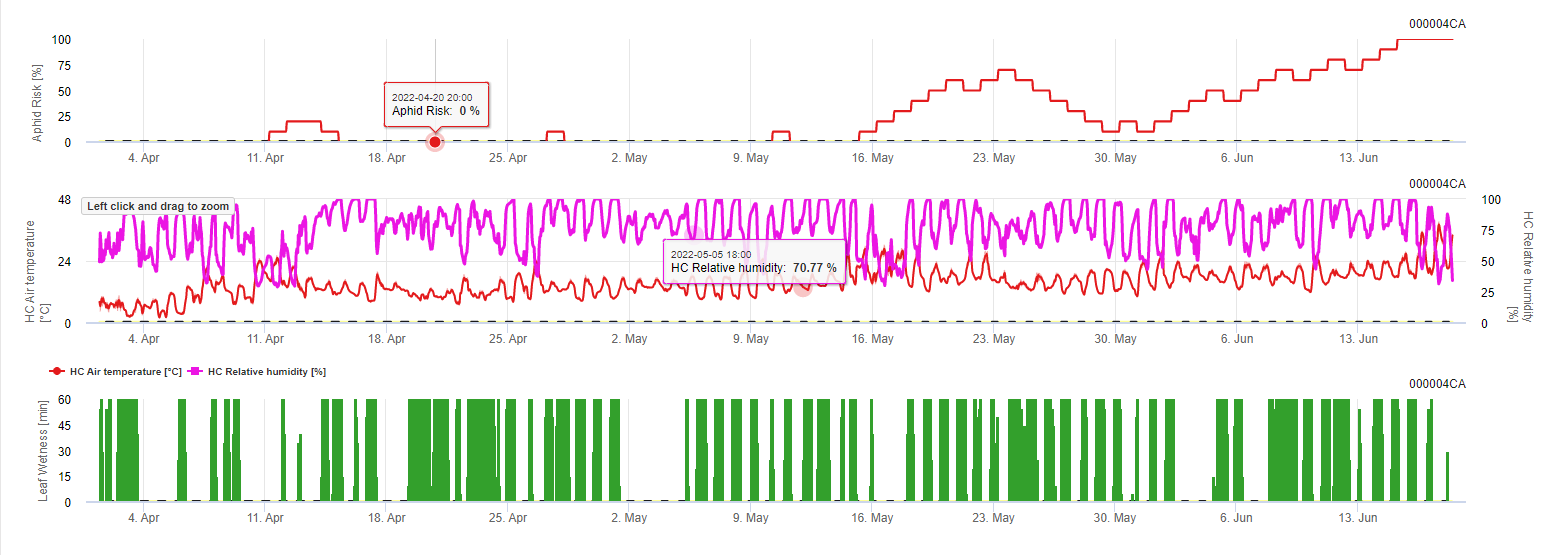
Conditions:
- in the morning when sun raises and relative humidity decreases, optimum temperatures between 20°C and 32°C – good flight is indicated.
- If temperatures are not in the optimum range (too cold/hot) or it is too wet (leaf wetness) risk decreases.
- Output is the daily risk.
So optimum temperatures and falling relative humidities during the morning are indicating a good flight day. When it is wet during the night and temperatures are to low this is bad for propagation. The same when it is hot and moist during the day.
Powdery mildew is a common disease on many types of plants. Several powdery mildew fungi cause similar diseases on different plants (such as Podosphaera species on apple and stone fruits; Sphaerotheca species on berries and stone fruits; Erysiphe necator on grapevines). Powdery mildew fungi generally require moist conditions to release overwintering spores and for those spores to germinate and infect plant tissue. However, no moisture is needed for the fungus to establish itself and grow after infecting the plant. Powdery mildews normally favour warm, Mediterranean-type climates.
Powdery mildew can be recognized easily on most plants by the white to gray powdery mycelium and spore growth that forms on both sides of leaves, flowers, fruits and on shoots. On tree fruits a rough corky spot on the skin will develop where infection occurred.

All powdery mildew fungi require living plant tissue to grow. On deciduous perennial hosts such as grapevine, raspberry, and fruit trees, powdery mildew survives from one season to the next in infected buds or as fruiting bodies called chasmothecia, which reside on the bark of cordons, branches, and stems.
Most powdery mildew fungi grow as thin mycelium layer on the surface of the affected plant part. Spores, which are the primary means of dispersal, make up the bulk of the powdery growth and are produced in chains that can be seen with a hand lens. In contrast, spores of downy mildew grow on branched stalks that look like tiny trees. Also downy mildew colonies are gray instead of white and occur mostly on the lower leaf surface.
Powdery mildew spores are carried by wind to host plants. Although humidity requirements for germination vary, many powdery mildew species can germinate and infect in the absence of water. In fact, spores of some powdery mildew fungi are killed and germination and mycelial growth are inhibited by water on plant surfaces. Moderate temperatures and shade are generally the most favorable conditions for powdery mildew development, since spores and mycelium are sensitive to extreme heat and direct sunlight.
This fungus overwinters as mycelia inside the budscales, primary infection occurs as leaves emerge from these infected buds. Secondary infections occur when conidia produced by primary and subsequent secondary infections are blown or splashed by rain onto susceptible tissues. Fruit (before pit hardening) and succulent terminal growth are susceptible to infection.
The average minimum, optimum, and maximum temperatures for S. pannosa are about 5°, 24° and 24°C. Many more conidia are formed in dry air than in humid air at all temperatures (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli; 1954)
Powdery mildew is common under similar relative humidity and temperatures as cherry powdery mildew.
Fieldclimate Model: Fungal disease is modeled by the factors temperature and duration of leaf wetness. For example on May 11th the leaf wetness period under moderate temperatures supported the development of the disease and a risk of 100% could be determined.
Literature:
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954): Temperature relations of Powdery Mildews. HILGARDIA. A Journal of Agricultural Science Published by the California Agricultural Experiment Station. University of California. Volume 22/Number 17.
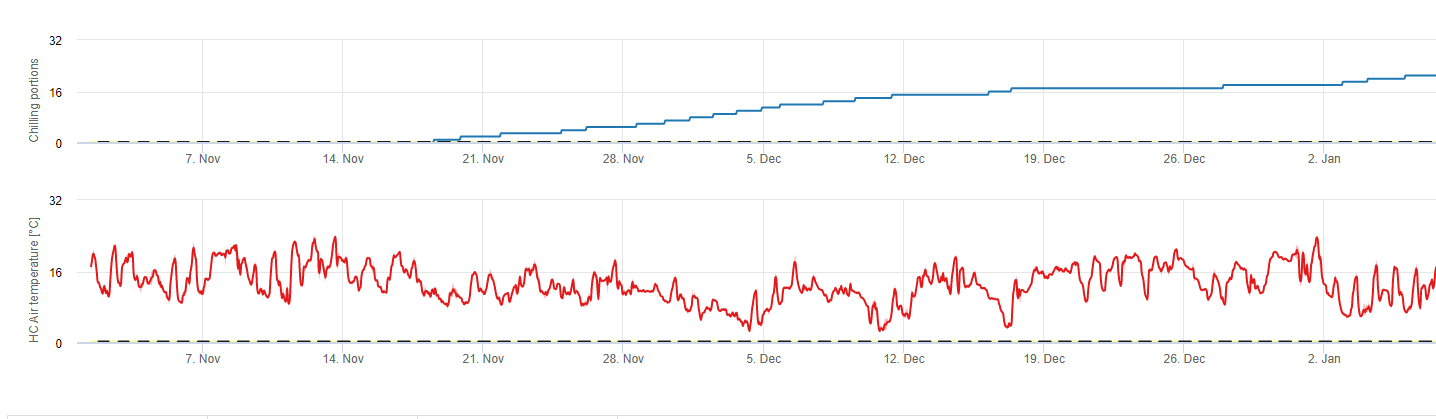
Chilling
Stone fruit trees develop their vegetative and fruiting buds in the summer and, as winter approaches, the already developed buds go dormant in response to both shorter day lengths and cooler temperatures. This dormancy or sleeping stage protects these buds from oncoming cold weather. Once buds have entered dormancy, they will be tolerant to temperatures much below freezing and will not grow in response to mid-winter warm spells. These buds remain dormant until they have accumulated sufficient chilling units (CU) of cold weather. When enough chilling accumulates, the buds are ready to grow in response to warm temperatures. As long as there have been enough CUs the flower and leaf buds develop normally. If the buds do not receive sufficient chilling temperatures during winter to completely release dormancy, trees will develop one or more of the physiological symptoms associated with insufficient chilling: 1) delayed foliation, 2) reduced fruit set and increased buttoning and, 3) reduced fruit quality.
Insufficient Chilling Symptoms
Delayed Foliation:
A classic symptom of insufficient chilling is delayed foliation. A tree may have a small tuft of leaves near the tips of the stems and be devoid of leaves for 12 to 20 inches below the tips. Lower buds will break eventually but full foliation is significantly delayed, fruit set is reduced, and the tree is weakened. Furthermore, heavy suckering from lower parts of the tree causes management problems, and normal development of next year’s fruit buds can be impaired.
Reduced Fruit Set and Buttoning:
Flowering, in response to insufficient chilling, often follows the pattern seen with leaf development. Bloom is delayed, extended, and due to abnormalities in pistil and pollen development, fruit set is reduced. In many peach cultivars, flowers drop before or around shuck split, but in others such as ‘Jersey Queen’ and ‘Harvester’, buttons form. Buttons result from flowers which apparently have set but never develop into full-size fruit. The fruit remains small and misshapen as they ripen. If you cut these fruit open, the seed is dead. Because buttoning is not apparent early in the season, growers can not thin off the abnormal fruit and the developing buttons serve as a food source and overwintering site for insects and diseases.
Reduced Fruit Quality:
The effects of insufficient chilling on fruit quality are probably the least discussed but appear to be very common especially in central and south Texas. The effects on leaf growth and fruit set are dramatic but the effects of insufficient chill on fruit quality are subtle and can occur when other symptoms do not. Insufficient chilling will cause many cultivars to have an enlarged tip and reduced firmness. Furthermore, fruit ground coloration may be greener than usual, possibly due to the fruit losing firmness before the ground color can fully change from green to yellow. The extent of these quality problems depends on the cultivar and the degree of chilling deficiency.
Models
There are various models used to calculate chilling, each one defining what a chilling unit is. The three most common models are the number of hours below 45 degrees F (7°C) model, the number of hours between 32 and 45 degrees F (2 and 7°C) model, and the Utah model. The first two models are simple and define a chilling unit as one hour below or between certain temperatures. The Utah method is more complex because it introduces the concept of relative chilling effectiveness and negative chilling accumulation (or chilling negation).
In fieldClimate.com we use the model for calculation of chill portions (CP). Chilling accumulations are calculated as chill portions, using a temperature range from 2 to 7°C. Calculations of chill proportions end after 96 hours of equal or more then >15°C ‘(it holds between7 and 15°C)
Calculations are based on the work of Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276: 165-174.