





Pera modelos de enfermedad
Acumulación de lluvia
La lluvia intensa arrastrará los pesticidas que queden en las hojas de las vides u otras plantas. La resistencia a la lluvia de los fungicidas modernos ha mejorado mucho desde 1980. Actualmente podemos esperar que los fungicidas más modernos resistan hasta 30 mm de lluvia si han tenido la oportunidad de secarse en la hoja. Si la lluvia comenzara inmediatamente después de la pulverización o durante la misma, la resistencia a la lluvia podría reducirse considerablemente.
Con las formulaciones antiguas de fungicidas de contacto debemos esperar una dureza de la lluvia inferior a 12 mm. Como estábamos acostumbrados en los años 70. Para mojar las hojas en un viñedo se necesitan aproximadamente 2 mm de lluvia. Por lo tanto en este mdoule acumulamos solamente lluvias con son más grandes que 2m m dentro de un período de la mojadura de la hoja. Esto significa que podría haber en total 6 mm de lluvia durante un solo día, pero este módulo no acumula nada de eso porque las hojas se han secado de nuevo antes de que llovieran 2 mm.
La lluvia se acumula durante 3, 5 y 7 días. Durante un periodo más largo, el crecimiento de la planta es mucho más importante para el efecto de los fungicidas de contacto que la resistencia a la lluvia de los compuestos.
Fuego bacteriano
Fuente:P.W. Steiner, T. van der Zwet y A. R. Biggs
El fuego bacteriano es una enfermedad bacteriana destructiva de manzanas y peras que mata flores, brotes, ramas y, a veces, árboles enteros. La enfermedad se da en casi todas las zonas productoras de manzanas de clima moderado a cálido del mundo. Aunque los brotes suelen ser muy erráticos, causando graves pérdidas en algunos huertos en algunos años y poco o ningún daño significativo en otros. Esta ocurrencia errática se atribuye a las diferencias en la disponibilidad de inóculo de hibernación, los requisitos específicos que rigen la infección, las variaciones en las condiciones climáticas locales específicas y el estado de desarrollo de los cultivares disponibles. El potencial destructivo y la naturaleza esporádica del fuego bacteriano, junto con el hecho de que las epidemias a menudo se desarrollan en varias fases diferentes, hacen que esta enfermedad sea difícil y costosa de controlar.
Síntomas
Los chancros invernales, que albergan el patógeno del fuego bacteriano, son a menudo claramente visibles en troncos y ramas grandes como zonas de corteza descolorida de ligera a profundamente deprimidas, que a veces están agrietadas en los márgenes. El mayor número de cancros, sin embargo, son mucho más pequeños y no se distinguen tan fácilmente. Aparecen en ramas pequeñas donde se produjeron infecciones de flores o brotes el año anterior y, a menudo, alrededor de los cortes realizados para eliminar las ramas afectadas. Dado que muchos de estos cancros se establecen más tarde en la temporada, no suelen estar muy deprimidos y rara vez muestran grietas en la corteza en sus márgenes. Además, suelen ser bastante pequeños, de menos de 2,5 cm, con una corteza entre rojiza y púrpura que puede estar cubierta de minúsculos cuerpos fructíferos de hongos negros (sobre todo Botryosphaeria obtusael patógeno de la podredumbre negra de la manzana).
Los síntomas del tizón de la flor aparecen con mayor frecuencia una o dos semanas después de la floración y suelen afectar a todo el racimo de flores, que se marchitan y mueren, volviéndose marrones en el manzano y bastante negras en el peral. Cuando el clima es favorable para el desarrollo del patógeno, pueden verse glóbulos de exudado bacteriano en las flores. El espolón que soporta el racimo de flores también muere y la infección puede extenderse y matar partes de la rama de soporte. Las puntas de los brotes jóvenes infectados se marchitan, formando un síntoma muy típico de "cayado de pastor". Los brotes más viejos que se infectan después de haber desarrollado unas 20 hojas pueden no mostrar este síntoma de rizado en la punta. A medida que la infección se propaga por el eje del brote, las hojas muestran primero vetas oscuras en las venas medias, luego se marchitan y se vuelven marrones, permaneciendo fuertemente unidas al brote durante toda la temporada. Al igual que con las infecciones de flores, el patógeno a menudo invade y mata una parte de la rama que soporta el brote infectado. El primer síntoma en brotes de agua y brotes que son invadidos sistémicamente desde cancros activos cercanos es el desarrollo de una decoloración amarilla a naranja de la punta del brote antes de que se produzca el marchitamiento. Además, los pecíolos y las venas medias de las hojas basales de estos brotes suelen necrosarse antes que las de la punta del brote.
Dependiendo del cultivar y de su estado de desarrollo en el momento en que se produce la infección, la infección de una sola flor o brote puede provocar la muerte de una rama entera, y cuando se invade la rama central o el tronco del árbol, puede morir una parte importante del árbol en una sola temporada. En general, las infecciones de cualquier tipo que se producen entre la caída de los pétalos y la brotación terminal suelen provocar la mayor pérdida de ramas y árboles. Además, los árboles fuertemente estructurados tienden a sufrir una pérdida de ramas menos severa que aquellos formados con sistemas más débiles para una alta productividad. Cuando se infectan portainjertos de manzano muy susceptibles (M.26, M.9), gran parte del tronco de la púa y las ramas principales por encima de la unión del injerto no suelen presentar síntomas, mientras que se desarrolla un chancro marrón oscuro alrededor del portainjerto. A medida que este cancro rodea el árbol, la parte superior muestra síntomas de decaimiento general (mal color del follaje, crecimiento débil) a mediados o finales de la temporada. En algunos casos, el follaje de los árboles afectados por el chancro del portainjerto desarrolla un color rojo otoñal a finales de agosto o principios de septiembre, parecido al que se asocia a menudo con la enfermedad de la podredumbre del cuello causada por un hongo del suelo. Algunos árboles con infecciones de portainjertos pueden no mostrar síntomas de decaimiento hasta la primavera siguiente, momento en el que pueden observarse cancros que se extienden hacia arriba en la parte inferior del tronco.
Ciclo de la enfermedad
El patógeno bacteriano causante del fuego bacteriano inverna casi exclusivamente en los chancros de las ramas infectadas la temporada anterior. El mayor número de cancros y, por lo tanto, los más importantes en la aportación de inóculo, se producen en ramas de menos de 38 mm de diámetro, especialmente alrededor de los cortes realizados el año anterior para eliminar las ramas afectadas. A principios de la primavera, en respuesta a temperaturas más cálidas y rápido desarrollo de las yemasEn la primavera, las bacterias de los márgenes del cancro comienzan a multiplicarse rápidamente y producen un exudado espeso de color amarillento a blanco que se elabora sobre la superficie de la corteza hasta varias semanas antes del periodo de floración. Muchas especies de insectos (sobre todo moscas) se sienten atraídas por el exudado y, posteriormente, dispersan las bacterias por todo el huerto. Una vez que las primeras flores abiertas están colonizadas por la bacteria, los insectos polinizadores trasladan rápidamente el patógeno a otras flores, iniciando más tizón de la flor. Estas flores colonizadas están sujetas a la infección en cuestión de minutos después de cualquier evento de humectación causada por lluvia o rocío intenso cuando el las temperaturas medias diarias son iguales o superiores a 16 °C mientras los pétalos de la flor están intactos (los receptáculos florales y los frutos jóvenes son resistentes tras la caída de los pétalos). Una vez que se producen las infecciones de la flor, se pueden esperar síntomas tempranos con la acumulación de at al menos 57 grados-día (DD) superiores a 13 °C que, dependiendo de las temperaturas diarias, puede requerir de 5 a 30 días naturales.
Con la aparición de los síntomas del tizón de la flor, el número y la distribución de fuentes de inóculo en el huerto aumentan en gran medida. El inóculo procedente de estas fuentes se propaga además por viento, lluvia, y muchos visitantes casuales de insectos a las puntas de los brotes jóvenes, aumentando la probabilidad de un brote de tizón de los brotes. Investigaciones recientes llevadas a cabo en Pensilvania indican que la alimentación de los áfidos no contribuye al tizón de los brotes. Se necesita más investigación para determinar si las chicharritas desempeñan o no un papel en la incidencia del tizón de los brotes. La mayoría de las infecciones de las puntas de los brotes se producen entre el momento en que los brotes tienen entre nueve y diez hojas y la brotación terminal, cuando se dispone de fuentes de inóculo e insectos vectores y las temperaturas diarias alcanzan una media de 16 °C o más.
En los años en que no se producen infecciones en flor, la fuentes primarias de inóculo para la fase del tizón de los brotes son los cancros invernales y, en particular, los brotes de agua jóvenes cercanos a estos cancros, que se infectan a medida que las bacterias se desplazan hacia ellos sistémicamente desde los márgenes del cancro. Tales infecciones sistémicas de los brotes, denominadas tizón del cancro, se inician aparentemente alrededor de 111 DD mayores de 13 °C después de la punta verde, aunque los síntomas visibles pueden no ser aparentes hasta la acumulación de al menos 157 DD mayores de 13 °C después de la punta verde. En ausencia de infecciones de la flor, el desarrollo de las infecciones del tizón de los brotes se localiza a menudo alrededor de las zonas con cancros invernantes.
Aunque los tejidos maduros de los brotes y las extremidades suelen ser resistentes a la infección por E. amylovoraLas lesiones causadas por el granizo, las heladas tardías de -2 °C o menos y los fuertes vientos que dañan el follaje pueden crear una situación de tizón traumático en la que los mecanismos de defensa normales de los tejidos maduros se ven afectados y se producen infecciones. Se sabe que se producen casos de tizón traumático incluso en cultivares normalmente resistentes como 'Delicious'.
El fuego bacteriano del portainjerto, otra fase del fuego bacteriano, se ha reconocido recientemente y se asocia principalmente con los portainjertos M.26, M.9 y Mark, que son muy susceptibles. En estos árboles, sólo unas pocas infecciones de flores o brotes en el cultivar de púa pueden suministrar bacterias que luego se desplazan sistémicamente al portainjerto, donde a menudo, pero no siempre, se desarrolla un cancro que acaba por ceñir el árbol. Los árboles afectados por el tizón del portainjerto suelen mostrar síntomas de decaimiento y muerte prematura a mediados o finales de la temporada, pero pueden no ser evidentes hasta la primavera siguiente.
(c) Dr. Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2007
Modelo del tizón de la pera
Síntomas
El modelo requiere que el usuario reconozca específico y siempre cambiante eventos locales y los aspectos de su huerto que pueden aumentar o disminuir el riesgo de niebla del peral y del manzano en relación con otros huertos de la región. El modelo requiere que el usuario asuma que existe riesgo de infección de niebla del peral y del manzano siempre que haya flores en los árboles, especialmente durante el periodo de floración. caída de los pétalos y periodo "post-floración, cuando pueden quedar flores dispersas en muchas variedades de manzanos y perales. Se pide al usuario del modelo que evalúe cuidadosamente la situación en su lugar específico y que inicie medidas de control si hay flores presentes, los niveles de riesgo son "Altos" o "Extremos", y es probable que la floración se produzca en algún momento durante las próximas 24 horas.
Estructura del modelo
Temperaturas y humedad: El proceso clave del fuego bacteriano que debe modelarse es el potencial de crecimiento bacteriano en los estigmas de las flores del manzano y el peral. Este crecimiento depende de la temperatura, por lo que una predicción fiable del riesgo de infección requiere el uso de un método de medición que refleje con la mayor exactitud el crecimiento de las colonias de Erwinia amylovora. El principal desacuerdo entre los modelos es cómo debe hacerse.
Salida del modelo de niebla del peral y del manzano en FieldClimate
El modelo Cougar blight estima la tasa de crecimiento bacteriano con las horas-grado basándose en una curva específica de tasa de crecimiento. Esta curva de crecimiento se basa en la tasa de crecimiento de E. amylovora bacterias en pruebas de laboratorio. Los valores de grados hora se acumulan cada hora del día en que las temperaturas superan los 15 °C. Los valores horarios aumentan a medida que las temperaturas suben de 15 °C a 29 °C, disminuyen a temperaturas más altas y llegan a cero en cualquier hora con temperaturas superiores a 40 °C.
Modelo de la plaga de la flor del peral
- La flor debe estar abierta con estigmas y pétalos intactos, los estigmas tienen que estar expuestos para la colonización, las flores en caída de pétalos son resistentes;
- la acumulación de al menos 110 °C horas > 18,3°C en los últimos 44°C días > 4,4°C define el potencial de infección epifítica de la flor abierta más antigua y, por tanto, más colonizada del huerto
- Un acontecimiento húmedo en forma de rocío o 0,2 mm de lluvia o 2,5 mm de lluvia el día anterior permite el desplazamiento de las bacterias de los estigmas colonizados a los nectártodos.
- una temperatura media diaria de >= 15,6°C: Esto puede influir en la velocidad a la que las bacterias migran a los nectártodos, así como en la multiplicación de bacterias necesaria para establecer infecciones.
Cuando se cumplen estos cuatro requisitos mínimos en la secuencia mostrada, se producen las infecciones y se puede esperar que aparezcan los primeros síntomas tempranos del tizón de la flor con la acumulación de 57 °C días adicionales > 12,7 °C. Esto puede ocurrir entre 5 y 30 días después de la infección. Cuando las condiciones del huerto son inferiores a estos requisitos mínimos, se producen pocos o ningún síntoma y no se desarrolla ninguna epidemia significativa. (STEINER P.W. 1996)
Presentación gráfica del fuego bacteriano en las peras
En FieldClimate, los dos modelos de fuego bacteriano se muestran en un gráfico. El modelo Cougar Blight se denomina Fire Blight DIV y el modelo Maryblight se denomina Blossomblight. Para interpretar los resultados del Cougar Blight, el gráfico se subraya con 5 colores diferentes. La distribución de estos colores se realiza en función de los ajustes del primer historial de tizón del huerto. Los 5 colores indican la clase de riesgo de los valores DIV.
La oportunidad de una Infección en Flor se indica mediante una barra que va de 0 - 1 (se cumplen o no las condiciones) en el mismo gráfico. Las configuraciones sobre el historial del huerto no están integradas en este modelo. ¡Cada vez que se calcula una barra con infección de floración en FieldClimate.com es una infección de fuego bacteriano!
Uso práctico El fuego bacteriano en las peras
El objetivo de los modelos de niebla del peral y del manzano es evaluar la probabilidad de infecciones por Erwinia amyloflora en el huerto.
En Modelo Mary Blight que se está evaluando para el tizón de la flor indica muy bien situaciones de infección de gran impacto económico. Por este motivo, se utiliza con bastante frecuencia para indicar el uso de antibióticos contra este patógeno.
Tizón del puma proporciona información sobre el riesgo de infección por el fuego bacteriano debido a las posibilidades de propagación del patógeno. Su ponderación realizada por el historial de un huerto es muy útil para indicarnos con qué cuidado tenemos que comprobar los huertos en busca de síntomas de fuego bacteriano, incluso en situaciones en las que el Mary Blight no indicará una infección.
(c) Dr. Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2008
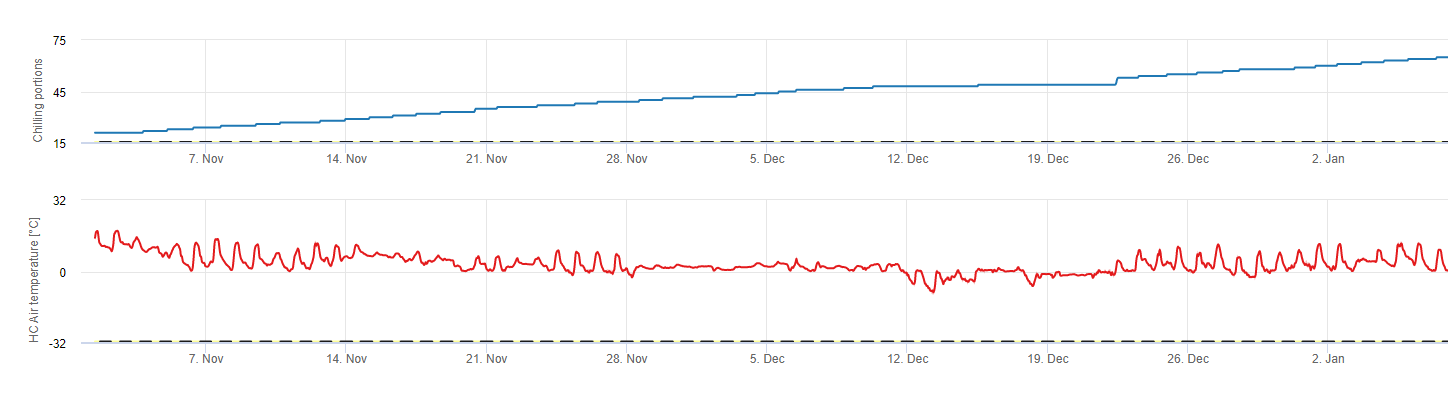
Porciones frías
Escalofriante
Los frutales de hueso desarrollan sus yemas vegetativas y fructíferas en verano y, a medida que se acerca el invierno, las yemas ya desarrolladas entran en reposo en respuesta a la menor duración del día y a las temperaturas más bajas. Esta fase de latencia protege a las yemas del frío. Una vez que las yemas han entrado en letargo, toleran temperaturas muy por debajo del punto de congelación y no crecen en respuesta a los periodos cálidos de mediados de invierno. Estas yemas permanecen latentes hasta que han acumulado suficientes unidades de frío (UC). Cuando suficiente frío se acumulan, los brotes están listos para crecer en respuesta a las temperaturas cálidas. Mientras haya suficientes CU, los botones florales y foliares se desarrollan con normalidad. Si los brotes no reciben temperaturas de enfriamiento suficientes durante el invierno para liberar completamente el letargo, los árboles desarrollarán uno o más de los síntomas fisiológicos asociados a un enfriamiento insuficiente: 1) retraso en la foliación, 2) reducción del cuajado y aumento del abotonamiento y, 3) reducción de la calidad del fruto.
Síntomas de enfriamiento insuficientes
Foliación retardada:
Un síntoma clásico de enfriamiento insuficiente es el retraso en la foliación. Un árbol puede tener un pequeño mechón de hojas cerca de las puntas de los tallos y estar desprovisto de hojas de 12 a 20 pulgadas por debajo de las puntas. Los brotes inferiores se romperán con el tiempo, pero la foliación completa se retrasa significativamente, la fructificación se reduce y el árbol se debilita. Además, la fuerte succión de las partes bajas del árbol causa problemas de gestión, y el desarrollo normal de las yemas frutales del año siguiente puede verse afectado.
Fructificación y abotonamiento reducidos:
La floración, en respuesta a un enfriamiento insuficiente, suele seguir el patrón observado en el desarrollo de las hojas. La floración se retrasa, se prolonga y, debido a anomalías en el desarrollo del pistilo y el polen, se reduce la fructificación. En muchos cultivares de melocotón, las flores caen antes o alrededor de la división de la cáscara, pero en otros como 'Jersey Queen' y 'Harvester', se forman botones. Los botones son el resultado de flores que aparentemente han cuajado pero que nunca se convierten en frutos de tamaño completo. Los frutos siguen siendo pequeños y deformes a medida que maduran. Si se abren estos frutos, la semilla está muerta. Dado que la formación de botones no es evidente al principio de la temporada, los cultivadores no pueden eliminar los frutos anormales y los botones en desarrollo sirven como fuente de alimento y lugar de hibernación para insectos y enfermedades.
Reducción de la calidad de la fruta:
Los efectos de un frío insuficiente en la calidad de la fruta son probablemente los menos discutidos, pero parecen ser muy comunes, especialmente en el centro y sur de Texas. Los efectos sobre el crecimiento de las hojas y el cuajado de la fruta son dramáticos, pero los efectos de un frío insuficiente sobre la calidad de la fruta son sutiles y pueden aparecer cuando otros síntomas no lo hacen. Un enfriamiento insuficiente hará que muchos cultivares tengan una punta agrandada y una firmeza reducida. Además, la coloración de fondo de la fruta puede ser más verde de lo habitual, posiblemente debido a que la fruta pierde firmeza antes de que el color de fondo pueda cambiar completamente de verde a amarillo. El alcance de estos problemas de calidad depende del cultivar y del grado de frío.
Modelos
Existen varios modelos utilizados para calcular el enfriamiento, cada uno de los cuales define lo que es una unidad de enfriamiento. Los tres modelos más comunes son el modelo del número de horas por debajo de 7 °C, el modelo del número de horas entre 2 y 7 °C y el modelo de Utah. Los dos primeros modelos son sencillos y definen una unidad de refrigeración como una hora por debajo o entre determinadas temperaturas. El método Utah es más complejo porque introduce el concepto de eficacia relativa del enfriamiento y acumulación negativa de frío (o negación del enfriamiento).
En FieldClimate utilizamos el modelo de cálculo de las porciones frías (CP). Las acumulaciones de frío se calculan como proporciones de frío, utilizando un intervalo de temperaturas de 2 a 7°C. Los cálculos de las proporciones de frío terminan después de 96 horas de igual o más entonces >15°C '(se mantiene entre7 y 15°C)
Los cálculos se basan en el trabajo de Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276: 165-174.
Sarna del peral
Fuente: J.W. Travis, J.L. Rytter y K.S. Yoder
Introducción
La sarna del peral es una enfermedad de importancia económica en todo el mundo y puede causar graves pérdidas en cultivares susceptibles. La enfermedad es más problemática en los países europeos que en Norteamérica y es especialmente preocupante en Japón. A veces llamada mancha negra, la sarna del peral se parece a la sarna del manzano (Venturia inaequalis) en casi todos los aspectos, y está causada por un hongo estrechamente relacionado, V. pirina. Los cultivares de pera difieren en susceptibilidad a la sarna; sin embargo, los cultivares resistentes en una región del país pueden no serlo en otra.
Síntomas
Los síntomas de la sarna del peral son muy similares a los de la sarna del manzano. Las lesiones en hojas y pecíolos comienzan como manchas redondas de color marrón que acaban adquiriendo un aspecto aterciopelado. Dentro de estas lesiones se producen conidios. Más adelante en la temporada, pueden observarse pequeñas manchas en la superficie inferior de las hojas. Suelen ser el resultado de infecciones de finales de primavera o principios de verano. La infección de las hojas del peral no es tan común como la sarna del manzano.
Ciclo de la enfermedad
Las lesiones de sarna en la fruta se producen en el extremo del cáliz y, con el tiempo, en los lados de la fruta. A medida que estas lesiones se agrandan, adquieren un color marrón oscuro y forman grandes áreas negras al unirse. Las lesiones en frutos inmaduros son pequeñas manchas circulares aterciopeladas. A medida que la fruta madura aparecen manchas más oscuras y puntiformes. La fruta infectada a menudo adquiere una forma irregular. A diferencia de la sarna del manzano, la sarna del peral suele infectar las ramas. Al principio de la temporada de crecimiento, las lesiones en los brotes jóvenes aparecen como manchas marrones aterciopeladas. Más tarde, estas lesiones se convierten en áreas corchosas, parecidas a cancros. En la primavera siguiente, se desarrollan pústulas dentro de estas lesiones. Estas pústulas producen esporas (conidios) que perpetúan la propagación de la enfermedad. El hongo pasa el invierno en las hojas del suelo y también en forma de micelio en las ramas infectadas. La infección del follaje y los frutos del peral se produce en condiciones similares a las requeridas para la infección del manzano por el hongo de la sarna del manzano. Las ascosporas son la principal fuente de inóculo primario. La infección se produce en primavera, alrededor de la fase de punta verde del desarrollo del botón floral. Las ascosporas de las hojas invernadas se liberan como consecuencia de la lluvia y son transportadas por las corrientes de aire hasta las hojas jóvenes y la fruta. Las ascosporas continúan madurando durante un periodo de seis a ocho semanas. Las conidias son la fuente de inóculo secundario y se producen en las lesiones primarias iniciadas por las ascosporas o en las pústulas de las ramas infectadas. Pueden producirse muchos ciclos secundarios a lo largo de una temporada de crecimiento. La duración del período de mojado y la temperatura necesaria para la infección dependen del número de horas de mojado continuo y de la temperatura durante este período de mojado===. La tabla de Mills para determinar los períodos de infección de la sarna del manzano junto con un registrador de humedad foliar o un higrotermógrafo pueden proporcionar la información para determinar los períodos de infección de la sarna del peral. Las lesiones de sarna pueden desarrollarse en tan sólo ocho días después de la infección en hojas jóvenes y en hasta dos meses en hojas más viejas. Los frutos también son más susceptibles cuando son jóvenes; sin embargo, los frutos maduros pueden infectarse si la duración del periodo de humectación es suficientemente larga.
Supervisión
No es necesario que los cultivadores realicen ningún seguimiento durante el período de latencia. Consulte con el personal regional del Servicio de Extensión Cooperativa para determinar el inicio de la madurez de las ascosporas. El conocimiento de la situación del inóculo de la sarna en huertos adyacentes abandonados o comerciales puede influir en las decisiones de control de la sarna a principios de la temporada. Durante el período previo a la floración y hasta el cuajado de la fruta, tanto fresca como transformada, determine los períodos de infección de la sarna del peral observando la duración de la humedad de las hojas y las temperaturas medias durante el período húmedo.
En Venturia pirina Modelo de infección diseñado por Spotts, R. A., y Cervantes, L. A. 1991
Variables de entrada medioambientales: Temperatura, duración de la humedad.
Descripción del modelo: Spotts y Cervantes presentan datos de un experimento en ambiente controlado con plántulas de peral, así como de experimentos de embolsado de ramas en el campo, sobre los efectos de la temperatura y la duración de la humedad en las infecciones conidiales de plántulas, hojas y frutos de peral. No han evaluado las condiciones de infección por Ascosporas, pero sugieren que deberían ser bastante similares a las condiciones de infección por conidios y, por tanto, su modelo puede utilizarse para predecir la infección primaria por Ascosporas.
Umbral de acción: Los desarrolladores del modelo observaron que la duración mínima de la humedad requerida para la infección del follaje por conidios se situaba entre los valores requeridos para la infección "ligera" y "moderada" de la manzana por V. inequalis. Ascosporas según la tabla de Mills. Por lo tanto, al utilizar la tabla de Mills para la infección por ascosporas o conidios de la sarna del peral, los autores recomiendan el uso de horas de humedad para la infección "leve" a fin de ser más conservadores.
(c) Dr. Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2009
Mancha de la hoja de Fabraea
Mancha foliar, causada por el hongo Fabrea maculatumes una enfermedad muy extendida y destructiva de la punta roja (Photinia fraseri), níspero (Eriobotrya japonica), espino blanco (Rhaphiolepis indica), algunos cultivares de pera (Pyrus sp.) y otros miembros de la familia de las rosáceas. Esta enfermedad es más dañina para las plantas del paisaje y los viveros durante los periodos de tiempo fresco y húmedo y cuando se produce un crecimiento activo.
Síntomas
Los primeros síntomas de la enfermedad son pequeñas manchas circulares de color rojo brillante en el haz y el envés de las hojas jóvenes en expansión. Entomosporium mancha foliar. Numerosas manchas pequeñas pueden unirse en grandes manchas granates en hojas muy enfermas. Las manchas foliares en hojas maduras tienen centros de color marrón ceniza a gris claro con un borde distintivo de color rojo oscuro a granate. En el centro de cada mancha foliar suelen observarse pequeñas motas negras, cuerpos productores de esporas del hongo. Manchas similares a las de las hojas pueden desarrollarse en los pecíolos de las hojas y en los tallos tiernos durante periodos prolongados de tiempo fresco y húmedo.
Los niveles bajos de mancha foliar no suelen causar más que daños estéticos, pero mantienen una fuente de esporas para futuras infecciones. Sin embargo, las infecciones graves suelen provocar la caída temprana y abundante de las hojas. La fuerte caída de las hojas reduce gravemente el valor paisajístico de la punta roja y puede causar la muerte de la planta. Algunos cultivares de espino de la India se ven tan gravemente afectados como el espino rojo.
Ciclo de la enfermedad
Las manchas en las hojas y los brotes jóvenes son importantes para la supervivencia del Entomosporium hongo de la mancha foliar. Las hojas caídas y enfermas son fuentes menos importantes del hongo. Las masas de esporas se liberan durante los periodos de tiempo húmedo desde las estructuras productoras de esporas del hongo situadas en el centro de las manchas desde finales del invierno hasta gran parte del año, excepto durante los periodos calurosos del verano. Estas esporas se propagan al follaje sano mediante una combinación de salpicaduras de agua y viento. Los nuevos síntomas de la mancha foliar aparecen entre 10 y 14 días después de un período de infección húmeda: Las conidias cuatricelulares, con una apariencia distintiva de insecto, se propagan principalmente desde la hojarasca invernante, y algunas desde los cancros de las ramitas, por salpicaduras de agua de lluvia o riego por aspersión. Los periodos de humectación para la infección pueden variar de 8 a 12 horas a temperaturas de 10°C - 25°C. Las lesiones comienzan a aparecer unos 7 días después del inicio del periodo de infección. La enfermedad puede avanzar rápidamente a finales del verano a medida que el viento y la lluvia distribuyen los conidios por todo el árbol. La susceptibilidad de las hojas y los frutos a la infección no disminuye con la madurez. Casi todas las peras de ascendencia europea son susceptibles a esta mancha foliar. Se inicia cuando la humedad de las hojas comienza con la lluvia. Se detiene cuando la humedad de la hoja se interrumpe durante más de 1 hora.
(c) Dr. Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2008
Mancha marrón
Las manchas marrones de la pera están causadas por el hongo patógeno Stemphylium vesicariumque también causa enfermedades en ajos, puerros, cebollas y espárragos. En el peral, el hongo infecta las hojas, los frutos y, en menor medida, las ramas. La necrosis resultante y la podredumbre del fruto se deben a la penetración del hongo en los estomas y lenticelas y a la producción de compuestos químicos que obligan al huésped a matar las células de la zona infectada, lo que provoca las manchas marrones. Las manchas marrones causan graves daños, especialmente en el sur de Europa. Sin embargo, la enfermedad también se ha encontrado en el Adicionalmente, el cultivar de pera predominante en Holanda y Bélgica, es muy susceptible (Montesinos et al., 1995a).
Influencia de la temperatura y la duración de la humedad en la infección conidial por S. vesicarium en pera ha sido estudiada previamente (Montesinos et al., 1995b). Los resultados condujeron al desarrollo de un sistema de predicción de la mancha marrón (Llorente et al., 2000). Las condiciones climáticas en Europa difieren de sur a norte. En el sur de Europa podemos esperar tormentas incluso durante los periodos cálidos, mientras que en el norte pueden producirse periodos de lluvia fría de varias horas incluso a mediados de verano. Es obvio que el sistema de previsión debe revalidarse o incluso adaptarse para su uso en condiciones climáticas diferentes.
Los periodos húmedos que conducen a Stemphylium versicarium infecciones en pera tienen que ser muy largas siguiendo este modelo. El modelo se remonta a los trabajos realizados por Llorente, I., Vilardell, P., Moragrega, C. y Montesinos, E. y la adopción a estaciones meteorológicas electrónicas realizada por A. Boshuizen, P.F. de Jong y B. Heijne de Holanda. Estos periodos húmedos pueden verse interrumpidos. La duración de la interrupción depende de la humedad relativa o del déficit de presión de vapor.
En el cálculo del modelo de FieldClimate, la perturbación puede durar siempre que la humedad relativa sea superior o igual a 75%. Con una humedad relativa entre 65% y 74% la interrupción del periodo húmedo puede durar 12 horas. Si la humedad relativa está entre 55% y 64%, la interrupción puede durar 9 horas. Si la humedad relativa está entre 45% y 54%, la alteración puede durar 8 horas. Si la humedad relativa está entre 35% y 44%, la interrupción puede durar 6 horas. Si la humedad relativa es inferior a 34%, la perturbación puede durar 4 horas.
Las curvas que muestran el progreso de las infecciones leves, moderadas y graves pueden utilizarse como umbrales de actuación para huertos con un historial de la enfermedad diferente. En los huertos con más de 1% de incidencia de la enfermedad en la fruta, el control químico de la enfermedad debe realizarse en todas las infecciones leves. En los huertos con menos de 1% de incidencia de la enfermedad en los frutos, el control químico debe realizarse en todas las infecciones moderadas completadas. Y en los huertos que hasta ahora no tenían incidencia de Stemphylium Deben iniciarse métodos de control si las infecciones graves alcanzarán los 100%.
Condiciones:
Temperatura: 8°C - 38°C
Humedad de la hoja > 0 o humedad relativa >90%
Factor: 600; máx: 60000 (infección 100%).
(c) Dr. Heinrich Denzer, Pessl Instruments GmbH, Weiz, 2009
Equipamiento recomendado
Compruebe qué conjunto de sensores se necesita para vigilar las posibles enfermedades de este cultivo.