, de formation et de libération des ascospores, des valeurs quotidiennes sont utilisées. Il montre que les périodes d'humidité des feuilles entraînent des libérations d'ascospores et des courbes pour les ascospores matures et libérées. La gravité de la libération et le nombre d'ascospores matures ne peuvent pas être évalués en valeurs absolues. Les valeurs entre 0 et 100% indiquent la quantité relative d'ascospores matures et libérées en fonction des conditions climatiques.
1. La maturation, la formation et l'évacuation de l'ascospore :
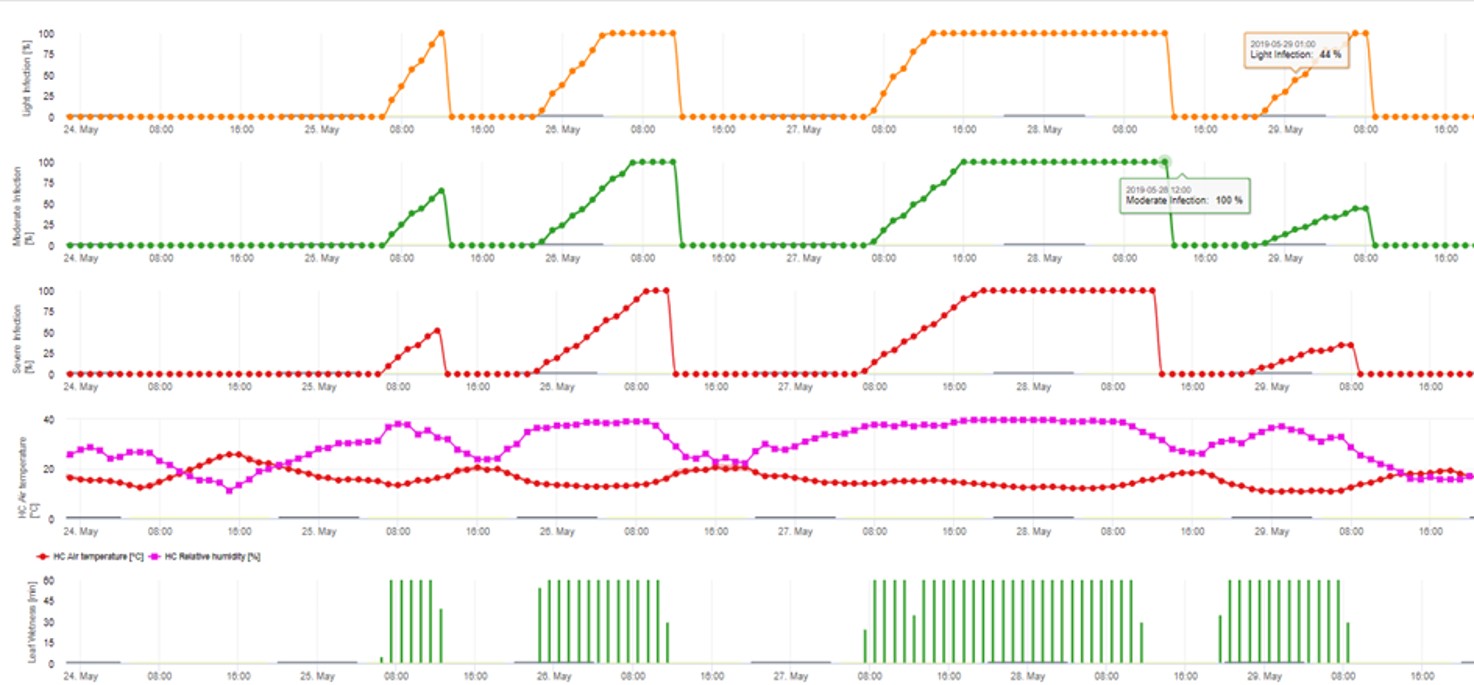
2. Infection par les ascospores montrant trois classes de gravité différentes (faible : orange, modérée : vert, sévère : rouge). Lorsque l'infection par 100% a été atteinte, les conditions optimales pour que l'ascospore infecte les tissus végétaux ont été déterminées. Les mesures de protection des plantes (prophylactiques avant l'infection, curatives après l'infection par 100%) doivent donc être prises en compte :
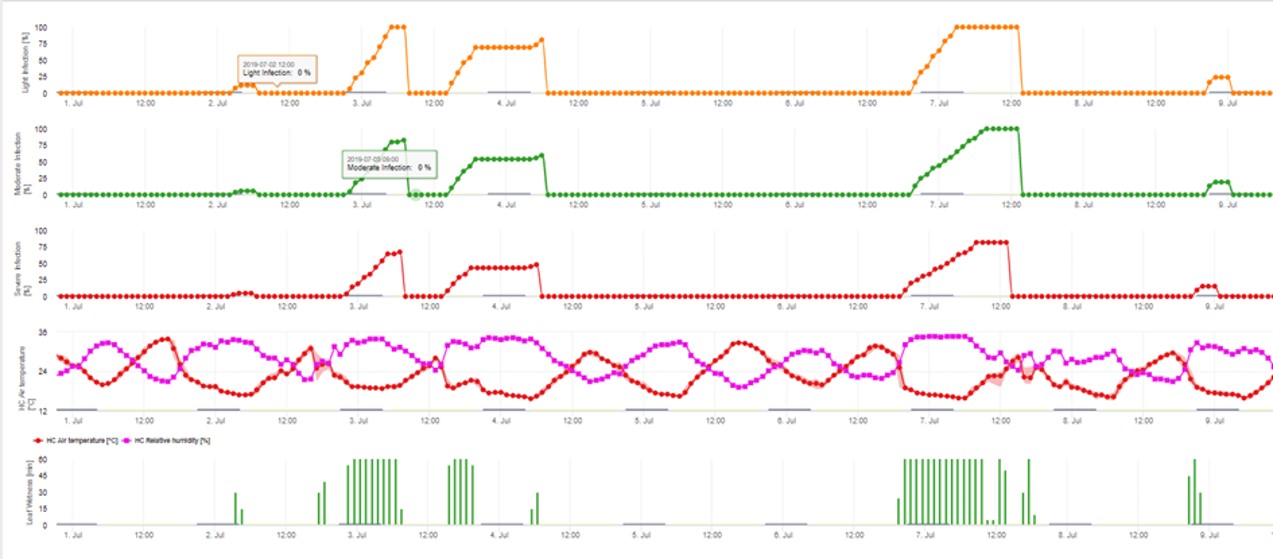
3. L'infection par les conidies apparaît dans trois classes de gravité différentes (faible : orange, modérée : verte, sévère : rouge). Lorsque l'infection par 100% a été atteinte, les conditions optimales pour que l'ascospore infecte les tissus végétaux ont été déterminées. Les mesures de protection des plantes (prophylactiques avant l'infection, curatives après l'infection par 100%) doivent donc être prises en compte :
Les pommes cultivées commercialement doivent être exemptes de tavelure. Les pommes tavelées ne seront vendues que pour la transformation. C'est pourquoi toutes les activités de protection des plantes, tant en culture conventionnelle qu'en culture biologique, ont pour objectif d'obtenir des fruits exempts de tavelure. Les modèles qui montrent la décharge des ascospores de la tavelure du pommier et l'infection par les ascospores et les conidies sont des outils très importants pour atteindre cet objectif.
Deux types fondamentaux de fongicides contre la tavelure du pommier sont utilisés dans les systèmes de culture conventionnels : a) des produits préventifs comme le Captan, le Mancozèbe, le Dithianone et les Strobilurines ou b) des produits curatifs comme le Cyprodinil (Chorus) ou le Pyrimethanil (Scala) ou, pour l'application dans les dernières périodes chaudes de la saison, les fongicides DMI. En fait, la plupart des producteurs suivent une stratégie préventive. Néanmoins, une stratégie préventive pratique n'est pas en mesure de protéger complètement les pommiers, car ceux-ci poussent et développent des fleurs, des fruits et des feuilles. Par conséquent, la pulvérisation préventive ne protège que pendant une période de 4 à 7 jours, en fonction de la croissance réelle de l'arbre. Des intervalles de pulvérisation aussi courts ne sont pas gérables ; c'est pourquoi les producteurs intégreront leur expérience du climat local, des prévisions météorologiques et des modèles de tavelure du pommier dans leur gestion des pulvérisations. Ils programmeront les pulvérisations préventives en fonction de leur expérience et des prévisions météorologiques. Les modèles d'infection de la tavelure du pommier leur indiqueront la date exacte de l'infection (faible, modérée et grave) ainsi que le modèle de décharge des ascospores/conidies et, grâce à leur expérience, ils sont en mesure d'estimer l'importance d'une infection. Cela permet d'agir avec un produit curatif si une infection par la tavelure du pommier s'est produite trop longtemps après la dernière pulvérisation préventive.
Dans la production biologique de pommes, le soufre de chaux s'est avéré être l'agent de contrôle le plus efficace contre la tavelure. Le contrôle optimal peut être atteint si la pulvérisation est effectuée peu avant l'infection ou au début de l'infection. Cela doit être planifié en fonction des prévisions météorologiques. Il arrive que cette période optimale ne soit pas respectée et qu'il faille pulvériser sur les feuilles humides d'une infection presque complète de la tavelure, ce qui donne tout de même une bonne efficacité. L'efficacité du traitement n'en sera pas moins bonne. Les modèles de tavelure aident à décider si une pulvérisation d'émergence dans une infection presque complète est nécessaire.
Symptômes de la brûlure bactérienne
Les chancres hivernants abritant le pathogène du feu bactérien sont souvent clairement visibles sur les troncs et les grandes branches sous la forme de zones d'écorce décolorées légèrement à profondément déprimées, parfois fissurées sur les bords. Le plus grand nombre de chancres, cependant, sont beaucoup plus petits et ne sont pas aussi faciles à distinguer. Ils apparaissent sur les petites branches où des infections de fleurs ou de pousses se sont produites l'année précédente et souvent autour des coupes effectuées pour éliminer les branches atteintes. Comme beaucoup de ces chancres s'établissent plus tard dans la saison, ils ne sont pas souvent fortement déprimés et présentent rarement des fissures de l'écorce sur leurs bords. En outre, ils sont souvent assez petits, moins de 2,5 cm, avec une écorce rougeâtre à pourpre qui peut être couverte de minuscules fructifications de champignons noirs (notamment les Botryosphaeria obtusale pathogène de la pourriture noire du pommier).
Les symptômes de la brûlure de la fleur apparaissent le plus souvent une à deux semaines après la floraison et concernent généralement l'ensemble de la grappe de fleurs, qui se flétrit et meurt, prenant une couleur brune sur le pommier et une couleur assez noire sur le poirier. Lorsque les conditions météorologiques sont favorables au développement du pathogène, des globules d'exsudat bactérien peuvent être observés sur les fleurs. L'éperon portant la grappe de fleurs meurt également et l'infection peut s'étendre à des parties de la branche porteuse et les tuer. Les extrémités des jeunes pousses infectées se flétrissent, formant un symptôme très typique de "crosse de berger". Les pousses plus âgées qui sont infectées après avoir développé une vingtaine de feuilles peuvent ne pas présenter ce symptôme d'enroulement à l'extrémité. Au fur et à mesure que l'infection se propage le long de l'axe de la pousse, les feuilles présentent d'abord des stries sombres dans les nervures médianes, puis se flétrissent et brunissent, tout en restant fermement attachées à la pousse tout au long de la saison. Comme pour les infections des fleurs, le pathogène envahit et tue souvent une partie de la branche qui supporte la pousse infectée. Le premier symptôme sur les pousses d'eau et les pousses qui sont envahies de manière systémique par des chancres actifs proches est le développement d'une décoloration jaune à orange de l'extrémité de la pousse avant le flétrissement. En outre, les pétioles et les nervures centrales des feuilles basales de ces pousses se nécrosent généralement avant celles de l'extrémité de la pousse.
En fonction du cultivar et de son stade de développement au moment de l'infection, une seule fleur ou un seul rameau peut entraîner la mort d'une branche entière, et lorsque l'axe central ou le tronc de l'arbre est envahi, une grande partie de l'arbre peut être tuée en une seule saison. En général, les infections de tout type qui se produisent entre la chute des pétales et le débourrement des bourgeons terminaux entraînent la perte la plus importante de membres et d'arbres. En outre, les arbres fortement structurés ont tendance à subir des pertes de membres moins graves que ceux qui sont formés à des systèmes plus faibles en vue d'une productivité élevée. Lorsque des porte-greffes de pommiers très sensibles (M.26, M.9) sont infectés, une grande partie du tronc du greffon et des principales branches au-dessus du point de greffe ne présentent aucun symptôme, tandis qu'un chancre brun foncé distinct se développe autour du porte-greffe. Lorsque ce chancre du porte-greffe entoure l'arbre, la partie supérieure présente des symptômes de déclin général (couleur médiocre du feuillage, croissance faible) vers le milieu ou la fin de la saison. Dans certains cas, le feuillage des arbres affectés par le chancre du porte-greffe prend une couleur rouge automnale précoce entre fin août et début septembre, qui n'est pas sans rappeler celle souvent associée à la maladie de la pourriture du collet causée par un champignon présent dans le sol. Certains arbres infectés par le porte-greffe peuvent ne pas présenter de symptômes de dépérissement jusqu'au printemps suivant, moment où l'on peut observer des chancres qui s'étendent vers le bas du tronc.
Cycle de la maladie
La bactérie pathogène responsable du feu bactérien passe l'hiver presque exclusivement dans les chancres des branches infectées la saison précédente. Le plus grand nombre de chancres et, par conséquent, ceux qui contribuent le plus à l'inoculum, se trouvent sur les branches d'un diamètre inférieur à 38 mm, en particulier autour des coupes effectuées l'année précédente pour éliminer les branches atteintes par le feu bactérien. Au début du printemps, en réponse aux températures plus chaudes et au développement rapide des bourgeons, les bactéries présentes sur les bords des chancres commencent à se multiplier rapidement et produisent un épais suintement jaunâtre à blanc qui s'élabore sur la surface de l'écorce jusqu'à plusieurs semaines avant la période de floraison. De nombreuses espèces d'insectes (principalement des mouches) sont attirées par l'exsudat et dispersent ensuite les bactéries dans tout le verger. Une fois que les premières fleurs ouvertes sont colonisées par la bactérie, les insectes pollinisateurs déplacent rapidement l'agent pathogène vers d'autres fleurs, ce qui déclenche l'apparition d'une brûlure des fleurs. Ces fleurs colonisées sont sujettes à l'infection dans les minutes qui suivent tout événement humide causé par la pluie ou une forte rosée, lorsque les températures moyennes quotidiennes sont égales ou supérieures à 16 °C et que les pétales des fleurs sont intacts (les réceptacles des fleurs et les jeunes fruits sont résistants après la chute des pétales). Une fois que les infections des fleurs se produisent, les premiers symptômes peuvent être attendus avec l'accumulation d'au moins 57 degrés-jours (DJ) supérieurs à 13 °C, ce qui, en fonction des températures journalières, peut prendre de 5 à 30 jours calendaires.
Avec l'apparition des symptômes de la brûlure de la fleur, le nombre et la distribution des sources d'inoculum dans le verger augmentent considérablement. L'inoculum provenant de ces sources est ensuite propagé par le vent, la pluie et de nombreux insectes visiteurs occasionnels aux extrémités des jeunes pousses, ce qui augmente la probabilité d'une épidémie de brûlure des pousses. Des recherches récentes menées en Pennsylvanie indiquent que l'alimentation des pucerons ne contribue pas à la brûlure des pousses. Des recherches supplémentaires sont nécessaires pour déterminer si les cicadelles jouent un rôle dans l'incidence de la brûlure des pousses. La plupart des infections de l'extrémité des pousses se produisent entre le moment où les pousses ont environ neuf à dix feuilles et l'apparition des bourgeons terminaux, lorsque les sources d'inoculum et les insectes vecteurs sont disponibles et que les températures journalières atteignent en moyenne 16 °C ou plus.
Les années où il n'y a pas d'infection des fleurs, les principales sources d'inoculum pour la phase de dépérissement des pousses sont les chancres hivernants et, en particulier, les jeunes pousses d'eau à proximité de ces chancres, qui sont infectées lorsque les bactéries y pénètrent de manière systémique à partir des marges des chancres. Ces infections systémiques des pousses, appelées "canker blight", sont apparemment initiées à environ 111 JJ de plus de 13 °C après la pointe verte, bien que les symptômes visibles puissent ne pas être apparents avant l'accumulation d'au moins 157 JJ de plus de 13 °C après la pointe verte. En l'absence d'infections des fleurs, le développement des infections des pousses est souvent localisé autour des zones présentant des chancres hivernants.
Brûlure de la fleur de mannequin
Le modèle exige de l'utilisateur qu'il reconnaisse les événements locaux spécifiques et en constante évolution ainsi que les aspects de son verger qui peuvent augmenter ou diminuer le risque de feu bactérien par rapport à d'autres vergers de la région. Le modèle demande à l'utilisateur de supposer qu'il existe un risque d'infection par le feu bactérien chaque fois que des fleurs sont présentes sur les arbres, en particulier pendant la chute des pétales et la période "post-floraison", lorsque des fleurs éparses peuvent subsister sur de nombreuses variétés de pommiers et de poiriers. Il est demandé à l'utilisateur du modèle d'évaluer soigneusement la situation sur son site spécifique et de prendre des mesures de contrôle si des fleurs sont présentes, si les niveaux de risque sont "élevés" ou "extrêmes" et si l'humectation des fleurs est susceptible de se produire au cours des prochaines 24 heures.
Structure du modèle : Températures et humidité : Le processus clé du feu bactérien qui doit être modélisé est le potentiel de croissance bactérienne sur les stigmates des fleurs de pommiers et de poiriers. Cette croissance dépend de la température, de sorte qu'une prédiction fiable du risque d'infection nécessite l'utilisation d'une méthode de mesure qui reflète le plus précisément possible la croissance de la bactérie sur les stigmates. Erwinia amylovora colonies. Le principal désaccord entre les modélisateurs porte sur la manière de procéder.
Le modèle Cougarblight estime le taux de croissance bactérienne en degrés-heures sur la base d'une courbe de croissance spécifique. Cette courbe de croissance est basée sur le taux de croissance de E. amylovora des bactéries lors d'essais en laboratoire. Les valeurs des degrés-heure sont cumulées chaque heure de la journée où les températures sont supérieures à 15 °C. Les valeurs horaires augmentent lorsque les températures passent de 15 °C à 29 °C, diminuent lorsque les températures sont plus élevées et atteignent zéro pour une heure lorsque les températures sont supérieures à 40 °C.
Dans FieldClimate.com, les deux modèles sont présentés dans la même figure. Le modèle Fireblight DIV est basé sur Courgarblight et le modèle Blossomblight est basé sur Maryblight.
Modèle de lutte contre la peste porcine
L'incidence de la brûlure des fleurs est l'un des aspects les plus sporadiques des épidémies de brûlure bactérienne. Le modèle a été construit sur l'hypothèse que l'inoculum est abondant et que, pour qu'un événement d'infection des fleurs se produise, quatre conditions strictes doivent être remplies dans l'ordre. Ces conditions sont les suivantes
1) la fleur doit être ouverte avec les stigmates et les pétales intacts, les stigmates doivent être exposés pour la colonisation, les fleurs dont les pétales sont tombés sont résistantes ;
2) L'accumulation d'au moins 110 °C heures > 18,3 °C au cours des 66 derniers jours > 4,4 °C définit le potentiel d'infection épiphyte de la fleur la plus ancienne et donc la plus colonisée du verger.
3) un événement humide se produisant sous forme de rosée ou de 0,2 mm de pluie ou de 2,5 mm de pluie le jour précédent permet le déplacement des bactéries des stigmates colonisés vers les nectarthodes
4) Une température moyenne journalière de >= 15,6 °C : Cela peut influencer la vitesse à laquelle les bactéries migrent dans les nectarthodes ainsi que la multiplication des bactéries nécessaires à l'établissement des infections.
Présentation graphique des modèles
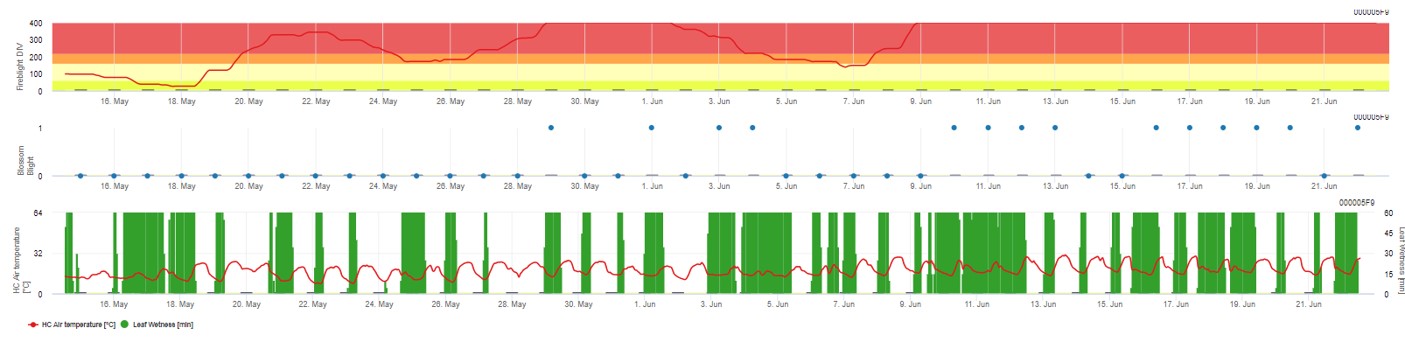
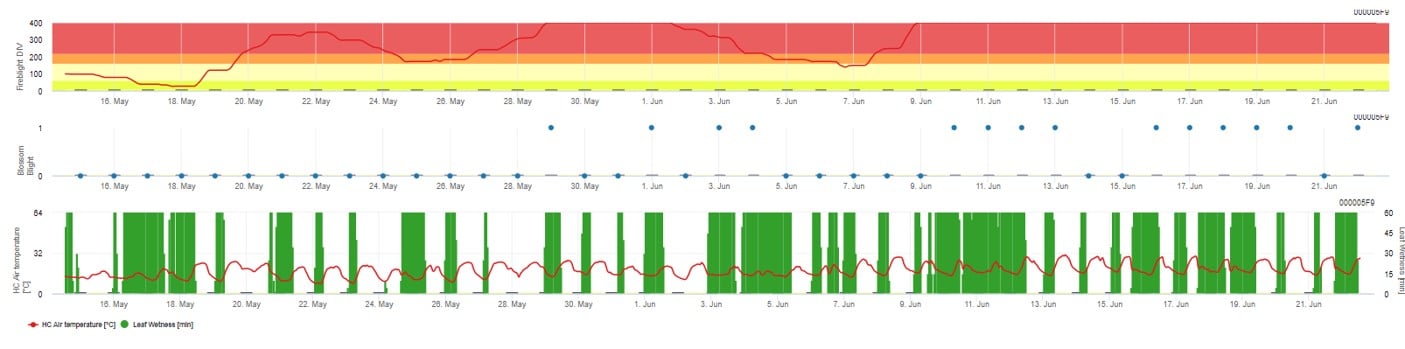
Les deux modèles sont affichés sur la même page. La présentation graphique des modèles de lutte contre le feu bactérien est plus facile à utiliser avec des valeurs agrégées sur une base quotidienne. Le graphique montre les valeurs climatiques liées à cette maladie. Les résultats du Cougar Blight nommés Fire Blight DIV et les résultats du Blossom Blight. Pour interpréter les résultats du Cougar Blight, le graphique est souligné par 5 couleurs différentes. La distribution de ces couleurs est faite en fonction des paramètres de l'historique de la première attaque du verger (voir bouton en haut à droite). Les 5 couleurs indiquent la classe de risque pour les valeurs DIV.
Interprétation des valeurs (en fonction de l'incidence du feu bactérien au cours des dernières années, configuration de base) :
Présence de chancres actifs à proximité
Réglez les paramètres de l'historique du feu bactérien en cliquant sur le bouton bleu "Paramètres" en haut à droite :
Affichage des résultats du modèle Fire Blight par 1) Modell Cougar Blight (appelé Fireblight DIV sur la base de l'historique et des couleurs de fond) et 2) Modell Blossom Blight
| Potentiel de présence de pathogènes | Faible | Modéré | Haut | Extrême |
| Potentiel de présence de pathogènes | Faible | Modéré | Haut | Extrême |
| Pas de feu bactérien au cours des deux dernières années | 0-350 | 350-500 | 500-800 | 800+ |
| Le feu bactérien dans la région pendant deux saisons | 0-300 | 300-500 | 500-750 | 750+ |
| Le feu bactérien dans la région pendant deux saisons | 0-250 | 250-450 | 450-700 | 700+ |
| Feu bactérien dans un verger l'année dernière | 0-200 | 200-350 | 350-500 | 500+ |
Seuil d'intervention :
La lutte est conseillée si une période d'infection à risque élevé ou extrême est détectée. Les facteurs déterminant les dommages sont : le nombre de fleurs, l'âge de l'arbre, la vigueur et la variété,... aussi.
La possibilité d'une infection par le mildiou de la fleur est représentée par une barre allant de 0 à 1 (toutes les conditions sont donc remplies ou non).
L'objectif des modèles de lutte contre le feu bactérien est d'évaluer la probabilité d'une infection par le feu bactérien. Erwinia amyloflora dans le verger.
Le modèle Mary Blight, qui évalue la brûlure de la fleur, indique très bien les situations d'infection ayant un impact économique élevé. C'est pourquoi il est très souvent utilisé pour indiquer l'utilisation d'antibiotiques contre ce pathogène. Le modèle Cougar blight fournit des informations sur le risque d'infection par le feu bactérien en raison des possibilités globales de propagation du pathogène. Sa pondération par l'historique d'un verger est très utile pour nous indiquer à quel point nous devons contrôler les vergers pour détecter les symptômes du feu bactérien, même dans les situations où le feu bactérien Mary n'indique pas d'infection.
Biologie du carpocapse
Le site C. pomonella passe l'hiver sous la forme d'une larve mature enveloppée dans un cocon situé dans des zones protégées de l'arbre (crevasses ou fissures de l'écorce), qui est l'endroit préféré, ou à la base de l'arbre dans la litière du sol. Dans les vergers de pommiers modernes à haute densité, il y a très peu de sites de cocooning sur les arbres en raison de l'écorce lisse. Les nymphes se forment dans le cocon tissé par la larve mature.
Le stade adulte émerge de l'enveloppe nymphale et, après une courte période de durcissement de l'exosquelette, il s'envole vers l'arbre. La plupart des publications indiquent que l'activité des papillons se produit dans le tiers supérieur de la canopée. Les adultes sont actifs au crépuscule pendant quelques heures, avec un seuil d'activité moyen compris entre 13°C et 15°C. L'accouplement a généralement lieu sur l'hôte et la ponte se produit immédiatement après. Il est possible, lors de soirées chaudes, qu'une femelle émerge, s'accouple et ponde des œufs en l'espace de deux jours. A C. pomonella Dans des conditions idéales, le papillon de nuit peut vivre pendant 20 jours, mais dans les champs, il est probable qu'il survive beaucoup moins longtemps, surtout pendant les chaleurs de l'été. Dans des conditions de laboratoire, une femelle pond jusqu'à 100 œufs, mais la fécondité réelle dans le champ est probablement inférieure et la plupart des œufs sont pondus dans les premiers jours suivant l'émergence et l'accouplement.
Les œufs sont pondus individuellement sur la face supérieure de la feuille, sur les rameaux ou sur les fruits. Dans la première génération, les œufs sont pondus uniquement sur les feuilles jusqu'à environ deux semaines après la nouaison des fruits, lorsque le duvet des fruits disparaît. Dans la deuxième génération, environ 65% des œufs sont pondus sur des feuilles, mais celles-ci sont généralement très proches d'un fruit. Les œufs sont d'un blanc crémeux lorsqu'ils sont pondus pour la première fois, ils présentent un anneau rouge à mi-chemin de leur développement et, lorsqu'ils sont proches de l'éclosion, on peut voir la capsule noire de la tête.
Une larve nouvellement éclose erre jusqu'à ce qu'elle trouve un fruit. Elle ronge la peau et se nourrit de la chair de la pomme sous la peau pendant quelques jours. En rongeant la peau et en se nourrissant sous la peau, la larve rejette des excréments à l'extérieur de la zone d'entrée. Un anneau rouge entoure souvent l'entrée, en particulier sur les jeunes fruits. Une fois que la larve a mué pour passer au stade suivant, elle se dirige vers le noyau où elle se nourrit de graines jusqu'à ce qu'elle arrive à maturité. Lorsqu'elle est presque arrivée à maturité, la larve creuse un tunnel entre le trognon et l'extérieur de la pomme et bouche le trou avec des excréments et de la soie. Lorsqu'elle est prête à tisser un cocon, la larve quitte le fruit, en traversant l'arbre ou en se laissant tomber au sol sur un fil de soie, et trouve un endroit protégé.
Modèle pour le carpocapse
Notre modèle est basé sur l'accumulation des températures effectives entre >= 10°C et <=32°C. Dans l'hémisphère nord, cette accumulation commence au premier janvier. Dans l'hémisphère sud, il commence au premier juillet. Nous cumulons chaque mesure dans son équivalent pour la journée (voir graphique ci-dessous). Les seuils suivants sont utilisés pour le développement des insectes :
*Génération complète= 650
| *De la ponte à l'éclosion de l'œuf | = 88 |
| *Premier stade larvaire | = 60 |
| *Deuxième stade larvaire | = 65 |
| *Troisième stade larvaire | = 55 |
| *stade de la larve | = 45 |
| *Cinquième stade larvaire et nymphe | = 335 |
Le graphique produit par le modèle montre la première apparition et la durée du stade larvaire 1 à 5 et la présence d'adultes dans 5 niveaux.
| *Niveau 1 : | Activité de vol, sans ponte |
| *Niveau 2 : | Activité de vol avec possibilité de ponte. |
| *Niveau 3 : | Activité de vol avec un peu de ponte. |
| *Niveau 4 : | Activité de vol avec une ponte raisonnable à forte. |
| *Niveau 5 : | Activité de vol avec une forte ponte. |
Présentation des stades adultes du carpocapse à 5 niveaux différents (premier graphique) ainsi que des degrés-jours cumulés (ligne rouge, deuxième graphique) pour le développement des adultes et des stades larvaires représentés en bleu (troisième graphique).
Plusieurs espèces de pucerons sont présentes dans les vergers de pommiers. Le puceron vert du pommier, le puceron de l'avoine ou puceron du pommier et le puceron rose du pommier. Dans les jardins privés, le puceron rose du pommier a une certaine importance. Ce puceron provoque des changements hormonaux importants à l'extrémité des pousses en cas d'infestation. Les feuilles se recroquevillent et la pousse se raccourcit. Plus tard, les feuilles jaunissent et meurent. Ce puceron affecte surtout les jeunes pousses. Le puceron rose du pommier peut déjà causer de graves dégâts en petite quantité. Par contre, le puceron vert du pommier et le puceron de l'herbe à pommier peuvent être présents en plus grande quantité sans causer de dommages économiques. Le puceron du pommier doit être considéré comme n'ayant aucune importance économique.
Puceron rose du pommier
Il s'agit d'une ancienne espèce européenne introduite aux États-Unis vers 1870. Elle est devenue un ravageur important des pommes vers la fin du 19e siècle. Pour prospérer, cette espèce doit disposer d'une abondance de sa plante hôte estivale, le plantain à feuilles étroites. Il existe un parallélisme remarquable entre l'introduction et la propagation du puceron rose du pommier et du plantain à feuilles étroites. Ce puceron est présent dans toutes les régions fruitières du monde.
Les étapes de la vie
Œuf : L'œuf est ovale et légèrement aplati du côté de l'écorce. Sa longueur varie de 0,49 à 0,56 mm. Lors de la première ponte, il est jaune vif et recouvert d'une substance glutineuse qui durcit avec l'âge. La couleur passe progressivement au jaune verdâtre et enfin à un noir de jais brillant. Le temps nécessaire à ce changement de couleur varie, dans des conditions extérieures normales, d'environ neuf jours à plus de deux semaines.
Nymphe : Les individus qui éclosent des œufs sont tous des femelles vivipares sans ailes. Il y a cinq stades. Le dernier est la mère souche mature qui, peu après le quatrième stade, commence à produire des jeunes vivants parthénogénétiquement. Elle produit en moyenne cinq à six jeunes par jour. Les nymphes de la deuxième génération, qui sont toutes des femelles, atteignent la maturité en deux à trois semaines. La grande majorité des nymphes commencent à se reproduire sur le pommier, mais quelques-unes peuvent développer des ailes et migrer vers le plantain.
La troisième génération est produite en juin et début juillet. La majorité de cette génération développe des ailes et migre vers le plantain à feuilles étroites. Certaines saisons, les femelles sans ailes de la troisième génération produisent une quatrième génération sur le pommier. Ces dernières années, on a observé dans certaines régions que les populations nuisibles de pucerons roses du pommier persistaient dans les vergers jusqu'au milieu de l'été ou plus tard. On ne sait pas si ce changement est dû au processus de sélection, à des changements d'habitudes ou à l'absence de besoin d'un hôte alternatif.
Adulte : La couleur de l'adulte varie considérablement. La couleur générale est brun rosé, avec une dominante rosée due à une couverture poudreuse. Certains des adultes les plus âgés sont pourpres, tandis que les plus jeunes sont résolument rose rougeâtre.
Gamme d'hôtes
Le pommier est l'hôte préféré, mais le puceron se nourrit également de poirier et d'aubépine. Les variétés de pommes Cortland, Golden Delicious, Rhode Island Greening et Ida Red sont particulièrement sensibles. Le puceron est présent dans toutes les régions fruitières des États-Unis et du Canada.
Blessures ou dommages : L'alimentation du puceron rose du pommier provoque souvent l'enroulement des feuilles de pommier, à partir de la chute des pétales. Ces feuilles peuvent ensuite devenir rouge vif. L'alimentation des feuilles autour des grappes de fruits entraîne souvent la formation de grappes, le rabougrissement et la malformation des fruits en développement. Ces anomalies s'aggravent au fur et à mesure que les fruits se développent et peuvent finalement les rendre invendables.
Les grandes populations de pucerons peuvent produire de grandes quantités de miellat comme déchet de la sève dont ils se nourrissent. Le miellat excrété sur les fruits servira de terrain de croissance à la moisissure fuligineuse, qui affectera la finition de la pomme. Les toxines contenues dans la salive des pucerons servent également de "goutte d'eau", empêchant l'abscission des fruits (séparation naturelle de l'arbre) au moment de la récolte normale.
Histoire de la vie : Le puceron passe l'hiver au stade de l'œuf. L'éclosion a lieu au début du printemps, environ une semaine à 10 jours plus tard que le puceron des céréales et à peu près en même temps que le puceron du pommier. Les œufs éclosent lorsque les bourgeons commencent à s'ouvrir au printemps, sur une période de deux semaines. Dès l'éclosion, les jeunes recherchent les bourgeons du pommier qui s'ouvrent ; ils semblent préférer les bourgeons à fruits. Ils se nourrissent à l'extérieur des bourgeons de feuilles et des grappes de bourgeons de fruits jusqu'à ce que les feuilles commencent à se déployer. Ils descendent ensuite à l'intérieur des grappes et commencent à sucer la sève des tiges et des fruits nouvellement formés.
Leur alimentation provoque l'enroulement des feuilles, ce qui protège les pucerons des pulvérisations et de certains ennemis. L'enroulement sévère du feuillage provoqué par cette espèce est probablement l'élément le plus caractéristique de son travail. Une seule mère de tige située sur la face inférieure d'une feuille, près de la nervure centrale, provoque un pliage serré de la feuille. Il suffit de quelques mères de tige pour provoquer un enroulement sévère de toutes les feuilles entourant un bourgeon de fleur qui s'ouvre, offrant ainsi une protection idéale aux pucerons qui se développent rapidement. Les mères de tige atteignent leur maturité lorsque les pommiers commencent à fleurir.
Les mères tiges matures sont très inactives. Elles s'installent, se nourrissent et produisent des petits à un rythme rapide. Lorsqu'elles sont distribuées, elles retirent rapidement leur bec des tissus végétaux et cherchent un autre endroit où continuer. Les mères de tige arrivent à maturité environ deux semaines après l'éclosion. La durée de cette période dépend en grande partie des conditions météorologiques. La production de jeunes commence généralement deux ou trois jours après la dernière mue et se poursuit sans interruption pendant plus d'un mois.
La production totale d'une femelle est en moyenne de 185. Normalement, la période de reproduction s'étend de début mai à juin. En général, la période maximale d'activité reproductive se situe autour de la dernière semaine de mai et de la première semaine de juin, lorsque les jeunes fruits commencent à se former et à entamer une croissance active. Le puceron rose du pommier s'attaque rarement aux jeunes pousses à croissance rapide. Il se limite au feuillage, aux pédoncules floraux et aux jeunes fruits.
L'une des caractéristiques de cette espèce est le rassemblement des jeunes autour de la mère. Chaque mère de tige individuelle ou groupe de mères a des centaines de jeunes massés autour d'elle. Les feuilles infestées peuvent rapidement être recouvertes - dans certains cas, par plus d'une couche de pucerons. Cette habitude de se rassembler tue rapidement les feuilles infestées et provoque la migration forcée des pucerons. Les jeunes se déplacent activement et précipitamment, apparemment désireux de trouver un endroit propice à l'alimentation. Pendant cette période, on les trouve souvent rassemblés sur les fruits en formation ou attaquant le nouveau feuillage succulent qui se déploie.
La deuxième génération a besoin de quatre à 40 jours pour atteindre la maturité et produire des jeunes. La majorité des individus de la deuxième génération sont des femelles sans ailes. La production totale moyenne de chaque individu de la deuxième génération est d'environ 119. Les habitudes et les activités de la troisième génération ne diffèrent pas de celles de la deuxième. Les pucerons se rassemblent en très grand nombre sur la face inférieure du feuillage, provoquant un enroulement important. Ils s'attaquent également à la nouaison et aux fruits en développement, provoquant des lésions caractéristiques.
La majorité de cette troisième génération acquiert des ailes et migre vers le plantain à feuilles étroites, la plante hôte estivale. Une quatrième génération peut être produite. Toutes ces générations produisent des ailes et migrent vers le plantain à feuilles étroites. Après la dernière mue, les adultes ailés sont très tendres et inactifs. Ils restent sécrétés dans les feuilles enroulées pendant deux ou trois jours avant d'entreprendre leur vol migratoire. Juste avant de s'envoler, ils deviennent très actifs et nerveux, courant dans tous les sens ou bougeant leurs ailes de haut en bas dans l'attente de leur vol.
Puceron vert du pommier
Le puceron vert du pommier, Aphis pomi (DeGeer), est largement répandu dans toutes les régions productrices de pommes. Dans l'hémisphère nord, il apparaît pour la première fois dans les vergers à la fin du mois de mai ou au début du mois de juin. L'insecte suce la sève des feuilles sur les pousses d'eau et les pousses terminales succulentes. Le puceron vert du pommier se trouve généralement près des nervures principales, sur la face inférieure de la feuille. Le puceron vert du pommier s'attaque également au poirier, à l'aubépine, au cognassier, à la pomme de terre et à la spirée.
Description du puceron vert du pommier
Les œufs du puceron vert du pommier sont ovales et d'un noir brillant. Les nymphes et les adultes sont vert foncé ou clair. Le puceron vert du pommier a une tête verte, un abdomen uniformément vert ou vert jaunâtre, des pattes noires et de longues cornicules noires - les "tuyaux de la queue" (à l'extrémité arrière de l'insecte). Les femelles adultes sont vertes foncées à noires et ailées. Pendant l'été, ces femelles, ou alates, donnent naissance à des jeunes vivants parthénogénétiques.
Biologie du puceron vert du pommier
Le puceron vert du pommier passe l'hiver sous forme d'œufs sur les drageons, à la base des bourgeons et sur les cicatrices foliaires des pousses terminales. Les œufs éclosent généralement lorsque les bourgeons éclatent et que les premières feuilles se déploient. Les nymphes commencent immédiatement à se nourrir des feuilles en développement. Elles sont d'abord présentes sur la pousse terminale et se déplacent ensuite vers les feuilles plus âgées de la grappe. Les pucerons femelles adultes produisent des femelles sans ailes en deux semaines, chaque femelle étant capable de produire 50 à 100 descendants vivants. Les jeunes pucerons se développent en sept à dix jours. Les populations de pucerons verts du pommier se développent lentement au début du printemps (floraison, pétales, automne), puis plus rapidement lorsque les températures moyennes quotidiennes augmentent. Des pucerons ailés, ou alates, et des pucerons sans ailes sont produits pendant l'été. Les pucerons verts du pommier sont les plus nombreux en juillet et début août. Au début de l'infestation, il y a plus de nymphes que d'adultes ailés. Le pourcentage d'alates augmente au fur et à mesure que la population s'accroît. Les colonies de pucerons verts du pommier peuvent doubler en une semaine si les conditions sont favorables. Si les pucerons n'ont pas de nouvelles pousses succulentes et qu'ils doivent se nourrir de feuilles plus âgées, le nombre de jeunes produits peut chuter de 50%. Si les températures journalières sont de 30-32°C et plus, les femelles ne se reproduisent pas bien. Lorsque les températures sont élevées pendant plusieurs jours, les pucerons meurent. Plusieurs générations de pucerons verts du pommier se développent au cours de l'été et les formes ailées se dispersent dans le verger. Parfois, le puceron vert du pommier et le puceron rose du pommier, Dysaphis plantaginea Pars, ont des colonies sur la même feuille. Le puceron rose du pommier est rose ou violet. À l'automne, les mâles et les femelles sans ailes se développent et s'accouplent, et les femelles pondent des œufs pour l'hiver. Le plus grand nombre d'œufs se trouve à 15-20 cm de l'extrémité des rameaux. Les œufs se trouvent rarement sur les grandes branches d'échafaudage ou sur les troncs des pommiers.
Dégâts
Le puceron vert du pommier suce la sève de la feuille. Les fortes infestations réduisent la vigueur et la croissance des pousses. Ce phénomène est particulièrement préoccupant dans les pépinières et les jeunes vergers non productifs. L'insecte peut réduire la taille des bourgeons et la longueur des entre-nœuds, et provoquer l'enroulement des feuilles. L'alimentation des pucerons peut également stimuler la croissance des branches latérales, ce qui peut affecter la forme de l'arbre. L'enroulement des feuilles et les terminaisons affaiblies sont susceptibles d'être endommagées par l'hiver. Le miellat produit par les pucerons peut s'écouler sur les fruits et permettre le développement de champignons fuligineux. Ces champignons ternissent les fruits et réduisent leur valeur marchande. Lorsque les infestations sont importantes, le puceron vert du pommier peut se nourrir de pommes immatures et provoquer un roussissement.
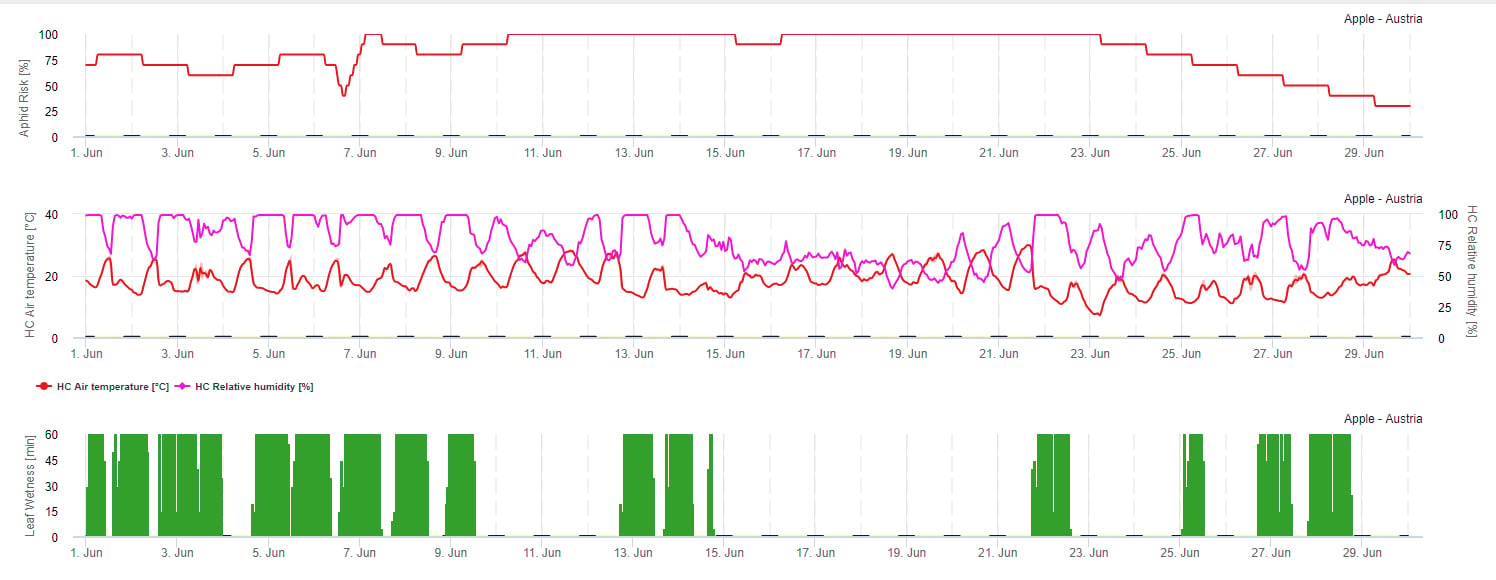
Modèle de risque de propagation
La propagation des pucerons est principalement influencée par la température. Plus le nombre d'heures est proche de la température optimale (20-32°C) et plus la source de nourriture est disponible, plus le taux de propagation est élevé. Les œufs et les jeunes stades sont sensibles à l'air sec. Une faible humidité relative peut également nuire aux populations de pucerons (en dessous de 30% r.h.). Outre les insecticides, les pucerons sont contrôlés par des prédateurs et des champignons parasites. Les champignons parasites dépendent de l'humidité. La plupart de ces espèces comme Beauvaria bassiana sont favorisés par l'humidité libre.
Nous augmentons le risque lorsque la température optimale se situe entre 20 et 32°C et l'humidité relative entre 30 et 95% r. h. et nous le diminuons lorsqu'elle est trop humide (humidité des feuilles, pluie, ou humidité relative supérieure à 95%), trop froide (températures inférieures à 20°C) ou trop chaude (températures supérieures à 32°C), nuits humides (humidité des feuilles pendant les heures de la nuit).