La roya de la soja, en concreto la roya asiática de la soja, es una enfermedad grave de la soja causada por el hongo patógeno Phakopsora pachyrhizi. Distribución de la enfermedad: Asia y Australia, y más recientemente desde África, Norteamérica y Sudamérica. La enfermedad se observó por primera vez en Japón en 1902. El patógeno se distribuyó por Asia y Australia a través de América. La roya de la soja no puede invernar en zonas con temperaturas bajo cero, pero puede propagarse rápidamente por el viento a distancias tan grandes, su desarrollo puede ser tan explosivo y puede causar una pérdida tan rápida de hojas que actualmente es una de las enfermedades más temidas en las zonas productoras de soja del mundo.
Síntomas y signos
Los primeros síntomas de la roya de la soja causada por Phakopsora pachyrhizi comienzan como manchas muy pequeñas de color marrón o rojo ladrillo en las hojas. En el campo, estas manchas suelen aparecer en la parte inferior de la copa en el momento de la floración o después, aunque las plántulas pueden infectarse en determinadas circunstancias. A menudo, las primeras lesiones aparecen en la base del folíolo, cerca del pecíolo y de las nervaduras de la hoja. Esta parte del foliolo probablemente retiene el rocío durante más tiempo, haciendo que las condiciones sean más favorables para la infección. Las lesiones siguen siendo pequeñas (2-5 mm de diámetro), pero aumentan en número a medida que avanza la enfermedad. En estas lesiones se forman pústulas, llamadas uredinias, principalmente en la superficie inferior de la hoja, que pueden producir muchas urediniosporas.
Las pústulas elevadas pueden verse a simple vista, sobre todo cuando esporulan. Aunque las lesiones son pequeñas, cada una de ellas suele tener varias pústulas (uredinia) . Las lesiones pueden estar completamente cubiertas de urediniosporas cuando las pústulas están activas. Las urediniosporas de la roya de la soja son de color marrón amarillento pálido a incoloras, con una ornamentación superficial equinulada (espinas cortas). Esta coloración es diferente de la de muchos otros patógenos de la roya, cuyas esporas suelen ser de color marrón rojizo (color óxido). Germinación de P. pachyrhizi urediniosporas se produce a través de un poro ecuatorial (central), produciendo un tubo germinativo que termina en un apresorio, que el hongo utiliza para penetrar en el huésped directamente o a través de un estoma.
A medida que se forman más y más lesiones en un foliolo, la zona afectada empieza a amarillear y, finalmente, el foliolo se cae de la planta. Aunque la roya de la soja suele empezar en la parte inferior de la planta, progresa rápidamente hacia arriba hasta que todas las hojas presentan algún nivel de enfermedad. Las plantas gravemente enfermas pueden quedar completamente defoliadas. La pérdida de tejido foliar efectivo se traduce en una reducción del rendimiento, tanto por la menor cantidad de semillas como por su menor tamaño. Se han registrado pérdidas de rendimiento de hasta 30 a 80%, pero la cuantía de la pérdida depende del momento en que comienza la enfermedad y de la rapidez con que progresa. Además de en las hojas, la roya de la soja también puede aparecer en pecíolos, tallos e incluso cotiledones, pero la mayoría de las lesiones de roya se producen en las hojas.
Las lesiones pueden ser de color bronceado o marrón rojizo. Las lesiones bronceadas tienen muchas pústulas que producen numerosas urediniosporas. Las lesiones de color marrón rojizo, que se consideran una reacción de resistencia moderada, sólo tienen unas pocas pústulas que producen unas pocas urediniosporas. Como se comentará en la sección Manejo de la enfermedad, este tipo de lesión depende de la cepa del patógeno, y puede aparecer en la misma hoja con lesiones de color bronceado, o las lesiones de color bronceado pueden volverse de color rojo-marrón con la edad. Los síntomas y signos en otros hospedantes, como el kudzu, son similares, aunque el tamaño de las lesiones puede diferir. A medida que las pústulas envejecen, pueden volverse negras. Esto se debe a la formación de una capa de teliosporas en las pústulas, convirtiendo las pústulas de uredinia en telia.
Las teliosporas tienen dos funciones: la supervivencia del hongo en ausencia de un huésped vivo (sobreestacionamiento) y la reproducción sexual. Las gruesas paredes de las teliosporas protegen al hongo del medio ambiente y del ataque de otros organismos. En las royas, las teliosporas germinan formando un basidio y cuatro basidiosporas durante las cuales se produce la recombinación sexual. La germinación de P. pachyrhizi teliosporas sólo se ha observado en el laboratorio y no parece contribuir de forma significativa a la perpetuación de esta enfermedad en el campo.
Biología patógena
Hay dos hongos estrechamente relacionados que causan roya en la soja: Phakopsora pachyrhizique a veces se denomina roya asiática o australiana de la soja, pero que ahora también está presente en el hemisferio occidental, y P. meibomiae, el llamado patógeno de la roya de la soja del Nuevo Mundo, que sólo se encuentra en el hemisferio occidental. Salvo por algunas características menores, los dos hongos parecen morfológicamente idénticos, pero P. pachyrhizi es mucho más agresivo en la soja que P. meibomiae. Hasta la fecha, P. meibomiae no se ha documentado que cause pérdidas significativas de rendimiento en América Central y del Sur. Las dos especies pueden distinguirse utilizando protocolos de análisis de ADN.
Al igual que otras royas, los patógenos de la roya de la soja son parásitos obligados que necesitan un hospedador vivo para crecer y reproducirse. Pueden sobrevivir lejos del huésped como urediniosporas sólo durante unos días en condiciones naturales. Hasta donde sabemos, ambos patógenos de la roya de la soja sólo producen dos tipos de esporas: urediniosporas y teliosporas (Figura 15). Esto contrasta con otras royas, que pueden tener hasta cinco estadios de esporas (por ejemplo, la roya del tallo del trigo). En el caso de la roya de la soja, como en la mayoría de las royas, el estadio uredinial es el que se repite. Esto significa que las urediniosporas pueden infectar al mismo huésped en el que se produjeron (soja) durante la misma temporada. Las epidemias pueden desarrollarse rápidamente a partir de unas pocas pústulas porque las pústulas productoras de esporas se producen en tan sólo 7 a 10 días después de la infección, y cada pústula puede producir cientos de urediniosporas. Las teliosporas se producen en lesiones antiguas, pero no parecen germinar en la naturaleza, y no se conoce ningún hospedador alternativo, ni aecia o espermogonia.
Sin la germinación de las teliosporas, la reproducción sexual no puede tener lugar. La falta de reproducción sexual debería limitar la variabilidad del hongo de la roya, pero sin embargo existe una variabilidad sustancial en P. pachyrhizi con respecto a la virulencia. Esto ha limitado el uso de genes únicos para la resistencia en la soja, porque en poco tiempo surgen nuevos aislados del patógeno que superan el gen de resistencia. Se desconoce cómo se origina esta variabilidad en P. pachyrhizi. Roya del trigo, Puccinia striiformistiene un ciclo de vida similar al de P. pachyrhizi sin estadio telial funcional y, por tanto, sin reproducción sexual, pero tiene muchas razas. Puede ser que cada gen de resistencia sea tan específico que una sola mutación en el gen correcto del hongo le permita ser virulento en huéspedes con el nuevo gen de resistencia.
Epidemiología
Las epidemias de roya de la soja comienzan con la llegada de inóculo aéreo (urediniosporas). Este patógeno es único entre las royas porque tiene muchos hospedadores alternativos, que pueden servir como fuentes de inóculo. Los hospedadores alternativos son otras plantas que pueden infectarse con el mismo patógeno, pero que no son necesarias para completar el ciclo de vida del patógeno. Los hospedantes alternativos no deben confundirse con el hospedante alternativo, que es una planta distinta del hospedante principal, necesaria para que un patógeno complete su ciclo vital. En zonas libres de heladas, como Sudamérica, Centroamérica, la cuenca del Caribe, el sur de Texas y Florida, la fuente de inóculo podría estar cerca en plantas de soja voluntarias, kudzu o algún otro hospedante alternativo. En zonas donde se producen heladas, como el Medio Oeste de Estados Unidos, el inóculo debe ser transportado por aire desde fuentes de hibernación que pueden estar a cientos de kilómetros de distancia.
La reintroducción de patógenos obligados en una región lejana se produce con otras enfermedades, como la roya del trigo y los mildiu, por ejemplo el moho azul del tabaco. Dado que las esporas de P. pachyrhizi son sensibles a la radiación ultravioleta, el desplazamiento a larga distancia de estas esporas de roya se produce probablemente en sistemas de tormentas en los que las nubes protegen a las esporas del sol. Una vez que las esporas viables han aterrizado en la superficie de la hoja de un huésped adecuado, la infección y el posterior desarrollo de la epidemia dependen de las condiciones ambientales. Por lo general, la infección se produce cuando las hojas están húmedas y la temperatura oscila entre 8°C y 28°C, con un óptimo de 16°C a 28°C. A 25°C, se produce cierta infección en la superficie de las hojas. A 25°C, se produce cierta infección en tan sólo 6 horas de humectación de las hojas, pero lo óptimo son 12 horas. Tras la infección, pueden aparecer lesiones y pústulas con urediniosporas en 7 u 8 días, y el siguiente ciclo de infección está listo para comenzar.
Este corto ciclo vital significa que, en las condiciones adecuadas, las epidemias de roya de la soja pueden pasar rápidamente de niveles casi indetectables a niveles muy altos. Las epidemias de roya de la soja pueden progresar desde niveles por debajo de lo detectable hasta la defoliación en un mes. Puede parecer que las epidemias progresan incluso más rápido que eso, porque las primeras infecciones se producen en la parte inferior de la canopia y son difíciles de encontrar. Además del medio ambiente, la edad de la planta afecta a las epidemias de roya de la soja. Normalmente, las lesiones de roya no se encuentran en la soja hasta la floración, a menos que haya altos niveles de inóculo al principio de la temporada. Esto puede deberse a una mayor susceptibilidad de las plantas a la roya a medida que el huésped entra en las etapas reproductivas, puede deberse a que en las partes bajas de la canopia las esporas están más protegidas de la radiación UV, o puede deberse a que las condiciones en la canopia se vuelven más húmedas a medida que ésta se cierra. En cualquier caso, las lesiones pueden formarse en cualquier fase de crecimiento, pero los mayores incrementos de la enfermedad no se producen hasta después de la floración.
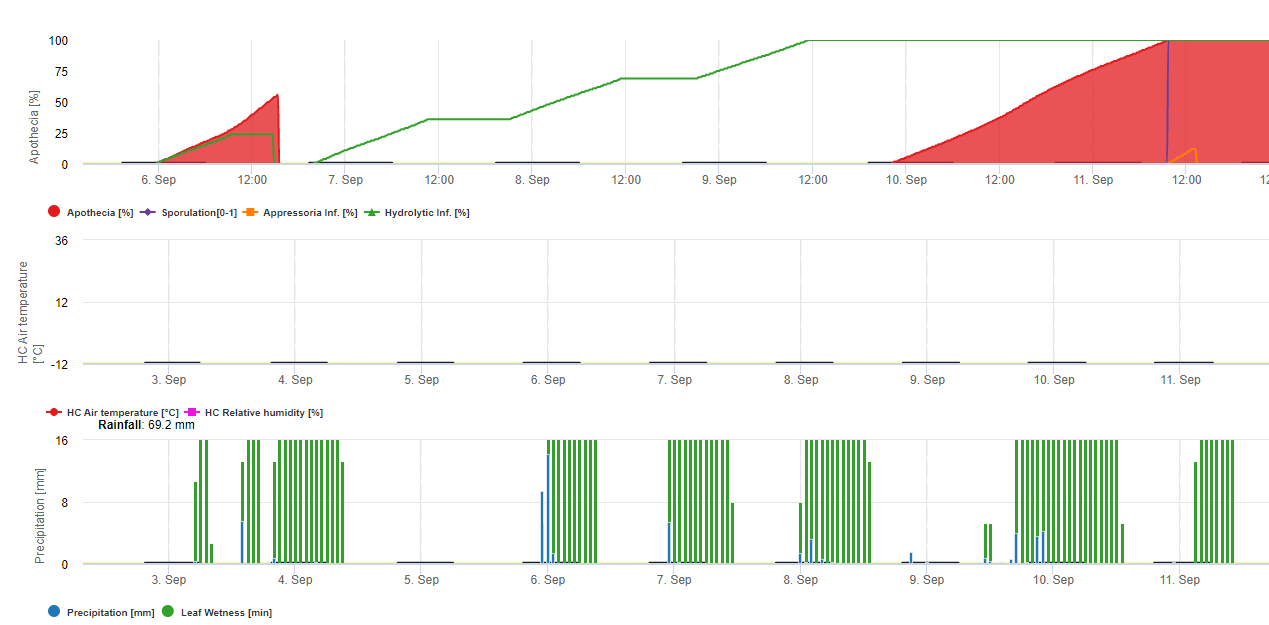
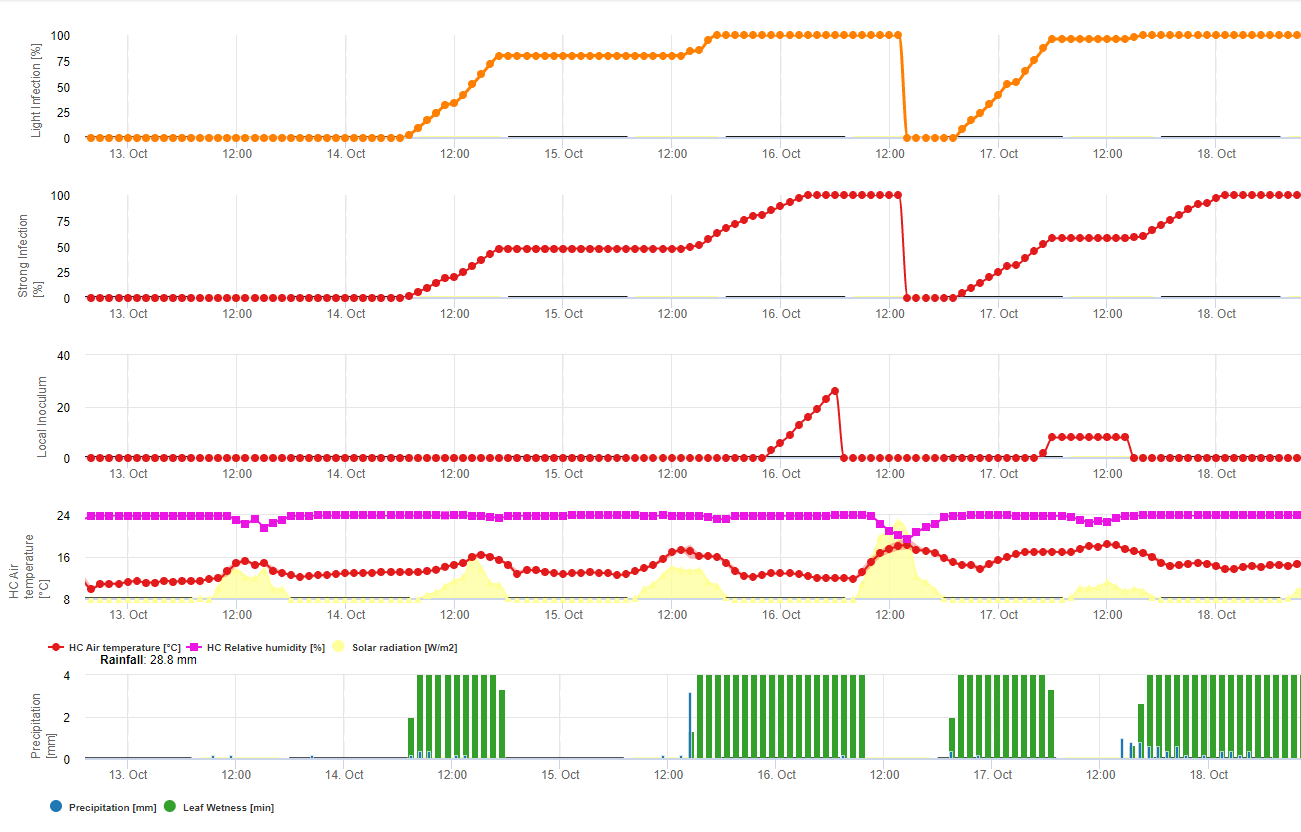
En FieldClimate se describen y calculan dos modelos para la roya de la soja:
Modelo 1: Los principales factores para el cálculo de la infección y la severidad de la roya de la soja son la humedad de la hoja y la temperatura (tenga cuidado la infección tiene que ser 100% para que los índices de severidad (del 1-3) empiecen a calcularse) Las hojas tienen que estar húmedas y la temperatura tiene que estar entre 8 y 28°C - la infección empieza a calcularse (la suma de 7200 °minutos es 100%). Si la infección es 100% y en dependencia de la temperatura y la duración de la humedad de la hoja se empiezan a calcular las clases de severidad de 1- 3.
Modelo 2: Aquí se incluyen muchos más factores en el cálculo de la roya de la soja: Temperatura, humedad de la hoja, humedad relativa, precipitaciones y radiación solar (las hifas del hongo pueden entrar en los estomas durante el día).
Cuando la infección de la roya de la soja alcanza 100% se empieza a calcular la severidad. Si ambos (infección y severidad son 100%) se predice la aparición (¿establecimiento?) de inóculo local.
Literatura:
- Ivancovich, A. y Botta, G. 2002. La roya de la soja en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L., y Royer, M. H. 1989. Effects of duration, frequency, and temperature of leaf wetness periods on soybean rust. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte y A.P. Dias. Conociendo el riesgo de la roya de la soja por comparación con Brasil. Departamento de Patología Vegetal, Universidad Estatal de Iowa.
- Yang, X. B. 1995. Evaluación y gestión del riesgo de roya de la soja. Proceeding of the soybean rust workshop, 9-11 de agosto de 1995. J. B.Sinclair y G. L. Hartman, eds. Laboratorio Nacional de Investigación de la Soja, Urbana, IL
La podredumbre del tallo por esclerotinia o moho blanco de la soja es una enfermedad causada por el hongo Sclerotinia sclerotiorum. La enfermedad puede causar importantes reducciones en la producción de semillas o incluso destruir por completo un cultivo cuando la soja se planta en suelo infestado y hay una cubierta vegetal densa con periodos prolongados de tiempo húmedo. Las pérdidas de rendimiento suelen producirse cuando la incidencia de la enfermedad es del 15 % o superior. En la mayoría de los años en Dakota del Norte, la podredumbre del tallo por Sclerotinia es sólo un problema de menor a moderado para los productores, y la enfermedad rara vez se observa durante los años de sequía. El clima húmedo es un factor importante en el desarrollo de la enfermedad. Ha sido un problema persistente en la soja de regadío. Además de la reducción de la producción de semillas, la enfermedad también provoca la reducción de la calidad de las semillas y la contaminación de las semillas con los esclerocios negros del hongo. La contaminación de las semillas puede ser un problema grave para las semillas exportadas, ya que puede dar lugar al rechazo del lote de semillas en los puertos de entrada extranjeros. Además, los esclerocios devueltos al suelo pueden afectar a otros cultivos de la rotación. Los agricultores pueden controlar la podredumbre del tallo por esclerotinia si conocen el patógeno y el ciclo de la enfermedad.
Síntomas
Los síntomas no suelen observarse hasta que el dosel del cultivo entre las hileras se ha cerrado, creando un microclima húmedo. El marchitamiento de las hojas seguido de la muerte de las plantas suelen ser los primeros síntomas observados. Una inspección minuciosa bajo el dosel de la planta revelará un crecimiento micelial algodonoso y blanco (hilos de hongo) en tallos, hojas o vainas. Las lesiones se desarrollan en los tallos principales y las ramas laterales. Con el tiempo, las lesiones ciñen los tallos y las partes de la planta situadas por encima mueren. Los tallos aparecen blanqueados y a veces astillados por el avanzado deterioro. A partir del micelio blanco que crece en el tejido vegetal se forman esclerocios grandes y negros de diferentes formas y tamaños. Los esclerocios también se forman en la médula del tallo y tienen una forma cilíndrica característica. Las semillas de las vainas enfermas suelen estar arrugadas y pueden estar infectadas por el hongo o sustituidas por esclerocios negros. La semilla suele contaminarse con esclerocios cuando se cosechan plantas infectadas.
Biología patógena
Este hongo tiene una amplia gama de hospedadores, con más de 370 especies de plantas, y causa enfermedades en una gran variedad de cultivos, como girasol, judía seca, colza, patata, alfalfa, trigo sarraceno, altramuz, mostaza, pataca, cártamo, lenteja, lino, guisantes forrajeros y muchas hortalizas. En Dakota del Norte es raro encontrar este patógeno causando daños graves en algunos de estos cultivos como el lino y la patata. También hay muchas malas hierbas de hoja ancha comunes como el saúco de los pantanos, la hierba de los corderos, la cerdilla, el cardo canadiense y la mostaza silvestre. El hongo que causa el moho blanco en la soja es el mismo que causa la enfermedad del moho blanco o Sclerotinia en el girasol, las judías secas, la colza y otros cultivos. Sclerotinia sclerotiorum pasa el invierno principalmente en forma de esclerocios en el suelo. Los esclerocios germinan y forman pequeños hongos de color canela a marrón llamados apotecios (de entre un octavo y un cuarto de pulgada de diámetro). Estos producen esporas denominadas ascosporas que inician la enfermedad en la soja y otros cultivos susceptibles.
Ciclo de la enfermedad
La humedad y la floración son factores críticos en el desarrollo de la enfermedad. Las enfermedades no suelen aparecer antes de que se cierre el dosel del cultivo, porque un dosel denso favorece las temperaturas frescas y un microclima húmedo alrededor de los tallos y mantiene una elevada humedad del suelo tras la lluvia o el riego. El inicio de la enfermedad también está estrechamente relacionado con la floración. Tras siete a catorce días de elevada humedad del suelo, los esclerocios germinan a varios centímetros de profundidad y forman apotecios en forma de hongo. Un solo esclerocio puede producir varios apotecios. Los apotecios expulsarán por la fuerza sus ascosporas al aire, donde serán transportadas por las corrientes de aire hasta las plantas de soja. La fuente más importante de ascosporas son los apotecios producidos dentro del campo, pero las ascosporas pueden llegar desde campos adyacentes o cercanos. Un apotecio puede producir enormes cantidades de ascosporas durante varios días. Las ascosporas sobreviven durante breves periodos en el tejido vegetal, pero no pasan el invierno. Las ascosporas necesitan una película de agua y una base alimenticia, como tejidos florales muertos o senescentes, para germinar y crecer antes de infectar la planta. El tejido floral es la base alimenticia más importante para iniciar las infecciones. A menudo, las infecciones se inician en las axilas de los tallos, donde los tejidos florales senescentes han caído y se han alojado. Las infecciones también pueden producirse a través de heridas causadas por el granizo u otras lesiones. Una película de agua en la superficie de la planta favorece el desarrollo de las lesiones y aumenta el daño tisular. El desarrollo inicial de la enfermedad requiere generalmente más de 40 horas de humedad continua de la superficie de la planta, pero una vez iniciada la enfermedad, periodos más cortos de humedad permitirán el desarrollo de las lesiones. Esta es la razón por la que la enfermedad se asocia con largos periodos de tiempo nublado, húmedo y lluvioso. A medida que la superficie de la planta se seca, el progreso de la enfermedad se ralentiza. Las temperaturas frescas entre 59 y 75 grados Fahrenheit favorecen el desarrollo de la enfermedad. Cuanto mayor sea la densidad de la cubierta vegetal, más favorables serán las condiciones ambientales para la enfermedad. Por lo tanto, la soja de siembra sólida y los cultivos alojados son más propensos al desarrollo de la enfermedad. Se formarán esclerocios a medida que el micelio crezca dentro y sobre los tejidos de la planta. Estos esclerocios no germinarán para formar más apotecios durante la temporada, sino que serán devueltos al suelo durante las operaciones de cosecha y laboreo y pasarán el invierno para convertirse en inóculo (fuente de hongo infeccioso) para un futuro cultivo susceptible. Los esclerocios son estructuras muy resistentes y sobreviven durante largos periodos en el suelo.
Fuente: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
En FieldClimate el modelo para Sclerotinia se calcula en función de los periodos de lluvia, la humedad relativa y la temperatura, así como la humedad de las hojas. Durante largos periodos húmedos se recomienda una infección a través del establecimiento de un apresorio por el hongo patógeno. Otra forma de infección se denomina "infección hidrolítica". Esta forma se basa en la liberación de enzimas hidrolíticas que degradan secuencialmente la cutícula de la planta, las láminas medias y las paredes celulares primarias y secundarias y la planta entera. Modelo descrito en detalle en esta página.