





Cherry disease models
The most important disease in cherries is brown rot, caused by the fungal pathogen Monilia laxa. Another important disease, also occurring by high moisture conditions is the disease shot hole, caused by the fungal pathogen Wilsonmyces carpophilu and the disease of cherry leaf spot, caused by Blumeriella jaapii.
Rain accumulation
Intense rain will wash off the pesticides residuing on the leaves of vines or other plants. There has been a big improvement of the rain hardness of modern fungicides since 1980. Actually we can expect the most modern fungicide to resist up to 30 mm of rain if they had a chance to dry on the leaf. If the rain was starting immediately after the spray or trough the spraying the rain resistance might be widely reduced.
Old fashioned formulations of contact fungicides we have to expect a rain hardness of less then 12 mm. Like we were used to it during the 1970th. To wet the leaves in a vineyard it needs approximately 2 mm or rain. Therefor in this mdoule we only accumulate rains with are bigger than 2mm within one leaf wetness period. This means there might be in total 6 mm of rain during a single day, but this module is not accumulating any of it because the leaves has got dry again before it was raining 2 mm.
Rain is accumulated for 3, 5 and 7 days. Over longer period plant growth is much more important for the effect of contact fungicides than rain resistance of the compounds.
Stone fruit scab model
Stone fruit scab is induced by the plant pathogenic fungus Cladosporium carpophilum. The pathogen occurs on peaches, nectarines, apricots, and plums, while losses are generally greater on peaches than on the other fruits.
The disease affects twigs, leaves and fruits. The most serious damage results from fruit infections.
Symptoms
Fruit lesions start as small, round, greenish spots. These spots generally don’t appear until the fruit is half grown even though infection occurred earlier in the season (about six to seven weeks after petal fall). Older lesions are approximately 1/4 inch in diameter and develop a dusty or velvety green appearance. The numerous lesions typically are clustered near the stem end of the fruit (this site is exposed to the sun). Extensive spotting can result in fruit cracks, which serve as entrance points for several fruit rotting fungi. Fruits may also drop prematurely or could not be stored well.
Leaves could also be infected. Small, round and yellowish- green spots occurr on the undersite of the leaf. The plant tissue may dry und drop down, leaving shot- holes. In a rainy season the infected leaves usually drop early.
On twigs cankers begin as small, reddish lesions on current season’s growth. These cankers expand slowly and may not be visible until mid summer. The small cankers have irregular margins, but do not cause sunken areas on the bark.
On the twigs the mycelium (or conidia) hibernates in the form of dark-brown spherical cells. From overwintering mycelium, conidia are produced in the spring, and the latter are carried to the leaves and fruits by wind or by rain. The conditions which favor disease development are temperatures above 16°C for spore production, over 10°C (optimal 22°C to 27°C) for spore germination, and between2°C and 35°C for disease development. Germination and penetration into the plant tissue follows shortly. Inoculations and infections continue to take place until about one month before the fruit matures. As the fungus grows on the fruit the mycelium attaches itself closely to the surface between the hairs, forming a mat of short, plump cells which give rise to conidiophores and conidia. The flesh of the peach is not penetrated, but the close contact of the fungus with the outer cells allows absorption of nutrition from the fruit through the unbroken walls. Evidently there is some injury to the outer cells.
In FieldClimate the risk of a Caldosporium carpophilum infection is determined by wet conditions during spring and early summer after petal fall. The disease is usually more serious in low- lying, shady and moist areas with low air movement.
In FieldClimate we determine infections within a temperature range of 7 to 24°C, with a temperature optimum around 20°C.
The FieldClimate Modell calculates in dependence of leaf wetness duration and temperature a risk model of Cladosporium carpohilum.
Shot hole
Shot hole is caused by the fungal pathogen Wilsonmyces carpophilu (Coryneum blight).
Most severe on apricots, but occurs on all stone fruit. The fungus infects the leaves, twigs and fruits.
Symptoms
Infected leaves show small brown spots with reddish margins (about 1 mm diameter), these spots expand to larger circular lesions (about 3mm diameter). These spots dry and fall out of the leaf, giving a shothole appearance. The infected twigs show clear- cut brown margins with a negrotic center, which does not drop out, but ooze large amounts of gum. Further on lignification of infected twings is hindered and the lesions will grow into cankers. In severe cases premature defoliation of the tree may result.
Fruits show firstly small circular, deep purple spots. As the disease progresses, the symptoms differ according to fruit type. On apricots the spots become brown, raised and rough, giving the fruit a scabby surface. On peaches and nectarines the scabs develop into deep indentations.
Infected fruit have spots of gum and in severe cases cracks in the skin.
The shothole fungus survives in infected buds. It is able to infect leaves, stems and fruits during cold, rainy weather periodes in spring and autumn. Rain periodes to infect healthy plant organs are needed.
The fungus is able to persist several years in the cankers or buds of infected twigs. Whenever conditions are favourable it may continue to grow, even during winter time. In spring time the conidia are splashed by raind to flowers and young leaves and infect them. In unfavourable periodes (dry conditions) the conidia are still viable for several months. Rain is necessary for dispersal and humid conditions are needed for germination. The fungus is able to grow above 2°C.
Literature:
- Adaskaveg JE, Ogawa JM, Butler EE (1990) Morphology and ontogeny of conidia in Wilsonomyces carpophilus, gen. nov., and comb. nov., causal
pathogen of shot hole disease of Prunusspecies. Mycotaxon 37, 275–290. - Ashkan M, Asadi P (1971) Shot hole of stone fruits in Iran. Iranian Journal of Plant Pathology 7, 39–63.Esfandiari E (1947) Les maladies des plantes cultivées et des arbres
fruitiers des régions subtropicales du nord de l’Iran. Entomologie et Phytopathologie Appliquées 5, 2. - Grove GG (2002) Influence of temperature and wetness period on infection of cherry and peach foliage by Wilsonomyces carpophilus. Canadian
Journal of Plant Pathology 24, 40–45. - Ogawa JM, Zehr EI, Bird GW, Ritchie DF, Uriu K, Uyemoto JK (1995) Compendium of stone fruit diseases. (The American Phytopathological
Society Press: St Paul, MN)
Powdery mildew
Powdery mildew is a common disease on many types of plants. Several powdery mildew fungi cause similar diseases on different plants (such as Podosphaera species on apple and stone fruits; Sphaerotheca species on berries and stone fruits; Erysiphe necator on grapevines). Powdery mildew fungi generally require moist conditions to release overwintering spores and for those spores to germinate and infect plant tissue. However, no moisture is needed for the fungus to establish itself and grow after infecting the plant. Powdery mildews normally favour warm, Mediterranean-type climates.
Powdery mildew can be recognized easily on most plants by the white to gray powdery mycelium and spore growth that forms on both sides of leaves, flowers, fruits and on shoots. On tree fruits a rough corky spot on the skin will develop where infection occurred.
All powdery mildew fungi require living plant tissue to grow. On deciduous perennial hosts such as grapevine, raspberry, and fruit trees, powdery mildew survives from one season to the next in infected buds or as fruiting bodies called chasmothecia, which reside on the bark of cordons, branches, and stems.
Most powdery mildew fungi grow as thin mycelium layer on the surface of the affected plant part. Spores, which are the primary means of dispersal, make up the bulk of the powdery growth and are produced in chains that can be seen with a hand lens. In contrast, spores of downy mildew grow on branched stalks that look like tiny trees. Also downy mildew colonies are gray instead of white and occur mostly on the lower leaf surface.
Powdery mildew spores are carried by wind to host plants. Although humidity requirements for germination vary, many powdery mildew species can germinate and infect in the absence of water. In fact, spores of some powdery mildew fungi are killed and germination and mycelial growth are inhibited by water on plant surfaces. Moderate temperatures and shade are generally the most favorable conditions for powdery mildew development, since spores and mycelium are sensitive to extreme heat and direct sunlight.
This fungus overwinters as mycelia inside the budscales, primary infection occurs as leaves emerge from these infected buds. Secondary infections occur when conidia produced by primary and subsequent secondary infections are blown or splashed by rain onto susceptible tissues. Fruit (before pit hardening) and succulent terminal growth are susceptible to infection.
The average minimum, optimum, and maximum temperatures for S. pannosa are about 5°, 24° and 24°C. Many more conidia are formed in dry air than in humid air at all temperatures (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli; 1954)
Powdery mildew is common under similar relative humidity and temperatures as cherry powdery mildew.
Literature:
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954): Temperature relations of Powdery Mildews. HILGARDIA. A Journal of Agricultural Science Published by the California Agricultural Experiment Station. University of California. Volume 22/Number 17.
Taphrina leaf curl
Peach leaf curl (fungal pathogen: Taphrina deformans) is a fungus disease that can cause severe early defoliation and crop loss on nearly all peach and nectarine cultivars.
Symptoms
The most common and striking symptom of leaf curl occurs on the leaves (foliage). Infected leaves are severely deformed and often display a variety of colors (light green and yellow to shades of red and purple). The fungus causes the meristematic cells at leaf margins to proliferate quickly and randomly, which results in the leaves becoming variously wrinkled, puckered, and curled (photo 2). As these infected leaves mature, naked asci containing ascospores of the pathogen are produced on the surface giving them a dusty appearance, after which the leaves turn brown, shrivel, and drop from the tree.
Many infected fruits drop early and go unnoticed; those that remain may become crooked at the stem end like a small yellow squash, while others develop reddish to purple and have “wart-like” deformities on the surface.
Disease cycle
The pathogen occurs commonly almost wherever peaches are grown. The fungal pathogen overwinters as conidia (blastospores, “hyphal like” spores) in protected sites in the bark and around the buds. Primary infections occur during the early spring. Starting when the buds swell until the first leaves appear from the buds. Infections on young peach leaves occur at temperatures of 10°C to 21°C. Few infection occurs below 7°C. Infections appear mainly when rain wash the overwintered spores into the buds and cold temperatures lengthen the development time of the leaves (they are exposed for a long time to the pathogen before they are fully expanded and are able to resist the penetration of the fungus). If temperatures after bud swelling are warm and leaves develop quickly, infections rarely become established, even when spring rains occur. Wetness from rain (or other factors) for over 12,5 hours are needed for leaf infection but only when the temperature is below 61°F (is 16°C) during the wet period. Maximum infection occurs when trees are wet for 2 days or more, a frequent occurrence west of the Cascades. Although infected, symptoms may not appear if temperatures remain above 69°F (21°C). Fruit are susceptibel after petal fall until air temperature remains above 19°C. Rainfall of 0,5 inch and wetness of 24hours are needed for fruit infection.
The risk of a Taphrina defomans infection is calculated in FieldClimate in two ways:
- using temperature values (old model).
- using rain accumulation during the last consecutive hours and temperature during that time period. Further on this model includes also incubation time (time, when symptoms are seen in the field) using temperature below 19°C for calculations.
Both models are available in FieldClimate under “Taphrina Leaf Curl disease”.
Leaf spot disease
Cherry leaf spot is caused by the fungus Blumeriella jaapii. The disease reduces flowering and weakens the tree. Cherry leaf spot often defoliates the tree by midsummer. Repeated defoliation makes the tree more susceptible to winter injury and may eventually kill it.
Symptoms
Leaves show small purple to brown spots with definite boundaries in early summer. The leaves turn yellow and fall. In July, the centers of the infected spots frequently fall out, giving a shot-hole appearance.
Cause
The fungus overwinters on fallen leaves. In spring following wet weather, spores form and winds blow them to infect leaves. When temperatures are favorable (16°C to 24°C), infection requires a wet period of only a few hours. Leaves are susceptible when fully unfolded — this usually occurs near petal fall. The initial leaf infections form spots, and more spores are produced in the spots. These spores are splashed by rain to other leaves and infect them. Secondary spread and infection by spores continues repeatedly, whenever wet warm weather occurs, until leaves fall in autumn.
Life cycle
Cherry leaf spot is caused by the ascomycete fungus Blumeriella jaapii (formerly known as Coccomyces hiemalis) anamorph Phloeosporella padi (Lib.) Arx. This fungus overwinters in dead leaves on the ground as its main form of survival. In the spring, apothecia develop on these leaves. Ascospores are produced in these fungal fruiting bodies and are forcibly ejected during rainy periods for about 6–8 weeks, starting at petal fall. This is the telemorph or sexual stage and results in new genetic recombination and variation for the disease. This pathogen mainly affects and is most noticeable on the leaves of cherry trees. The most obvious symptom of this disease is the yellowing of leaves before they drop in the early spring. Infected leaves are filled with numerous black lesions that cover an obvious portion of the leaf. These lesions first appear as small purple spots which turn red to brown and then black. Most spots are circular but can amalgamate to form large, irregular dead patches. These spots tend to enlarge and separate from healthy tissue. Lastly, they drop out of the leaf giving a “shot-hole” appearance. This shot hole effect is more common on sour than on sweet cherries. In more severe and advanced cases, lesions are found on the fruit, petioles, and stems. Leaves with lesions present typically fall off early in the growing season and it is common for seriously affected trees to be defoliated by midsummer. Leaves severely infected by the disease turn very yellow before falling off.
If the disease is not controlled and allowed to persist for several years there could be various adverse effects to the overall cherry production of a tree primarily due to the cycle of early season and repeated defoliation. These effects include dwarfed and unevenly ripened fruit which has poor taste, trees more susceptible to winter injury, death of fruit spurs, small and weak fruit buds, decreased fruit size and set, and eventual death of the tree. If the disease has significantly infected the leaves enough in early spring it is possible that the fruit will fail to mature in that year. This fruit will typically remain light colored, soft, and very low in soluble solids and sugar.
Disease Cycle
Primary infection
This fungus overwinters in dead leaves on the ground. In early spring (about petal fall), fungal fruiting bodies called apothecia develop in these leaves. Spores (ascospores) are produced in the apothecia and are forcibly discharged starting at petal fall. These ascospores are spread by wind or rain to the healthy, new, green leaves and thus serve as the primary inoculum in a new growing season. The ascospores can stick to the leaf and if a film of water and optimal temperature is present, germinate within hours of adhesion. Upon germination, the ascospores can enter the leaf through the stomata (natural openings) on the underside of the leaf. Ascospores cannot enter immature stomata very effectively. These ascospores grow in the intercellular spaces in the leaf. Incubation time will vary depending on the amount of available moisture and temperatures. The small purple lesions can appear within 5 days if there are damp conditions and the temperature remains steady between 60 (15°C) and 68 degrees F (20°C). This incubation period could take as long as 15 days though if lower temperatures and drier conditions occur. Generally leaves become less susceptible to the disease as they grow older during the course of the growing season. The first sign of disease may be on the suckers close to the ground. This is considered to be the primary reproduction cycle of cherry leaf spot. However, infection from this primary cycle is fairly low but is essential for the pathogen to be able to produce secondary spores.
Secondary infection
Once these lesions from the primary infection have formed, there is the potential for secondary infections. Secondary or summer spores called conidia will form on the underside of the leaf from slightly concaved eruptions called acervuli. These conidia are horn-shaped and highly water-soluble. The spores have a whitish appearance on the underside of the leaf. They are spread via wind or rain when broken open and can cause additional infections. Each of these spores can multiply and cause thousands of additional conidia to be produced in a relatively short amount of time. All of these conidia hold exactly the same genetic information and is simply the fungus just replicating its DNA. At this stage the fungal infection overwhelms the host plant and causes the plant to shed its leaves prematurely. The leaf litter produced then becomes the overwintering site of this fungus, and the cycle will begin the next spring. These secondary infections allow the disease to escalate into an epidemic very quickly.
Environment
The disease is generally distributed throughout the U.S., Canada, and various parts of the world where humid conditions occur. The cherry leaf spot fungus prefers moderately wet conditions, with temperatures above 60 degrees Fahrenheit (15°C) . Optimal temperature range for the spread of this fungus is between 60 (15) to 68 degrees Fahrenheit (20°C). Serious infection of a tree occurs in years with many rainy periods and cooler summers. Very few ascospores will be ejected from the apothecia if the temperature is less than 46 degrees F (7°C). This disease thrives in areas that do not use any type of sanitation practice as a preventative measure to prevent disease spread. When selecting a site for an orchard, use areas that have well drained soils, lots of sunlight, and good air circulation.
Modelling the infection by Blumeriella jaapii
The Ascopsore maturation Model: Under optimal Temperatures Ascospores will get mature within a 2 week period. Optimal temperatures are above 16°C. In least optimum conditions the period of ascospore discharge can take up to 6 weeks. The model is showing the period of possible ascospore discharge. During this period we have to expect primary infections of the disease.
The Ascospore discharge Model:
Ascospore discharge is depending on wetting of the apothecia and the following drying up. It is bigger when more apothecia are wetted (heavier rain or more rain) and when the temperature during and after the rain are higher. It will start at temperatures of 16°C.
The Infection model for Blummeriella jaapii:
Infection is predicted with the table by Eisensmith and Jones (Table 1), which uses leaf wetness duration and air temperature data. In the model, a wetting period is initiated when the leaf wetness sensor becomes wet. The wetting period ends when the RH drops below 90% and the leaf wetness sensor is dry. However, if the leaf wetness sensor again becomes wet within 8 hrs of the time the RH fell below 90%, it is considered to be the same wetting period with the dry hrs added as well. A wetting period may result in no infection or infection by the cherry leafspot pathogen.
Use of the model:
The cherry leaf spot model can be used as a guide to determine the need for eradicative sprays if a protectant coverage was weak during a rain episode. The model should not be used as a routine guide for eradicative spray applications- a protectant spray program is the most reliable approach for control of cherry leaf spot. The model is useful for comparing predicted leaf spot infection activity between years.
Ascospore and Conidia Infection Model based on:
- EISENSMITH, S.P. and A. L. Jones (1981): Infection Model for timing fungicide applications to control cherry leaf spot. Plant Dis. 65
- Eisensmith, S. P. and A. L Jones. 1981. A model for detecting infection periods of Coccomyces hiemalis on sour cherry. Phytopathology.71:728-732.
- Jones, A. L. and T. B. Sutton. 1996. Diseases of tree fruits in the east. Northcentral Regional Bulletin 45, Michigan State University
- Shane, B. (2011): About Enviro-weather’s Cherry Leaf Spot Report, see it here.
Monilia spp.
Brown rot, caused by Monilia spp. (Monilia laxa, Monilia fructigena and Monilia fructicola) belong to the most destructive diseases on stone fruits through Europe. Especially in cherry production the economic impact is significant.
Symptoms
Symptoms of the brown rot disease are the blight of the cherry blossom as well as the green tip of twigs due to the penetration of the pathogen into the open blossom through the stigma of pistils or anthers. This usually results in wilting of the whole part of an one-year-old twig. The leaves start to hang down, later they become brown and rigid, but usually do not fall down on the soil, they remain on the tree until the spring of the next year. Sometimes, especially under humid conditions droplets of gum are visible, which are symptoms of colonisation of the fungus as well as the established cankers. The infected fruits are covered by putrefactive spots, from which warty sporodochia (hyphe) with conidia of the “summer” form appear. Additionally, in late autumn and winter, the fungus produces sporodochia of the “winter” form on infected twigs. With time, severely affected fruits become mummified. The mycelium growing in such mummies gradually aggregates into sclerotia. Such fruits remain on the tree during winter.
Temperature and wetness duration are important environmental factors, determining the infection incidence of M. laxa on sweet cherry blossoms. Monilia laxa is well adapted to the relatively low temperatures during spring and cause infections at temperatures as low as 5°C within a very short period of wetness duration. The infection of the active bloom trough the stima does not need very much leaf wetness. Leaf wetness is only needed for germination of the conidia. Therefore infection of the young fruit needs longer leaf wetness periods. To infect the young fruit an appressoria has to be formed and free moisture is needed to build up the pressure to form the infection peg to enter the epidermis cell. With maturity of the fruit small scars on the fruits allow an infection without infection peg again and the needed leaf wetness duration becomes shorter again.
No sporulation is possible if relative humidity does not overcome 85% rel. humidity. The M.laxa model in fieldclimate.com calculates weak and severe infections. Weak infections are modelled for conditions to infect highly susceptible stages like flowers and wounded fruits close to maturity. To infect this tissue very short leaf wetness periods have shown to be sufficient. Several studies, for example:” Phenological Analysis of Brown Rot Blossom Blight of Sweet Cherry Caused by Monilinia laxa” (L. Tamm, Chr. E. Minder, and W. Flickiger; 1994) or “Effects of wounding, fruit age and wetness duration on the development of cherry brown rot in the UK.” ( X.-M. Xu*, C. Bertone and A. Berrie ;2003) confirmed low wetness periods needed for infections. Further on, immature fruits are more resistant for infections, but close to maturity they are becoming mor susceptible and conditions for severe infections are given.
Springtime dispersal patterns of Monilinia laxa conidia in apricot, peach, prune and almond trees. Canadian Journal of Botany (1974), 52: 167-176
Fieldclimate Modeling: It is probably that the time, needed for infection during bloom has to be shortened. Therefore the model is shorting infections down in the area of 2000 to 4800 degree hour above 5°C.
Western flower thrips
Western flower thrips are minute insects, about 0.03 inch long, with two pairs of fringed wings. The adult has three color forms that vary in abundance depending on the time of year. There is a pale form that is white and yellow, except for slight brown spots or blemishes on the top of the abdomen; an intermediate color form with an orange thorax and brown abdomen; and a dark form that is dark brown. The intermediate form is present throughout the year, but in spring the dark form predominates while the pale form is most abundant at other times throughout the year.
First-instar nymphs are opaque or light yellow, turning to golden yellow after the first molt. The nymphal stage lasts from 5 to 20 days.
Damage
Nymphs hatch and feed in numbers on fruits, often under the drying calyx or flower parts. Their feeding scars the surface of the fruit. These scars enlarge as the fruit grows, and may cause fruit deformity. Thrips can also cause silvering just before fruits mature.
Although some feeding does take place on blossoms, little damage results until fruit forms. Thrips can damage terminal shoots and cause them to stop growing. Usually one to two small dead leaves cling to the terminal. Buds just below the terminal grow, giving the branch a bushy appearance.
Management
Western flower thrips overwinter as adults in weeds, grasses, alfalfa, and other hosts, either in the orchard floor or nearby. In early spring, if overwintering sites are disturbed or dry up, thrips migrate to flowering trees and plants and deposit eggs in the tender portions of the host plant, e.g. shoots, buds, and flower parts.
Cultural Control
Thrips are often attracted to weeds blooming on the orchard floor. To prevent driving thrips into the trees, do not disc the cover crop when trees are in bloom. Open, weedy land adjacent to orchards should be disced as early as possible to prevent thrips development and migration of adults into orchards.
Organically Acceptable Methods
Cultural controls, clean cultivation, and sprays of the Entrust formulation of spinosad are organically acceptable tools.
Monitoring and Treatment Decisions
Begin monitoring thrips as trees begin to bloom. Monitor for thrips by examining blossoms from trees by slapping a shoot with five to ten blossoms against a yellow card or look for the immature stages within the blossoms. Often nymphs are not dislodged by the slapping method so also dissect individual flowers and examine them with a hand lens for nymphs. First instar nymphs are white in color and often difficult to see, so be sure to check carefully. Check a minimum of 50 trees per orchard for nymphs. In warm springs, adults will often migrate in and out of a block without being detected so it is important to always sample for nymphs.
If two or more adult thrips are present or if any nymphs are found, a treatment is warranted. If a treatment is applied, make it before the calyx becomes tight around the developing ovary. If nymphs are found under the jacket after it tightens around the fruit, use methomyl.
Source: Literature from UC IPM Davis:
- K. R. Day, UC Cooperative Extension, Tulare County
- K.Tollerup, UC IPM Program, Kearney Agricultural Center, Parlier
In FieldClimate we use following conditions for calculation of the risk model:
Risk model for The western flower thrips Frankliniella occidentalis (Pergande) based on daily values:
- Risk increases, when air temperatures higher than 18°C and smaller than 32°C and relative humidity is higher than 70% by 20%
- Risk decreases, when air temperature higher than 32°C for more than 8 hours or
- If relative humidity is never higher than 70% or if all temperatures below 15°C reduce risk by 10%.
Bacterial canker
Bacterial canker is a disease of the stems and leaves of Prunus, especially plums and cherries, but also apricots, peaches and ornamental Prunus species. It causes sunken patches of dead bark and small holes in leaves.
The species P. syringae exists as a large number of pathovars, (abbreviated to pv.), so-called because although all look the same, they have different, specific hosts. The pathovar morsprunorum is restricted to Prunus species, pv. syringae has a much wider host range, but both cause similar symptoms on Prunus.
The bacteria exist as surface dwellers (epiphytes) on leaves and, during wet weather in spring or early summer, can enter through the leaf pores (stomata), causing infections to develop in the young leaves. As the leaf matures these infections cease to expand and are revealed as small patches of dead tissue. As the leaf expands fully, the live tissues pull away from the dead patch, which drops out, leaving a ‘shothole’.
Cankers develop when the bacterial cells gain entry through wounds or leaf scars at the time of leaf fall. Cankers remain more or less dormant through summer, when tissues are resistant, and during autumn and winter when temperatures are low. In spring, the infections spread rapidly, killing the bark.
Symptoms
You may see the following symptoms:
X) On stems and spurs: Sunken, dead areas of bark develop in spring and early summer, often accompanied by a gummy ooze. If the infection spreads all round the branch it will die rapidly. However, it should be noted that gum production (gummosis) from the bark of Prunus species is actually quite common, and in the absence of dead, sunken bark is likely to have resulted from causes other than bacterial canker, for example physical damage or environmental stresses.
On leaves: Small brown spots appear which are often round and fall out later to leave holes – as if the leaf had been hit by shotgun pellets, leading to the popular name of ‘shothole’.
Non-chemical control
Where possible, carry out all pruning in July or August when tissues are most resistant. This is also the best time to prune in order to minimize the risk of infection by spores of the fungus causing silver leaf disease. Cut out all cankered areas, pruning back to healthy wood and paint promptly with a wound paint to protect the wound from re-infection.
Source: Royal Horticultural Society
FieldClimate model
Risk Model based on daily values:
- if soil water tension all day is smaller than 25 and rain is more than 2 mm and leaf wetness duration is longer than 6 hours risk increases by 20% else
- if soil water tension all day is higher than 40 risk decreases by 20% or
- if no leaf wetness all day risk decreases by 10% or
- if no hour with more than 70% rel humid risk decreases by 10%
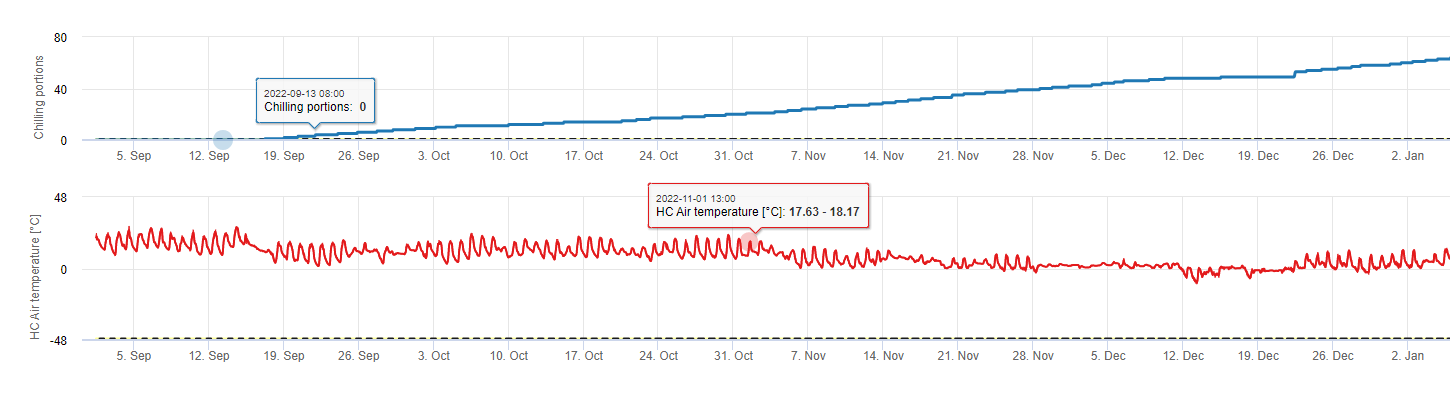
Chilling portions
Chilling
Stone fruit trees develop their vegetative and fruiting buds in the summer and, as winter approaches, the already developed buds go dormant in response to both shorter day lengths and cooler temperatures. This dormancy or sleeping stage protects these buds from oncoming cold weather. Once buds have entered dormancy, they will be tolerant to temperatures much below freezing and will not grow in response to mid-winter warm spells. These buds remain dormant until they have accumulated sufficient chilling units (CU) of cold weather. When enough chilling accumulates, the buds are ready to grow in response to warm temperatures. As long as there have been enough CUs the flower and leaf buds develop normally. If the buds do not receive sufficient chilling temperatures during winter to completely release dormancy, trees will develop one or more of the physiological symptoms associated with insufficient chilling: 1) delayed foliation, 2) reduced fruit set and increased buttoning and, 3) reduced fruit quality.
Insufficient Chilling Symptoms
Delayed Foliation:
A classic symptom of insufficient chilling is delayed foliation. A tree may have a small tuft of leaves near the tips of the stems and be devoid of leaves for 12 to 20 inches below the tips. Lower buds will break eventually but full foliation is significantly delayed, fruit set is reduced, and the tree is weakened. Furthermore, heavy suckering from lower parts of the tree causes management problems, and normal development of next year’s fruit buds can be impaired.
Reduced Fruit Set and Buttoning:
Flowering, in response to insufficient chilling, often follows the pattern seen with leaf development. Bloom is delayed, extended, and due to abnormalities in pistil and pollen development, fruit set is reduced. In many peach cultivars, flowers drop before or around shuck split, but in others such as ‘Jersey Queen’ and ‘Harvester’, buttons form. Buttons result from flowers which apparently have set but never develop into full-size fruit. The fruit remains small and misshapen as they ripen. If you cut these fruit open, the seed is dead. Because buttoning is not apparent early in the season, growers can not thin off the abnormal fruit and the developing buttons serve as a food source and overwintering site for insects and diseases.
Reduced Fruit Quality:
The effects of insufficient chilling on fruit quality are probably the least discussed but appear to be very common especially in central and south Texas. The effects on leaf growth and fruit set are dramatic but the effects of insufficient chill on fruit quality are subtle and can occur when other symptoms do not. Insufficient chilling will cause many cultivars to have an enlarged tip and reduced firmness. Furthermore, fruit ground coloration may be greener than usual, possibly due to the fruit losing firmness before the ground color can fully change from green to yellow. The extent of these quality problems depends on the cultivar and the degree of chilling deficiency.
Models
There are various models used to calculate chilling, each one defining what a chilling unit is. The three most common models are the number of hours below 45 degrees F (7°C) model, the number of hours between 32 and 45 degrees F (2 and 7°C) model, and the Utah model. The first two models are simple and define a chilling unit as one hour below or between certain temperatures. The Utah method is more complex because it introduces the concept of relative chilling effectiveness and negative chilling accumulation (or chilling negation).
In FieldClimate we use the model for calculation of chill portions (CP). Chilling accumulations are calculated as chill portions, using a temperature range from 2 to 7°C. Calculations of chill proportions end after 96 hours of equal or more then >15°C ‘(it holds between7 and 15°C)
Calculations are based on the work of Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276: 165-174.
Recommended equipment
Check which sensor set is needed for monitoring this crop’s potential diseases.