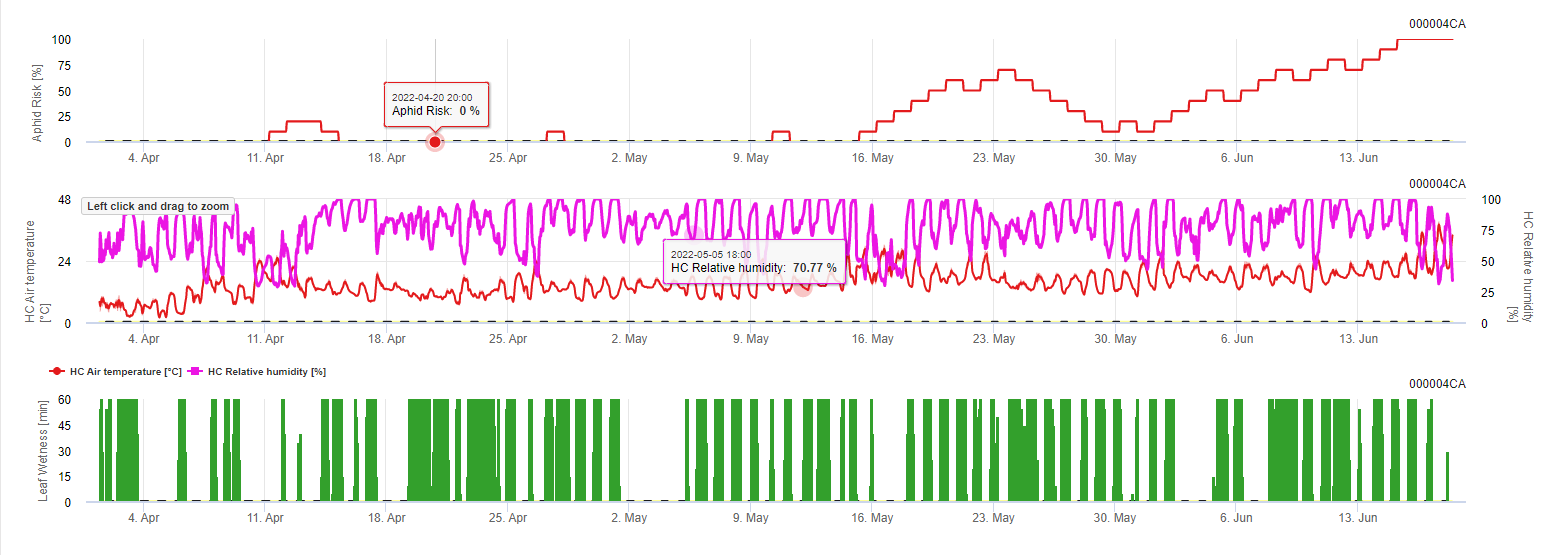
Condiciones:
- por la mañana, cuando sale el sol y disminuye la humedad relativa, temperaturas óptimas entre 20°C y 32°C - buen vuelo indicado.
- Si las temperaturas no están en el rango óptimo (demasiado frío/calor) o es demasiado húmedo (humedad de las hojas) el riesgo disminuye.
- La salida es el riesgo diario.
Así pues, las temperaturas óptimas y las humedades relativas en descenso durante la mañana indican un buen día de vuelo. Cuando hay humedad durante la noche y las temperaturas son demasiado bajas, esto es malo para la propagación. Lo mismo ocurre cuando hace calor y hay humedad durante el día.
El oídio es una enfermedad común en muchos tipos de plantas. Varios hongos de oídio causan enfermedades similares en diferentes plantas (como Podosphaera en manzanas y frutas de hueso; especies de Sphaerotheca en bayas y frutas de hueso; Erysiphe necator en la vid). Los hongos del oídio suelen requerir condiciones húmedas para liberar esporas invernantes y para que esas esporas germinen e infecten el tejido vegetal. Sin embargo, no se necesita humedad para que el hongo se establezca y crezca después de infectar la planta. Los oídio suelen favorecer calienteclimas de tipo mediterráneo.
El oídio puede reconocerse fácilmente en la mayoría de las plantas por el micelio pulverulento de color blanco a gris y crecimiento de esporas que se forma a ambos lados de las hojas, flores, frutos y en los brotes. En los frutos de los árboles aparecerá una mancha áspera y corchosa en la piel donde se ha producido la infección.

Todos los hongos del oídio necesitan tejido vegetal vivo para desarrollarse. En huéspedes perennes de hoja caduca como la vid, la frambuesa y los árboles frutales, el oídio sobrevive de una temporada a otra en yemas infectadas o como cuerpos fructíferos llamados chasmothecia, que residen en la corteza de cordones, ramas y tallos.
La mayoría de los hongos del oídio crecen como una fina capa de micelio en la superficie de la parte de la planta afectada. Las esporas, que son el principal medio de dispersión, constituyen la mayor parte del crecimiento pulverulento y se producen en cadenas que pueden verse con una lupa de mano. En cambio, las esporas del mildiu velloso crecen en tallos ramificados que parecen pequeños árboles. Además, las colonias de mildiú velloso son grises en lugar de blancas y aparecen sobre todo en la superficie inferior de las hojas.
Las esporas del oídio son transportadas por viento a las plantas huésped. Aunque los requisitos de humedad para la germinación varían, muchas especies de oídio pueden germinan e infectan en ausencia de agua. De hecho, el agua sobre la superficie de las plantas mata las esporas de algunos hongos del oídio e inhibe la germinación y el crecimiento del micelio. Temperaturas moderadas y sombra suelen ser las condiciones más favorables para el desarrollo del oídio, ya que las esporas y el micelio son sensibles al calor extremo y a la luz solar directa.
Este hongo pasa el invierno en forma de micelio en el interior de las yemas, la infección primaria se produce cuando las hojas emergen de estas yemas infectadas. Las infecciones secundarias se producen cuando las conidias producidas por las infecciones primarias y secundarias posteriores son sopladas o salpicadas por la lluvia sobre tejidos susceptibles. Los frutos (antes del endurecimiento del hueso) y el crecimiento terminal suculento son susceptibles a la infección.
Las temperaturas medias mínima, óptima y máxima de S. pannosa son de unos 5°, 24° y 24°C. Se forman muchas más conidias en aire seco que en aire húmedo a todas las temperaturas (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli; 1954)
El oídio es común en condiciones de humedad relativa y temperaturas similares a las del oídio del cerezo.
Modelo Fieldclimate: La enfermedad fúngica se modela mediante los factores temperatura y duración de la humedad foliar. Por ejemplo, el 11 de mayo, el periodo de humedad foliar bajo temperaturas moderadas favoreció el desarrollo de la enfermedad y se pudo determinar un riesgo de 100%.
Literatura:
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954): Temperature relations of Powdery Mildews. HILGARDIA. A Journal of Agricultural Science Published by the California Agricultural Experiment Station. Universidad de California. Volumen 22/Número 17.
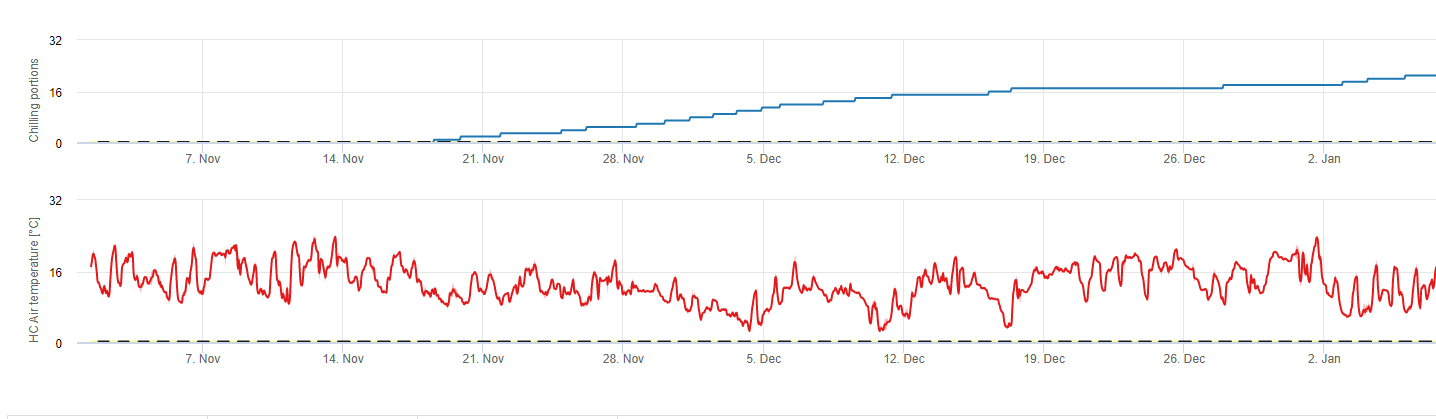
Escalofriante
Los frutales de hueso desarrollan sus yemas vegetativas y fructíferas en verano y, a medida que se acerca el invierno, las yemas ya desarrolladas entran en reposo en respuesta a la menor duración del día y a las temperaturas más bajas. Esta fase de latencia protege a las yemas del frío. Una vez que las yemas han entrado en letargo, toleran temperaturas muy por debajo del punto de congelación y no crecen en respuesta a los periodos cálidos de mediados de invierno. Estas yemas permanecen latentes hasta que han acumulado suficientes unidades de frío (UC). Cuando suficiente frío se acumulan, los brotes están listos para crecer en respuesta a las temperaturas cálidas. Mientras haya suficientes CU, los botones florales y foliares se desarrollan con normalidad. Si los brotes no reciben temperaturas de enfriamiento suficientes durante el invierno para liberar completamente el letargo, los árboles desarrollarán uno o más de los síntomas fisiológicos asociados a un enfriamiento insuficiente: 1) retraso en la foliación, 2) reducción del cuajado y aumento del abotonamiento y, 3) reducción de la calidad del fruto.
Síntomas de enfriamiento insuficientes
Foliación retardada:
Un síntoma clásico de enfriamiento insuficiente es el retraso en la foliación. Un árbol puede tener un pequeño mechón de hojas cerca de las puntas de los tallos y estar desprovisto de hojas de 12 a 20 pulgadas por debajo de las puntas. Los brotes inferiores se romperán con el tiempo, pero la foliación completa se retrasa significativamente, la fructificación se reduce y el árbol se debilita. Además, la fuerte succión de las partes bajas del árbol causa problemas de gestión, y el desarrollo normal de las yemas frutales del año siguiente puede verse afectado.
Fructificación y abotonamiento reducidos:
La floración, en respuesta a un enfriamiento insuficiente, suele seguir el patrón observado en el desarrollo de las hojas. La floración se retrasa, se prolonga y, debido a anomalías en el desarrollo del pistilo y el polen, se reduce la fructificación. En muchos cultivares de melocotón, las flores caen antes o alrededor de la división de la cáscara, pero en otros como 'Jersey Queen' y 'Harvester', se forman botones. Los botones son el resultado de flores que aparentemente han cuajado pero que nunca se convierten en frutos de tamaño completo. Los frutos siguen siendo pequeños y deformes a medida que maduran. Si se abren estos frutos, la semilla está muerta. Dado que la formación de botones no es evidente al principio de la temporada, los cultivadores no pueden eliminar los frutos anormales y los botones en desarrollo sirven como fuente de alimento y lugar de hibernación para insectos y enfermedades.
Reducción de la calidad de la fruta:
Los efectos de un frío insuficiente en la calidad de la fruta son probablemente los menos discutidos, pero parecen ser muy comunes, especialmente en el centro y sur de Texas. Los efectos sobre el crecimiento de las hojas y el cuajado de la fruta son dramáticos, pero los efectos de un frío insuficiente sobre la calidad de la fruta son sutiles y pueden aparecer cuando otros síntomas no lo hacen. Un enfriamiento insuficiente hará que muchos cultivares tengan una punta agrandada y una firmeza reducida. Además, la coloración de fondo de la fruta puede ser más verde de lo habitual, posiblemente debido a que la fruta pierde firmeza antes de que el color de fondo pueda cambiar completamente de verde a amarillo. El alcance de estos problemas de calidad depende del cultivar y del grado de frío.
Modelos
Existen varios modelos utilizados para calcular el enfriamiento, cada uno de los cuales define lo que es una unidad de enfriamiento. Los tres modelos más comunes son el modelo del número de horas por debajo de 7 °C, el modelo del número de horas entre 2 y 7 °C y el modelo de Utah. Los dos primeros modelos son sencillos y definen una unidad de refrigeración como una hora por debajo o entre determinadas temperaturas. El método Utah es más complejo porque introduce el concepto de eficacia relativa del enfriamiento y acumulación negativa de frío (o negación del enfriamiento).
En fieldClimate.com utilizamos el modelo de cálculo de las porciones frías (CP). Las acumulaciones de frío se calculan como proporciones de frío, utilizando un intervalo de temperaturas de 2 a 7°C. Los cálculos de las proporciones de frío terminan después de 96 horas de igual o más entonces >15°C '(se mantiene entre7 y 15°C)
Los cálculos se basan en el trabajo de Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276: 165-174.