La germinación y la penetración de ascosporas de Venturia inaequalis en hojas o frutos dependen de los periodos de humectación de las hojas (hojas húmedas o humedad relativa >80% (mantiene el nivel de infección hasta por debajo de 80% h.r.). y de la temperatura del aire (por encima de 2°C). Las temperaturas más altas conducen, en condiciones húmedas, a una penetración más rápida, mientras que a temperaturas más bajas se necesitan periodos más largos de humectación de las hojas. Esta relación fue publicada por primera vez por MILLS y LAPLANTE (1945). Nuestro cálculo se basa en las publicaciones de SCHWABE (1980). SCHWABE describió la gravedad de las infecciones de sarna en función de la temperatura. Estas observaciones coinciden con las nuestras.
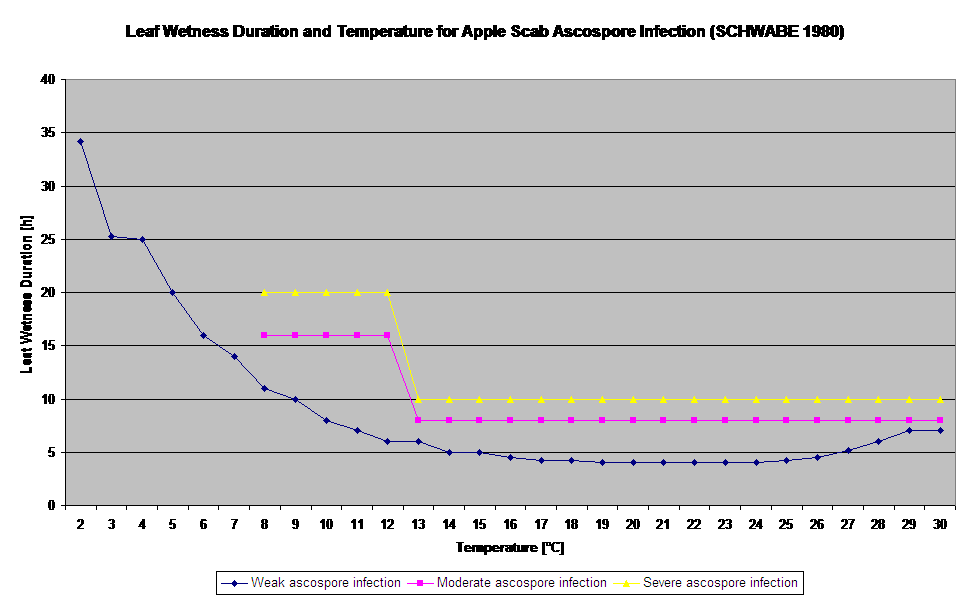
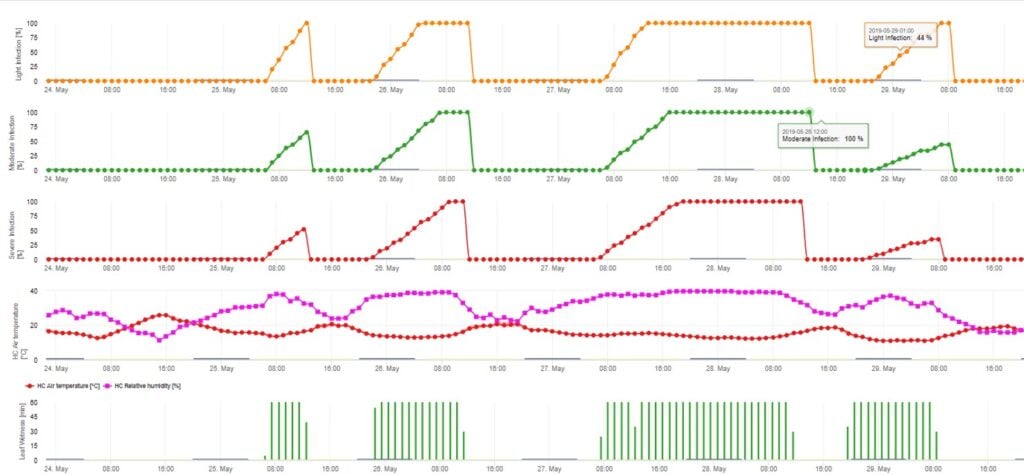
El modelo de infección por ascosporas calcula el progreso de la infección, en función de la humedad y la temperatura de la hoja, para infecciones débiles, moderadas y graves. Dividir la predicción en tres "clases de gravedad" es muy tradicional en los modelos de sarna del manzano. El progreso de la infección se muestra en forma de gráfico para los valores horarios. Una infección ya se ha producido en cuanto la curva de progreso de una infección débil alcanza 100%. Una infección moderada o severa está completa si las curvas de progreso para estas clases de severidad alcanzan (100%). A menudo, en la práctica o en la realidad, una infección alcanza 70% u 80% a primera hora de la mañana y en función del tiempo real podrá saber si esta infección terminará o no. La visualización de los valores máximos diarios de la curva del proceso de infección ofrece una visión general sobre las fechas previstas de infección en la temporada.
Uso principal del modelo
Para el mercado es absolutamente importante tener manzanas libres de enfermedades para conseguir un buen precio. Por lo tanto, los cultivadores tienden a no tener ningún tipo de tolerancia a la sarna en sus huertos. El modelo debe implementarse en un programa de aplicación curativa (aplicación de pulverizaciones que contengan compuestos de ciprodinil o pirimetanil en prefloración) cuando el modelo señale una infección (sin importar si es débil o grave). En el periodo de floración y posfloración temprana, las infecciones graves son más frecuentes y todos los periodos húmedos se han cubierto con pulverizaciones protectoras. Más adelante, los tejidos de la planta crecen muy rápido, por lo que la protección puede que sólo funcione durante 4 días. En este caso, también puede ser necesaria una pulverización curativa tras un periodo de infección incompleto. Pero en este caso, es relevante si ha sido una infección débil o severa y también si había un número bajo o alto de ascosporas listas para la infección.
Literatura
- Schwabe WFS, 1980. Requisitos de humedad y temperatura para la infección de hojas de manzano por Venturia inaequalis en Sudáfrica. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Climatología favorable a la infección de sarna del manzano en Sudáfrica. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management (Biología, epidemiología y gestión de la sarna del manzano). St. Paul, MN: Am. Phytopathol. Soc.: 545.
Infecciones secundarias por conidios
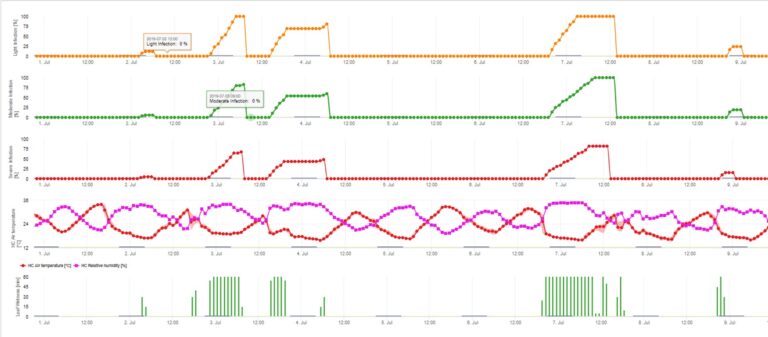
Las infecciones de sarna del manzano por conidios (asexuales) dependen de los mismos factores que las infecciones causadas por ascosporas. La diferencia es que los conidios pueden descargarse en condiciones de luz y oscuridad, y las infecciones por conidios se producen sobre todo en verano.
Dependiendo de los regímenes de temperatura del aire, los conidios de Venturia inaequalis necesitan períodos más o menos largos de humectación de las hojas para germinar y penetrar en las hojas o frutos del manzano. Esta relación fue publicada por primera vez por MILLS y LAPLANTE (1945). Nuestros cálculos se basan en las publicaciones de SCHWABE (1980). Mientras que Mills y LAPLANTE (1945) sólo supusieron que los conidios necesitan un determinado período de tiempo de humedad de la hoja, SCHWABE (1980) también incluyó la importancia de la temperatura en el proceso de infección.
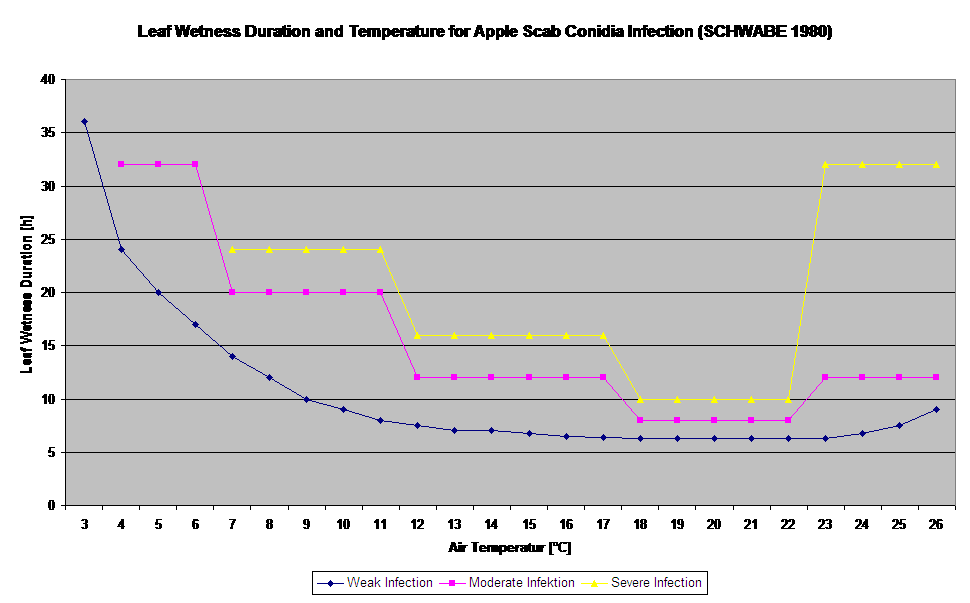
El modelo de infección por conidios calcula el progreso de la infección en función de la humedad y la temperatura de la hoja para infecciones débiles, moderadas y graves. Dividir la predicción en "tres clases de gravedad" es muy tradicional en los modelos de sarna del manzano. El progreso de la infección se muestra en forma de gráfico para los valores horarios. Una infección ya se ha producido en cuanto la curva de progreso de una infección débil alcanza 100%. Una infección moderada o severa está completa si las curvas de progreso para estas clases de severidad alcanzan 100%. A menudo, en la práctica o en la realidad, una infección alcanza 70% u 80% a primera hora de la mañana y, en combinación con el tiempo real, podrá saber si esta infección finalizará o no. La visualización de los valores máximos diarios de la curva de progreso de la infección ofrece una visión general sobre las fechas previstas de infección en la temporada.
Literatura:
- Infección por Venturia inaequalis, señalando las fechas de infección por ascosporas y conidios (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management (Biología, epidemiología y gestión de la sarna del manzano). St. Paul, MN: Am. Phytopathol. Soc.: 545.
Presentación gráfica del modelo
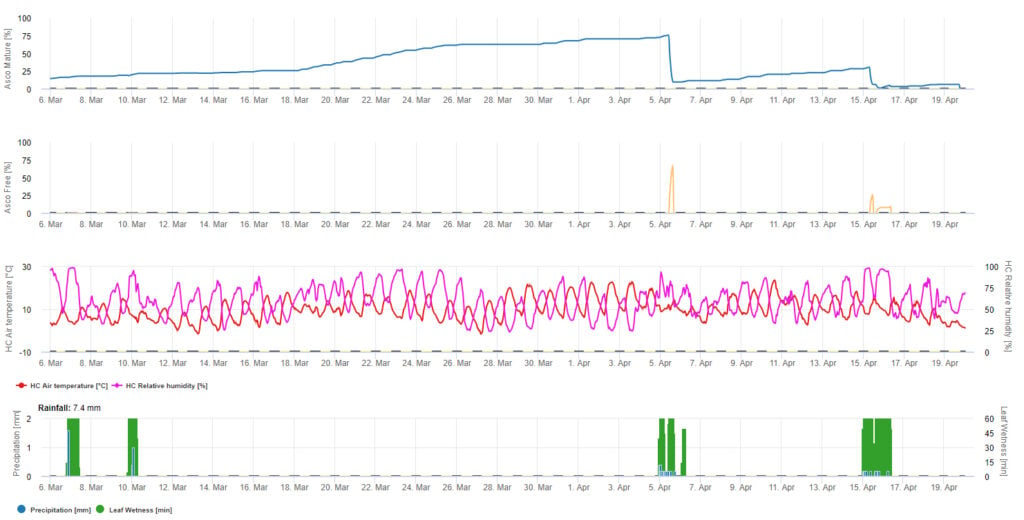
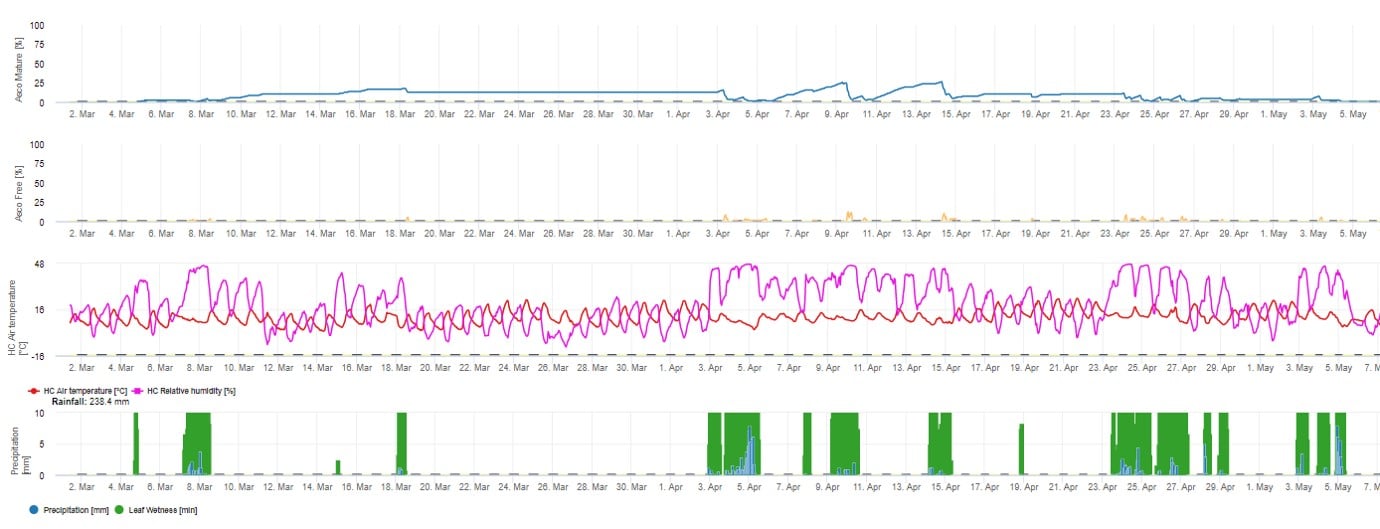
Para la presentación gráfica del modelo de maduración, formación y liberación de ascosporas se utilizan valores diarios. Muestra que los periodos de humedad de las hojas provocan liberaciones de ascosporas y curvas de ascosporas maduras y liberadas. La gravedad de la liberación y el número de ascosporas maduras no pueden evaluarse en valores absolutos. Los valores entre 0 y 100% indican la cantidad relativa de ascosporas maduras y liberadas en función de las condiciones climáticas.
1. Maduración, formación y descarga de ascosporas:
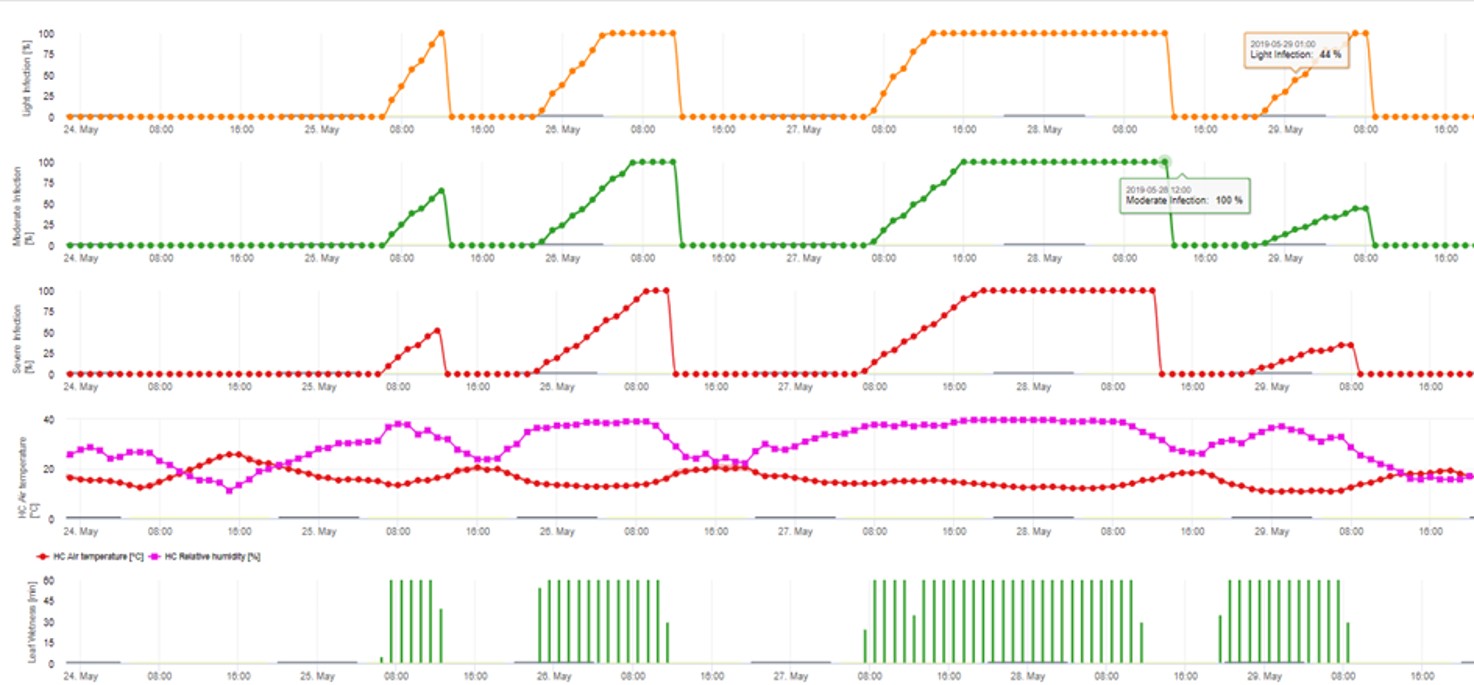
2. Infección por ascosporas mostrada en tres clases de gravedad diferentes (débil: naranja, moderada: verde, grave: rojo). Cuando se alcanzó la infección por 100% se determinaron las condiciones óptimas para que la ascospora infectara el tejido vegetal. Por tanto, deben tenerse en cuenta las medidas de protección de las plantas (profiláctica antes de la infección, curativa después de la infección por 100%):
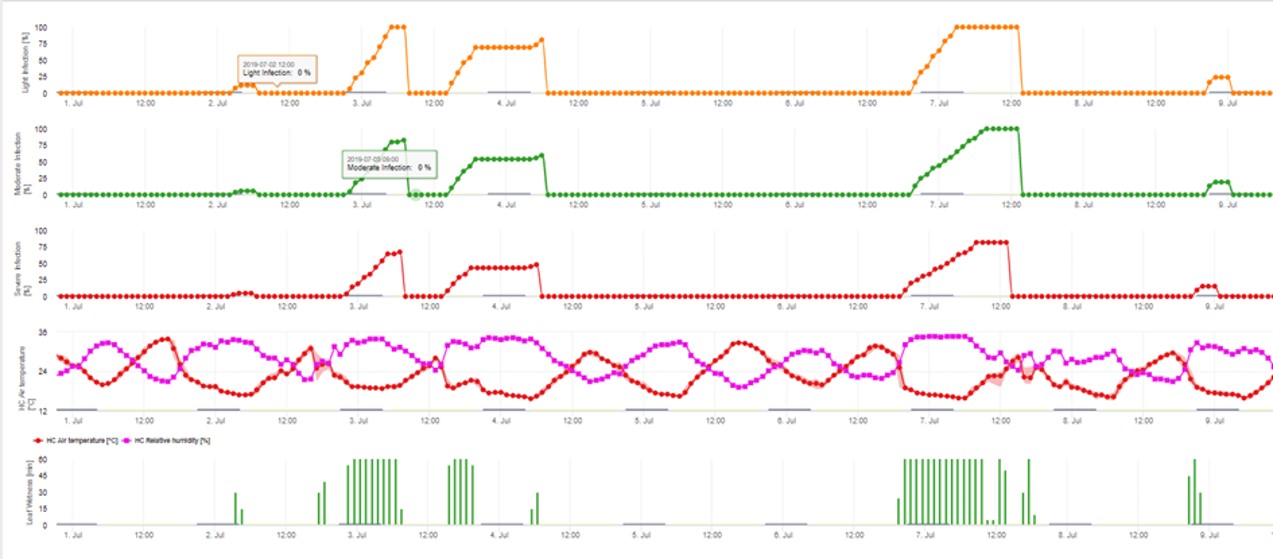
3. Infección por conidios mostrada en tres clases de gravedad diferentes (débil: naranja, moderada: verde, grave: rojo). Cuando se alcanzó la infección por 100% se determinaron las condiciones óptimas para que la ascospora infectara el tejido vegetal. Por tanto, deben tenerse en cuenta las medidas de protección de las plantas (profiláctica antes de la infección, curativa después de la infección por 100%):
Las manzanas cultivadas comercialmente tienen que estar libres de sarna. Las manzanas con costra sólo se venderán para procesado. Por lo tanto, el objetivo de todas las actividades fitosanitarias, tanto en el cultivo convencional como en el ecológico, es tener frutas libres de sarna. Los modelos que muestran la descarga de ascosporas de la sarna del manzano y la infección por ascosporas/conidios son herramientas muy importantes para alcanzar este objetivo.
En los sistemas de cultivo convencionales se utilizan dos tipos básicos de fungicidas contra la sarna del manzano: a) productos preventivos como Captan, Mancozeb, Dithianone y Strobilurins o b) productos curativos como Cyprodinil (Chorus) o Pyrimethanil (Scala) o para su aplicación en los últimos periodos más cálidos de la temporada los fungicidas DMI. En realidad, la mayoría de los productores siguen una estrategia preventiva. Sin embargo, una estrategia preventiva práctica no es capaz de proteger completamente los manzanos porque el manzano crece y desarrolla flores, frutos y hojas. Por lo tanto, la pulverización preventiva sólo protege durante un periodo de 4 a 7 días en función del crecimiento real del árbol. Unos intervalos de pulverización tan estrechos no son manejables, por lo que los cultivadores integrarán su experiencia sobre el clima local, la previsión meteorológica y los modelos de sarna del manzano en su gestión de la pulverización. Programarán las pulverizaciones preventivas basándose en su experiencia y en las previsiones meteorológicas. Los modelos de infección de la sarna del manzano les mostrarán la fecha exacta de la infección (débil, moderada y grave), así como el modelo de descarga de ascosporas/conidios, y con su experiencia podrán estimar la importancia de una infección. Esto da la posibilidad de actuar con un producto curativo si una infección de sarna del manzano se produjo demasiado tiempo después de la última pulverización preventiva.
En la producción ecológica de manzanas, el azufre calcáreo ha resultado ser el agente de control más eficaz contra la sarna. El control óptimo puede lograrse si se pulveriza poco antes de la infección o al principio de la misma. Esto debe planificarse en función de la previsión meteorológica. A veces no se alcanza este periodo óptimo y hay que pulverizar sobre las hojas húmedas de una infección de sarna casi completa. Aún así, la eficacia será buena. Los modelos de sarna del manzano ayudan a decidir si es necesaria una pulverización de emergencia en una infección casi completa.
Síntomas del fuego bacteriano
Los chancros invernales, que albergan el patógeno del fuego bacteriano, son a menudo claramente visibles en troncos y ramas grandes como zonas de corteza descolorida de ligera a profundamente deprimidas, que a veces están agrietadas en los márgenes. El mayor número de cancros, sin embargo, son mucho más pequeños y no se distinguen tan fácilmente. Aparecen en ramas pequeñas donde se produjeron infecciones de flores o brotes el año anterior y, a menudo, alrededor de los cortes realizados para eliminar las ramas afectadas. Dado que muchos de estos cancros se establecen más tarde en la temporada, no suelen estar muy deprimidos y rara vez muestran grietas en la corteza en sus márgenes. Además, suelen ser bastante pequeños, de menos de 2,5 cm, con una corteza entre rojiza y púrpura que puede estar cubierta de minúsculos cuerpos fructíferos de hongos negros (sobre todo Botryosphaeria obtusael patógeno de la podredumbre negra de la manzana).
Los síntomas del tizón de la flor aparecen con mayor frecuencia una o dos semanas después de la floración y suelen afectar a todo el racimo de flores, que se marchitan y mueren, volviéndose marrones en el manzano y bastante negras en el peral. Cuando el clima es favorable para el desarrollo del patógeno, pueden verse glóbulos de exudado bacteriano en las flores. El espolón que soporta el racimo de flores también muere y la infección puede extenderse y matar partes de la rama de soporte. Las puntas de los brotes jóvenes infectados se marchitan, formando un síntoma muy típico de "cayado de pastor". Los brotes más viejos que se infectan después de haber desarrollado unas 20 hojas pueden no mostrar este síntoma de rizado en la punta. A medida que la infección se propaga por el eje del brote, las hojas muestran primero vetas oscuras en las venas medias, luego se marchitan y se vuelven marrones, permaneciendo fuertemente unidas al brote durante toda la temporada. Al igual que con las infecciones de flores, el patógeno a menudo invade y mata una parte de la rama que soporta el brote infectado. El primer síntoma en brotes de agua y brotes que son invadidos sistémicamente desde cancros activos cercanos es el desarrollo de una decoloración amarilla a naranja de la punta del brote antes de que se produzca el marchitamiento. Además, los pecíolos y las venas medias de las hojas basales de estos brotes suelen necrosarse antes que las de la punta del brote.
Dependiendo del cultivar y de su estado de desarrollo en el momento en que se produce la infección, la infección de una sola flor o brote puede provocar la muerte de una rama entera, y cuando se invade la rama central o el tronco del árbol, puede morir una parte importante del árbol en una sola temporada. En general, las infecciones de cualquier tipo que se producen entre la caída de los pétalos y la brotación terminal suelen provocar la mayor pérdida de ramas y árboles. Además, los árboles fuertemente estructurados tienden a sufrir una pérdida de ramas menos severa que aquellos formados con sistemas más débiles para una alta productividad. Cuando se infectan portainjertos de manzano muy susceptibles (M.26, M.9), gran parte del tronco de la púa y las ramas principales por encima de la unión del injerto no suelen presentar síntomas, mientras que se desarrolla un chancro marrón oscuro alrededor del portainjerto. A medida que este cancro rodea el árbol, la parte superior muestra síntomas de decaimiento general (mal color del follaje, crecimiento débil) a mediados o finales de la temporada. En algunos casos, el follaje de los árboles afectados por el chancro del portainjerto desarrolla un color rojo otoñal a finales de agosto o principios de septiembre, parecido al que se asocia a menudo con la enfermedad de la podredumbre del cuello causada por un hongo del suelo. Algunos árboles con infecciones de portainjertos pueden no mostrar síntomas de decaimiento hasta la primavera siguiente, momento en el que pueden observarse cancros que se extienden hacia arriba en la parte inferior del tronco.
Ciclo de la enfermedad
El patógeno bacteriano causante del fuego bacteriano inverna casi exclusivamente en los chancros de las ramas infectadas la temporada anterior. El mayor número de chancros y, por lo tanto, los más importantes en la aportación de inóculo, se producen en ramas de menos de 38 mm de diámetro, especialmente alrededor de los cortes realizados el año anterior para eliminar las ramas afectadas. Durante el comienzo de la primavera, en respuesta a las temperaturas más cálidas y al rápido desarrollo de los brotes, las bacterias de los márgenes de los cancros comienzan a multiplicarse rápidamente y producen un exudado espeso de color amarillento a blanco que se elabora sobre la superficie de la corteza hasta varias semanas antes del periodo de floración. Muchas especies de insectos (sobre todo moscas) se sienten atraídas por el exudado, y posteriormente dispersan las bacterias por todo el huerto. Una vez que las primeras flores abiertas están colonizadas por la bacteria, los insectos polinizadores trasladan rápidamente el patógeno a otras flores, iniciando más tizón de la flor. Estas flores colonizadas están sujetas a infección en cuestión de minutos después de cualquier evento de humectación causado por lluvia o rocío pesado cuando las temperaturas medias diarias son iguales o superiores a 16 °C mientras los pétalos de las flores están intactos (los receptáculos de las flores y los frutos jóvenes son resistentes después de la caída de los pétalos). Una vez que se producen las infecciones de la flor, se pueden esperar síntomas tempranos con la acumulación de al menos 57 días-grado (DD) superiores a 13 °C que, dependiendo de las temperaturas diarias, pueden requerir de 5 a 30 días naturales.
Con la aparición de los síntomas del tizón de la flor, el número y la distribución de las fuentes de inóculo en el huerto aumentan considerablemente. El viento, la lluvia y muchos insectos casuales que visitan las puntas de los brotes jóvenes propagan el inóculo de estas fuentes, lo que aumenta la probabilidad de que se produzca un brote de tizón de los brotes. Investigaciones recientes llevadas a cabo en Pensilvania indican que la alimentación de los áfidos no contribuye al tizón de los brotes. Se necesita más investigación para determinar si las chicharritas desempeñan o no un papel en la incidencia del tizón de los brotes. La mayoría de las infecciones de las puntas de los brotes se producen entre el momento en que los brotes tienen entre nueve y diez hojas y la brotación terminal, cuando se dispone de fuentes de inóculo e insectos vectores y las temperaturas diarias alcanzan una media de 16 °C o más.
En los años en los que no se producen infecciones en flor, las principales fuentes de inóculo para la fase de tizón de los brotes son los cancros invernantes y, en particular, los brotes de agua jóvenes cercanos a estos cancros, que se infectan a medida que las bacterias se desplazan hacia ellos sistémicamente desde los márgenes de los cancros. Tales infecciones sistémicas de los brotes, denominadas tizón del cancro, se inician aparentemente alrededor de 111 DD mayores de 13 °C después de la punta verde, aunque los síntomas visibles pueden no ser aparentes hasta la acumulación de al menos 157 DD mayores de 13 °C después de la punta verde. En ausencia de infecciones de la flor, el desarrollo de las infecciones del tizón de los brotes se localiza a menudo alrededor de las zonas con cancros invernantes.
Tizón de la flor modelo
El modelo requiere que el usuario reconozca los acontecimientos locales específicos y siempre cambiantes y los aspectos de su huerto que pueden aumentar o disminuir el riesgo de niebla del peral y del manzano en relación con otros huertos de la región. El modelo requiere que el usuario suponga que existe riesgo de infección por niebla del peral y del manzano siempre que haya flores en los árboles, especialmente durante el período de caída de pétalos y "postfloración", cuando pueden quedar flores dispersas en muchas variedades de manzanos y perales. Se pide al usuario del modelo que evalúe cuidadosamente la situación en su emplazamiento específico y que inicie medidas de control si hay flores, los niveles de riesgo son "Alto" o "Extremo", y es probable que se produzca la mojadura de las flores en algún momento durante las próximas 24 horas.
Estructura del modelo: Temperaturas y humedad: El proceso clave del fuego bacteriano que debe modelarse es el potencial de crecimiento bacteriano en los estigmas de las flores del manzano y el peral. Este crecimiento depende de la temperatura, por lo que una predicción fiable del riesgo de infección requiere el uso de un método de medición que refleje con la mayor exactitud el crecimiento de la bacteria en los estigmas. Erwinia amylovora colonias. El principal desacuerdo entre los modelistas es cómo debe hacerse.
El modelo Cougar blight estima la tasa de crecimiento bacteriano con las horas-grado basándose en una curva específica de tasa de crecimiento. Esta curva de crecimiento se basa en la tasa de crecimiento de E. amylovora bacterias en pruebas de laboratorio. Los valores de grados hora se acumulan cada hora del día en que las temperaturas superan los 15 °C. Los valores horarios aumentan a medida que las temperaturas suben de 15 °C a 29 °C, disminuyen a temperaturas más altas y llegan a cero en cualquier hora con temperaturas superiores a 40 °C.
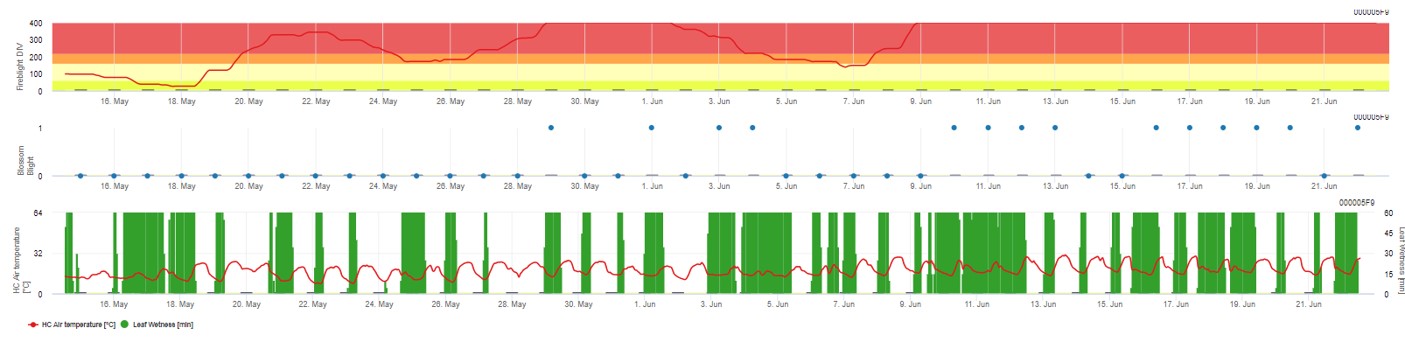
En FieldClimate.com ambos modelos aparecen en la misma figura. El modelo Fireblight DIV se basa en Courgarblight y el modelo Blossomblight en Maryblight.
Modelo cougar blight
La incidencia del tizón de la flor es uno de los aspectos más esporádicos de las epidemias de fuego bacteriano. El modelo se construyó partiendo del supuesto de que existe una abundancia de inóculo y de que, para que se produzca un evento de infección de la flor, deben cumplirse en secuencia cuatro condiciones estrictas. Estas condiciones son:
1) la flor debe estar abierta con estigmas y pétalos intactos, los estigmas tienen que estar expuestos para la colonización, las flores en caída de pétalos son resistentes;
2) la acumulación de al menos 110 °C horas > 18,3°C en los últimos 44°C días > 4,4°C define el potencial de infección epifítica de la flor abierta más antigua y, por tanto, más colonizada del huerto
3) un acontecimiento húmedo en forma de rocío o de 0,2 mm de lluvia o de 2,5 mm de lluvia el día anterior permite el desplazamiento de las bacterias de los estigmas colonizados a los nectártodos
4) una temperatura media diaria de >= 15,6°C: Esto puede influir en la velocidad a la que las bacterias migran a los nectártodos, así como en la multiplicación de bacterias necesaria para establecer infecciones.
Presentación gráfica de los modelos
Ambos modelos se muestran en la misma página. La presentación gráfica de los modelos de niebla del peral y del manzano es mejor utilizarla con valores agregados sobre la base diaria. El gráfico muestra los valores climáticos relacionados con esta enfermedad. El resultado del Tizón del puma denominado Tizón del fuego DIV y los resultados del Tizón de las flores. Para interpretar los resultados del tizón del puma, el gráfico se subraya con 5 colores diferentes. La distribución de estos colores se realiza en función de los ajustes del primer historial de tizón del huerto (véase el botón de la parte superior derecha). Los 5 colores indican la clase de riesgo de los valores DIV.
Interpretación de los valores (en función de la incidencia del fuego bacteriano en los últimos años, configuración básica):
Presencia de cancros activos en las proximidades
Configure los ajustes del historial de fuego bacteriano con el botón azul "Ajustes" situado en la parte superior derecha:
Visualización de los resultados del modelo Fire Blight mediante 1) Modell Cougar Blight (llamado Fireblight DIV en función de la historia y los colores de fondo) y 2) Modell Blossom Blight
| Presencia potencial de patógenos | Bajo | Moderado | Alta | Extremo |
| Presencia potencial de patógenos | Bajo | Moderado | Alta | Extremo |
| No ha habido fuego bacteriano en los dos últimos años | 0-350 | 350-500 | 500-800 | 800+ |
| Dos temporadas de fuego bacteriano en la zona | 0-300 | 300-500 | 500-750 | 750+ |
| Dos temporadas de fuego bacteriano en la zona | 0-250 | 250-450 | 450-700 | 700+ |
| Fuego bacteriano en un huerto el año pasado | 0-200 | 200-350 | 350-500 | 500+ |
Umbral de acción:
Se aconseja el control si se detecta un periodo de infección de riesgo alto o extremo. Los factores que determinan los daños son: número de flores, edad del árbol, vigor y variedad,... también.
La oportunidad de una infección de Tizón de la Flor se muestra mediante una barra de 0- 1 (por lo que todas las condiciones se cumplen o no).
El objetivo de los modelos de niebla del peral y del manzano es evaluar la probabilidad de una infección por Erwinia amyloflora en el huerto.
El modelo de Mary Blight que se está evaluando para el tizón de la flor indica muy bien situaciones de infección de alto impacto económico. Por esta razón, se utiliza con bastante frecuencia para indicar el uso de antibióticos contra este patógeno. El tizón del puma proporciona información sobre el riesgo de infecciones de fuego bacteriano debido a las posibilidades de propagación del patógeno. Su ponderación realizada por el historial de un huerto es muy útil para indicarnos con qué cuidado tenemos que comprobar los huertos en busca de síntomas de fuego bacteriano, incluso en situaciones en las que el tizón de Mary no indicará una infección.
Biología de la polilla del bacalao
En C. pomonella pasa el invierno como una larva madura hilada en un capullo que se encuentra en zonas protegidas del árbol (grietas o hendiduras de la corteza), que es el lugar preferido, o en la base del árbol en la hojarasca del suelo. En los manzanos modernos de alta densidad hay muy pocos lugares de capullo en los árboles debido a la corteza lisa. Las pupas se forman en el capullo hilado por la larva madura.
El estado adulto emerge de la caja pupal y, tras un breve periodo para endurecer el exoesqueleto, vuela hacia el interior del árbol. La mayor parte de la bibliografía indica que la actividad de la polilla se produce en el tercio superior de la copa de los árboles. Los adultos están activos al anochecer durante un par de horas con un umbral de actividad medio de entre 13°C y 15°C. El apareamiento suele tener lugar en el hospedador y la puesta de huevos se produce inmediatamente después. En las noches cálidas, es posible que una hembra emerja, se aparee y ponga huevos en un periodo de dos días. A C. pomonella La polilla puede vivir más de 20 días en condiciones ideales, pero en el campo es probable que sobreviva mucho menos tiempo, sobre todo en el caluroso verano. En condiciones de laboratorio, una polilla hembra pone hasta 100 huevos, pero la fecundidad real en el campo es probablemente menor y la mayoría son puestos en los primeros días tras la emergencia y el apareamiento.
Los huevos se ponen individualmente en la superficie superior de la hoja, en ramitas o en frutos. En la primera generación, los huevos se depositan únicamente en las hojas hasta unas dos semanas después del cuajado de los frutos, momento en que desaparece la borrosidad de éstos. En la segunda generación, alrededor del 65% de los huevos se ponen en hojas, pero éstas suelen estar muy cerca de un fruto. Los huevos son de color blanco cremoso cuando se ponen por primera vez, tienen un anillo rojo a mitad de su desarrollo y cuando están a punto de eclosionar se puede ver la cápsula negra de la cabeza.
Una larva recién eclosionada deambula hasta que localiza una fruta. Mastica la piel y se alimenta de la pulpa de la manzana bajo la piel durante unos días. En el proceso de masticar la piel y alimentarse por debajo, la larva expulsa los excrementos por la zona de entrada. A menudo aparece un anillo rojo alrededor de la entrada, especialmente en la fruta joven. Cuando la larva muda a la siguiente fase, perfora hacia el núcleo, donde se alimentará de semillas hasta que madure. Cuando está a punto de madurar, la larva hace un túnel desde el corazón hasta el exterior de la manzana y tapa el agujero con excrementos y seda. Cuando está lista para hilar un capullo, la larva abandona la fruta, paseándose por el árbol o cayendo al suelo sobre un hilo de seda, y busca un lugar protegido.
Modelo para la polilla del bacalao
Nuestro modelo se basa en la acumulación de las temperaturas efectivas entre >= 10°C y <=32°C. En el hemisferio norte, esta acumulación comienza el 1 de enero. En el hemisferio sur, comienza el 1 de julio. Acumulamos cada medición en su equivalente para el día (véase el gráfico siguiente). Se utilizan los siguientes umbrales para el desarrollo de los insectos:
*Generación Completa= 650
| *De la puesta a la eclosión del huevo | = 88 |
| *Primer estadio larvario | = 60 |
| *Segundo estadio larvario | = 65 |
| *Tercer estadio larvario | = 55 |
| *Etapa de larva avanzada | = 45 |
| *Quinto estadio larvario más pupas | = 335 |
El gráfico elaborado por el modelo muestra la primera aparición y la duración de las larvas de los estadios 1 a 5 y la presencia de adultos en 5 niveles.
| *Nivel 1: | Actividad de vuelo, no puesta de huevos |
| *Nivel 2: | Actividad de vuelo con posible puesta de huevos. |
| *Nivel 3: | Actividad de vuelo con algo de puesta de huevos. |
| *Nivel 4: | Actividad de vuelo con puesta de huevos de razonable a fuerte. |
| *Nivel 5: | Actividad de vuelo con fuerte puesta de huevos. |
Presentación de los estadios adultos de la polilla del bacalao en 5 niveles diferentes (primer gráfico), así como los grados-día acumulados (línea roja, segundo gráfico) para el desarrollo de los adultos y los estadios larvarios mostrados en azul (tercer gráfico).
En los manzanos hay varias especies de pulgones. El pulgón verde del manzano, el pulgón de la avena o pulgón de la manzana y el pulgón rosado del manzano. En los jardines privados, sobre todo, el pulgón rosado del manzano tiene cierta importancia. Este pulgón provoca fuertes cambios hormonales en las puntas de los brotes con infestaciones. Las hojas se rizan y el grito se acorta. Más tarde, las hojas amarillean y mueren. Este pulgón afecta sobre todo a los brotes jóvenes. El pulgón de la manzana rosada puede producir daños graves ya en pequeña abundancia. Mientras que el pulgón verde de la manzana y el pulgón de la hierba del manzano pueden aparecer en mayor abundancia sin provocar daños económicos. El pulgón de la manzana debe considerarse de importancia no económica.
Pulgón rosado del manzano
Se trata de una antigua especie europea introducida en Estados Unidos hacia 1870. Se convirtió en una plaga importante de las manzanas a finales del siglo XIX. Para prosperar, esta especie debe tener abundancia de su planta huésped de verano, el plátano de hoja estrecha. Existe un notable paralelismo entre la introducción y propagación del pulgón rosado del manzano y el llantén de hoja estrecha. Este pulgón está presente en todas las zonas frutícolas del mundo.
Etapas de la vida
Huevo: El huevo es ovalado y ligeramente aplanado en el lado próximo a la corteza. Su longitud varía entre 0,49 y 0,56 mm. Cuando se pone por primera vez, es de color amarillo brillante y está cubierto de una sustancia glutinosa que se endurece con la edad. El color cambia gradualmente a amarillo verdoso y finalmente a un negro azabache brillante. El tiempo necesario para este cambio de color varía, en condiciones normales al aire libre, de unos nueve días a más de dos semanas.
Ninfa: Los individuos que salen de los huevos son hembras vivíparas sin alas. Hay cinco instares. El último es la madre madura, que poco después del cuarto estadio comienza a producir crías vivas partenogenéticamente. Produce una media de cinco a seis crías al día. Las ninfas de la segunda generación, todas hembras, alcanzan la madurez en dos o tres semanas. La gran mayoría de las ninfas comienzan a reproducirse en la manzana, aunque unas pocas pueden desarrollar alas y migrar al plátano.
La tercera generación se produce en junio y principios de julio. La mayoría de esta generación desarrolla alas y migra al plátano de hoja estrecha. En algunas temporadas, las hembras sin alas de la tercera generación producen una cuarta generación en el manzano. En los últimos años, se ha observado en algunas zonas que las poblaciones dañinas de pulgón rosado del manzano han persistido en los huertos hasta mediados del verano o más tarde. No se sabe si este cambio se debe al proceso de selección, a cambios de hábitos o a la falta de necesidad de un hospedador alternativo.
Adulto: El adulto varía considerablemente en las marcas de color. El color general es marrón rosado, con un tinte rosáceo debido a una cubierta pulverulenta. Algunos de los adultos más viejos son morados, mientras que los más jóvenes son decididamente rosa rojizo.
Alcance del host
El manzano es el huésped preferido, pero el pulgón también se alimenta de peral y espino. Cortland, Golden Delicious, Rhode Island Greening e Ida Red son variedades de manzana especialmente sensibles. El pulgón se encuentra en todas las zonas frutícolas de Estados Unidos y Canadá.
Lesiones o daños: La alimentación del pulgón rosado del manzano a menudo provoca que las hojas se enrosquen, empezando por la caída de los pétalos. Posteriormente, estas hojas pueden volverse de color rojo brillante. La alimentación de las hojas alrededor de los racimos de frutos suele provocar el amontonamiento, el retraso del crecimiento y la malformación de los frutos en desarrollo. Estas anomalías se agravan a medida que la fruta se desarrolla y pueden llegar a hacerla invendible.
Las grandes poblaciones de pulgones pueden producir grandes cantidades de melaza como residuo de la savia de la que se alimentan. La melaza excretada sobre la fruta servirá de caldo de cultivo para el hongo moho de hollín, que afectará al acabado de la manzana. Las toxinas presentes en la saliva de los pulgones también sirven como "gota de parada", impidiendo la abscisión de los frutos (separación natural del árbol) en la cosecha normal.
Historia vital: El pulgón pasa el invierno en fase de huevo. La eclosión se produce a principios de la primavera, entre una semana y 10 días más tarde que la del pulgón de la manzana y más o menos al mismo tiempo que la del pulgón de la manzana. Los huevos eclosionan cuando los brotes empiezan a abrirse en primavera, durante un periodo de dos semanas. En cuanto eclosionan, las crías buscan los brotes de apertura de la manzana; parecen preferir los brotes frutales. Se alimentan de la parte exterior de las yemas de las hojas y de los racimos de yemas frutales hasta que las hojas empiezan a desplegarse. Entonces descienden al interior de los racimos y empiezan a chupar la savia de los tallos y los frutos recién formados.
Su alimentación hace que las hojas se enrosquen, protegiendo a los pulgones de las pulverizaciones y de algunos enemigos. El severo enrollamiento del follaje provocado por esta especie es probablemente su rasgo más característico. Una sola madre de tallo situada en el envés de una hoja, cerca del nervio central, hará que la hoja se doble con fuerza. Bastan unas pocas madres de tallo para provocar un rizado severo de todas las hojas que rodean un capullo floral en apertura, proporcionando una protección ideal a los pulgones que se desarrollan rápidamente. Las madres del tallo alcanzan la madurez cuando los manzanos están floreciendo.
Las madres maduras son muy inactivas. Se instalan, se alimentan y producen crías a gran velocidad. Cuando se distribuyen, retiran rápidamente el pico de los tejidos vegetales y buscan otro lugar en el que continuar. Las madres de tallo maduran unas dos semanas después de la eclosión. La duración depende en gran medida de las condiciones meteorológicas. La producción de crías suele comenzar dos o tres días después de la última muda y continúa sin interrupción durante más de un mes.
La producción total de una sola hembra es de 185, por término medio. Normalmente, el periodo de reproducción se extiende desde principios de mayo hasta junio. Por lo general, el período máximo de actividad reproductiva se sitúa en torno a la última semana de mayo y la primera de junio, cuando los frutos jóvenes empiezan a cuajar e inician el crecimiento activo. El pulgón rosado del manzano rara vez ataca a los brotes jóvenes y de crecimiento rápido. Se limita al follaje, los tallos florales y los frutos jóvenes.
Uno de los rasgos característicos de esta especie es la congregación de las crías alrededor de la madre. Cada madre o grupo de madres tendrá cientos de crías. Las hojas infestadas pronto pueden estar cubiertas, en algunos casos, por más de una capa de pulgones. Este hábito de congregarse pronto mata las hojas infestadas y provoca la migración forzada de los pulgones. Los jóvenes se mueven activa y apresuradamente, aparentemente ansiosos por encontrar un lugar adecuado para alimentarse. Es frecuente encontrarlos durante este periodo congregados en los frutos en formación o atacando el nuevo y suculento follaje que se despliega.
La segunda generación necesita de cuatro a 40 días para alcanzar la madurez y producir crías. La mayor parte de la segunda generación son hembras sin alas. La producción total media de cada individuo de la segunda generación es de unos 119. Los hábitos y actividades de la tercera generación no difieren de los de la segunda. Los áfidos se congregan en grandes cantidades en el envés del follaje, causando graves rizos. También atacan el cuajado y los frutos en desarrollo, produciendo lesiones características.
La mayor parte de esta tercera generación adquiere alas y migra al plátano de hoja estrecha, la planta huésped de verano. Puede producirse una cuarta generación. Todos ellos producen alas y migran al plátano de hoja estrecha. Tras la última muda, los adultos alados son muy tiernos e inactivos. Permanecen retenidos en las hojas rizadas durante dos o tres días antes de emprender su vuelo migratorio. Justo antes de volar, se vuelven muy activos y nerviosos, correteando o moviendo las alas arriba y abajo en previsión de su vuelo.
Pulgón verde de la manzana
El pulgón verde del manzano, Aphis pomi (DeGeer), está ampliamente distribuido en todas las zonas productoras de manzanas. En el hemisferio norte aparece por primera vez en los manzanos a finales de mayo o principios de junio. Este insecto succiona la savia de las hojas de los brotes de agua y del crecimiento terminal suculento. El pulgón verde de la manzana suele encontrarse cerca de las venas principales, en el envés de la hoja. El pulgón verde de la manzana también ataca al peral, el espino blanco, el membrillo, el manzano cangrejo y la espirea.
Descripción del pulgón verde del manzano
Los huevos del pulgón verde de la manzana son ovalados y de color negro brillante. Las ninfas y los adultos son de color verde oscuro o claro. El pulgón verde de la manzana tiene la cabeza verde, el abdomen uniformemente verde o verde amarillento, las patas negras y largos cornículos negros, los "tubos de la cola" (en el extremo posterior del insecto). Las hembras adultas son de color verde oscuro a negro y tienen alas. Durante el verano, estas hembras, o alados, dan a luz a crías vivas de forma partenogénica.
Biología del pulgón verde de la manzana
El pulgón verde de la manzana pasa el invierno en forma de huevo en los chupones, en la base de las yemas y en las cicatrices de las hojas de los brotes terminales. Los huevos suelen eclosionar cuando estallan las yemas y se despliegan las primeras hojas. Las ninfas comienzan a alimentarse inmediatamente de las hojas en desarrollo. Inicialmente están presentes en el brote terminal y más tarde se desplazan a las hojas más viejas del racimo. Las hembras adultas de pulgón producen hembras sin alas en dos semanas, y cada hembra es capaz de producir entre 50 y 100 crías vivas. Los pulgones jóvenes se desarrollan en siete a diez días. Las poblaciones de pulgón verde de la manzana suelen acumularse en la manzana lentamente a principios de primavera (floración, pétalo, otoño), y más rápidamente a medida que aumentan las temperaturas medias diarias. Durante el verano se producen pulgones con alas, o alados, y pulgones sin alas. Los pulgones verdes de la manzana son más numerosos durante julio y principios de agosto. Hay más ninfas que adultos alados en las fases iniciales de una infestación. El porcentaje de alados aumenta a medida que crece la población. Las colonias de pulgón verde del manzano pueden duplicarse en una semana si las condiciones son favorables. Si los pulgones no tienen un nuevo crecimiento suculento y tienen que alimentarse de hojas más viejas, el número de crías producidas puede descender hasta 50%. Si las temperaturas diarias son de 30-32ºC y superiores, las hembras no se reproducen bien. Cuando las temperaturas son altas durante varios días, los pulgones mueren. Durante el verano se desarrollan varias generaciones del pulgón verde de la manzana, y las formas aladas se dispersan por el huerto. A veces, el pulgón verde del manzano y el pulgón rosado del manzano, Dysaphis plantaginea Pars. tienen colonias en la misma hoja. El pulgón rosado del manzano es de color rosa o púrpura. En otoño, machos y hembras sin alas se desarrollan y se aparean, y las hembras ponen huevos para el invierno. El mayor número de huevos se encuentra a 15-20 cm de las puntas de las ramitas. Los huevos rara vez se encuentran en las grandes ramas de los andamios o en los troncos de los manzanos.
Daños
El pulgón verde de la manzana succiona la savia de la hoja. Las infestaciones intensas reducen el vigor y el crecimiento de los brotes. Esto es especialmente preocupante en viveros y huertos jóvenes no productivos. El insecto puede reducir el tamaño de los brotes y la longitud de los entrenudos, y provocar el enrollamiento de las hojas. La alimentación del pulgón también puede estimular el crecimiento de ramas laterales que pueden afectar a la forma del árbol. El enrollamiento de las hojas y los terminales debilitados son susceptibles de sufrir daños en invierno. La melaza producida por los pulgones puede gotear sobre la fruta, lo que permite el crecimiento de hongos de hollín. El hongo mancha la fruta y reduce su valor de mercado. Cuando las infestaciones son abundantes, el pulgón verde de la manzana puede alimentarse de manzanas inmaduras y provocar russeting.
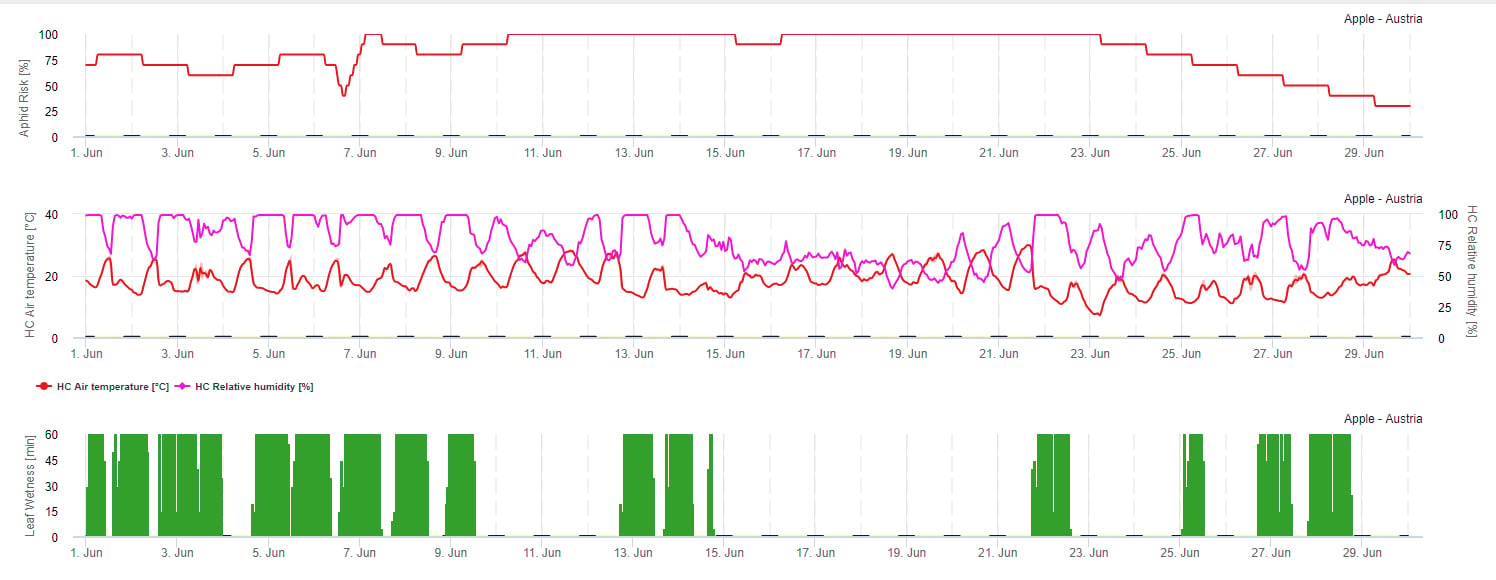
Modelo de riesgo de propagación
La propagación de los áfidos depende principalmente de la temperatura. Cuantas más horas estén cerca de la temperatura óptima (20-32°C) y haya una fuente de alimento disponible, mayor será la tasa de propagación. Los huevos y los estadios jóvenes son sensibles al aire seco. Una humedad relativa baja también puede dañar las poblaciones de pulgones (por debajo de 30% h.r.). Además de los insecticidas, los pulgones son controlados por depredadores y hongos parásitos. Los hongos parásitos dependen de la humedad. La mayoría de estas especies como Beauvaria bassiana se ven favorecidos por la humedad libre.
Aumentamos el riesgo cuando la temperatura óptima está entre 20 y 32°C y la humedad relativa entre 30 y 95% r. h. y disminuimos el riesgo cuando es demasiado húmeda (humedad de las hojas, lluvia o humedad relativa superior a 95%), demasiado fría (temperaturas inferiores a 20°C) o demasiado cálida (temperaturas superiores a 32°C), noches húmedas (humedad de las hojas en horas nocturnas).