





Fusarium-Kopffäule oder Schorf bei kleinen Körnern wird verursacht durch den Pilz Fusarium graminearum (Schwabe), obwohl die Niederlande und andere Gebiete in Mitteleuropa F. culmorum als die am weitesten verbreitete Art (Snidjers, 1989). In Polen, F. culmorum, F. graminearum und F. nivale haben ähnliche mäßige bis schwere Virulenzwerte gezeigt, während F. avenaceum hat sich als mittel bis mäßig virulent erwiesen. In mehreren Studien zur Identifizierung des verursachenden Organismus wurden jedoch bis zu 18 Fusarium spp.. wurden isoliert und identifiziert (Mihuta-Grimm und Foster, 1989; Reis, 1985).
Artikel von L. GilchristH.J. Dubin Der Schorf tritt vor allem in feuchtwarmen Regionen auf, in denen die Blütezeit mit Regenperioden zusammenfällt. Das Auftreten dieser Krankheit hat in den letzten zehn Jahren aus verschiedenen Gründen zugenommen. Der vielleicht wichtigste Grund ist die Ausweitung der Anbauflächen, auf denen Weizen im Wechsel mit Mais oder anderen Getreidesorten angebaut wird. Weitere Gründe sind Änderungen des Anbausystems zum Schutz des Bodens und die Verlagerung des Weizenanbaus von traditionellen in feuchtere, nicht-traditionelle Gebiete (Gilchrist et al., 1997).
Fusarienschorf verursacht weltweit schwere Produktionsverluste, die bis zu 50 % betragen können. Auch in Paraguay begünstigten die Witterungsbedingungen in den Jahren 1972 und 1975 Fusarium- und Septoria-Epidemien, die zusammen zu Verlusten von bis zu 70 % führten (Viedma, 1989). Polen, die Niederlande, das Vereinigte Königreich, die ehemalige Tschechoslowakei, die Russische Föderation, Frankreich und Österreich sind einige der europäischen Länder, die das Auftreten von Schorf melden. Auf dem amerikanischen Kontinent sind Gebiete in Kanada, den Vereinigten Staaten, Mexiko, Guatemala, Brasilien, Ecuador, Uruguay und Argentinien stark von Fusarium betroffen (Ireta und Gilchrist, 1994).
Schorf kann zu erheblichen Ertrags- und Qualitätseinbußen sowie zu Vergiftungen bei Tieren und Menschen führen (Ireta und Gilchrist, 1994; Baht et al., 1989; Luo, 1988; Snidjers, 1989; Marasas et al., 1988). Der durch Schorf verursachte Schaden in den Vereinigten Staaten wurde 1993 auf mehr als $1 Milliarde US und 1994 auf $500 Millionen US geschätzt. In China schätzt man, dass in Epidemiejahren bis zu 7 Mio. ha von Schorf befallen werden können und 2,5 Mio. t Getreide verloren gehen können. Krankheiten im Zusammenhang mit Fusarienmykotoxinen wurden beim Menschen in China, Indien und Japan festgestellt, während bei Tieren in zahlreichen Teilen der Welt Krankheiten festgestellt wurden (Dubin et al., 1997).
Heute gibt es weltweit Vorschriften für Mykotoxine (Van Egmond und Dekker, 1995). In vielen Ländern werden die Vorschriften jedoch nicht angewandt, und viele Menschen, vor allem in ländlichen Gebieten, verzehren das Getreide ohne jegliche Kontrolle, entweder als Getreidekörner oder in indirekter Form als Fleisch von Tieren, die mit kontaminiertem Getreide gefüttert wurden. Tabelle 16.1 zeigt die wichtigsten Auswirkungen auf Schweine und Geflügel durch die wichtigsten Toxine, die von einigen Fusarium Arten.
Feuchte und warme Witterungsbedingungen zwischen dem Ährenschieben und der Reife erhöhen den Schorfbefall. Der Eintrittspunkt von F. graminearum ist die Ähre, insbesondere die Blütenorgane. Dies beeinträchtigt den Samenansatz und die Kornfüllung. Befallene Ährchen verlieren schnell Chlorophyll und werden blass. Später färben sie sich rosa oder pfirsichfarben, vor allem an der Basis und den Rändern der Hüllspelzen (Tafel 41). Wenn die Umweltbedingungen weiterhin günstig sind, breitet sich die Infektion auf die benachbarten Ährchen aus und kann in einigen Fällen die gesamte Ähre einschließlich der Spindel und des Stiels befallen. Bei schwerem Befall sind die beschädigten Körner mit Myzelien bedeckt und sehen aus wie eine rosafarbene, watteartige Masse. Bei mäßigem Befall können die Körner schrumpelig sein, ein geringes Gewicht aufweisen und weißlich gefärbt sein (Grabsteinkörner).
Die wichtigsten toxischen Wirkungen der wichtigsten Toxine einiger Fusarium-Arten auf Schweine und Geflügel
| Mykotoxin | Klinische Anzeichen |
| Zearalenon | Geschwollene rote Vulva; Vaginalprolaps bei Schweinen; Saugferkel können vergrößerte Vulva aufweisen; Fruchtbarkeitsstörungen |
| Vomitoxin (Desoxynivalenol, DON) | Verminderte Futteraufnahme und Gewichtszunahme bei Schweinen mit DON >2 mg/kg Futter; Erbrechen; Futterverweigerung bei sehr hohen DON-Konzentrationen (>20 mg/kg Futter)a |
| Andere Trichothecene T-2-Toxin HT-2-Toxin Diacetoxyescirpenol> | Giftiger als DON; verringerte Futteraufnahme; Erbrechen; Haut- und Magen-Darm-Reizung; Neurotoxizität; abnorme Nachkommen; erhöhte Krankheitsanfälligkeit; Hämorrhagie |
| Ocharatoxin | Befall vor allem der proximalen Tubuli der Nieren bei Schweinen und Geflügel; die Nieren sind stark vergrößert und blass; Fettleber bei Geflügel |
amg/kg = Teile pro Million (ppm).
Quelle: Trenholm et al., 1984.
Fusarium graminearum (Tafel 42) kann Weizenpflanzen in allen Wachstumsstadien befallen und Sämlings-, Stengel- und Wurzelfäule verursachen. Primärinfektionen können entweder von Ascosporen oder Makrokonidien ausgehen, die auf Spelzen und ausgefahrenen Staubbeuteln abgelagert werden. Temperaturen von 10° bis 30°C und eine relative Luftfeuchtigkeit von über 95 % für 40 bis 60 Stunden reichen in der Regel aus, damit die Makrokonidien die Ähren erfolgreich infizieren können (Ireta, 1989).
Fusarium graminearum ist eine der wenigen Fusarium Art, die unter Feldbedingungen Perithecien produziert (Tafel 43, Tafel 44). Perithecien stellen das sexuelle Stadium des Pilzes Gibberella zeae dar und werden auf Weizenspelzen gebildet. Perithezien spielen eine wichtige Rolle für das Überleben des Erregers von Jahr zu Jahr (Khonga und Sutton, 1988) und koexistieren mit Myzelien in den Rückständen der vorherigen Ernte, um die anfängliche Inokulumquelle für Schorf zu bilden. In China durchgeführte Untersuchungen haben gezeigt, dass die niedrigste Temperatur für die Bildung von Perithecien 7° bis 10°C und die günstigste 15° bis 20°C beträgt. Der niedrigste Feuchtigkeitsgehalt des Bodens für die Bildung von Perithecien lag bei 50 bis 60 %, der geeignetste bei 70 bis 80 % (Wang, 1997).
Zu den Faktoren, die die Krankheitsentwicklung bestimmen, gehören Klima, Inokulummenge und Wachstumsstadium des Weizens. Weizenschorfepidemien hängen hauptsächlich von der Menge des primären Inokulums und weniger vom sekundären Inokulum ab. Die Invasion erfolgt meist während der Blütezeit. Die Sekundärinfektion mit Konidien (Tafel 45) erfolgt nach dem Erscheinen der kranken Ähren auf dem Feld.
Ernterückstände und kulturelle Praktiken spielen eine wichtige Rolle bei der Erhaltung von F. graminearum und infolgedessen Epidemien. Infektionen auf Weizen, der in ein Feld mit Maisrückständen gesät wurde, können zwei- bis dreimal so schwerwiegend sein (Teich und Nelson, 1984). Im Allgemeinen war der Erreger nur saprophytisch auf Reisstoppeln in den Reis-Weizen-Rotationsgebieten und auf Maisstängeln in den Trockenanbaugebieten Chinas (Wang, 1997). Wenn die Rückstände untergepflügt werden, sinkt die Überlebensrate der Perithecien und reduziert die primäre Inokulumquelle (Reis, 1989).
Fusarium graminearum ist ein fakultativer Parasit und pathogen für viele andere Gräser, einschließlich gängiger Unkräuter und Getreidekulturen (Roggen, Reis, Gerste und Triticale). Wenn Praktiken wie die Fruchtfolge mit Nicht-Wirtspflanzen oder die Bewirtschaftung von Ernterückständen allein nicht wirksam sind, können sie in Kombination die Quelle des primären Inokulums reduzieren. Die wirksame Krankheitsbekämpfung basiert auf einem integrierten Management, das angemessene agronomische Praktiken, die Verwendung resistenter oder toleranter Sorten und den Einsatz von Chemikalien umfasst.
Biologie der Fusarium-Kopffäule
Gelegentliche Krankheitserreger
FHB wird durch Pilzarten der Gattung Fusarium verursacht. Die häufigste Art, die FHB verursacht, ist Fusarium graminearum (sexuelles Stadium - Gibberella zeae). Dieser Pilz ist derselbe, der häufig mit der Stängelfäule von Mais in Verbindung gebracht wird. Eine weitere Fusarium Art, die FHB verursacht, ist Fusarium culmorum. Beide F. graminearum und F. culmorum kann auch Wurzelfäule bei kleinen Körnern verursachen. Bei Gerste sind zwei weitere Fusarium Arten, F. poae und F. avenaceumkann auch die Kernfäule verursachen.
Überleben und Ausbreitung
Der Pilz überdauert und vermehrt sich auf infizierten Ernterückständen von Kleingetreide und Mais. Bei feuchtem Wetter werden die Sporen des Pilzes vom Wind verweht oder auf die Köpfe der Getreidepflanzen gespritzt. Die Sporen können aus dem Inneren einer Kultur stammen oder von umliegenden Kulturen, manchmal über große Entfernungen, verweht werden. Weizen- und Hartweizenkulturen sind von der Blüte (Bestäubung) bis zum Hartteigstadium der Kornentwicklung anfällig für Infektionen. Die Sporen des verursachenden Pilzes können zur Blütezeit auf den freiliegenden Staubbeuteln landen und dann in die Körner, Spelzen oder andere Teile des Kopfes einwachsen. Bei Sommergerste, die blüht, wenn sich der Kopf im Stiefel befindet, ist die Infektion nach der Blütezeit am häufigsten, sobald der Kopf die Blattscheide durchbricht. Die Infektion kann in beiden Kulturen bis kurz vor der Kornreife andauern, wenn die Umweltbedingungen für den Organismus/die Organismen günstig sind.
Die günstigsten Bedingungen für eine Infektion sind längere Perioden (48 bis 72 Stunden) mit hoher Luftfeuchtigkeit und warmen Temperaturen (75 bis 85 Grad Fahrenheit (24°C bis 30°C)). Die Infektion erfolgt jedoch auch bei kühleren Temperaturen, wenn die hohe Luftfeuchtigkeit länger als 72 Stunden anhält. Bei frühen Infektionen können Sporen in der Luft entstehen, die für die sekundäre Ausbreitung der Krankheit verantwortlich sind, vor allem, wenn die Kultur aufgrund von Spätaustrieben eine ungleichmäßige Blüte aufweist.
Da die Entwicklung von FHB von der Blüte (Kopfaufgang bei Gerste) bis zur Kornentwicklung von günstigen Umweltbedingungen abhängt, variieren das Auftreten und die Schwere der Krankheit von Jahr zu Jahr. Eine Kombination von Faktoren, die zu den schwersten Ertrags- und Qualitätseinbußen führen können, sind: reichlich Inokulum, längere oder wiederholte Perioden von Nässe und hoher Luftfeuchtigkeit während der Blüte (Kopfaufgang bei Gerste) bis zur Kornentwicklung und die Verwendung einer sehr anfälligen Sorte.
Fusarium Head Blight Infektionsmodell
Es ist bekannt, dass die Pilzerreger des Fusarium Head Blight-Komplexes auf Weizen durch warme Temperaturen von 20 °C bis 30 °C und lange Feuchteperioden begünstigt werden. Mehrtägige Blattnässeperioden führen zu früh sichtbaren Syptomen. Symptome können aber auch nach einer längeren Latenzzeit auftreten, wenn auf die Infektion eine Blattnässeperiode von 18 Stunden oder noch kürzer folgt. Symptome können auch nach künstlicher Inokulation bei Temperaturen von 15°C auftreten.
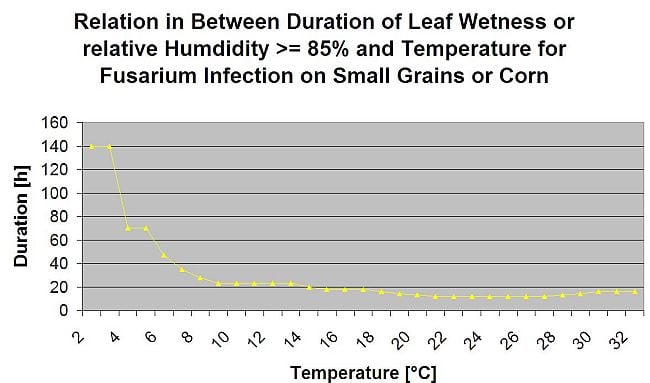

Bei der Zusammenfassung der verschiedenen Temperatur- und Feuchtigkeitskombinationen, die wir in der Literatur gefunden haben, sind wir zu dem Schluss gekommen, dass Fusarium-Kopfschimmel-Infektionen auftreten, wenn die Temperatur- und Blattnässeperioden oder Perioden mit einer relativen Luftfeuchtigkeit von mehr als 85% die in der folgenden Grafik dargestellten Werte überschreiten.
Die Infektionen werden durch einen Regen von 2 mm ausgelöst. Von einer Fusarium-Kopffieber-Infektion kann ausgegangen werden, wenn der Wert für den Infektionsfortschritt 100% erreicht. Die Berechnung dieses Fortschrittswertes folgt der Beziehung zwischen der Dauer der feuchten Bedingungen und der Temperatur, die in der obigen Grafik dargestellt ist.
Dieses Modell dient zur Visualisierung der Infektionstage und der Klimabedingungen während des Klimas. Das Wissen der Landwirte über das Entwicklungsstadium der verschiedenen Weizensorten gibt ihnen die Möglichkeit, unmittelbar nach der Infektion eine kurative Spritzung vorzunehmen.
Fusarium-Mykotoxin-Alarm
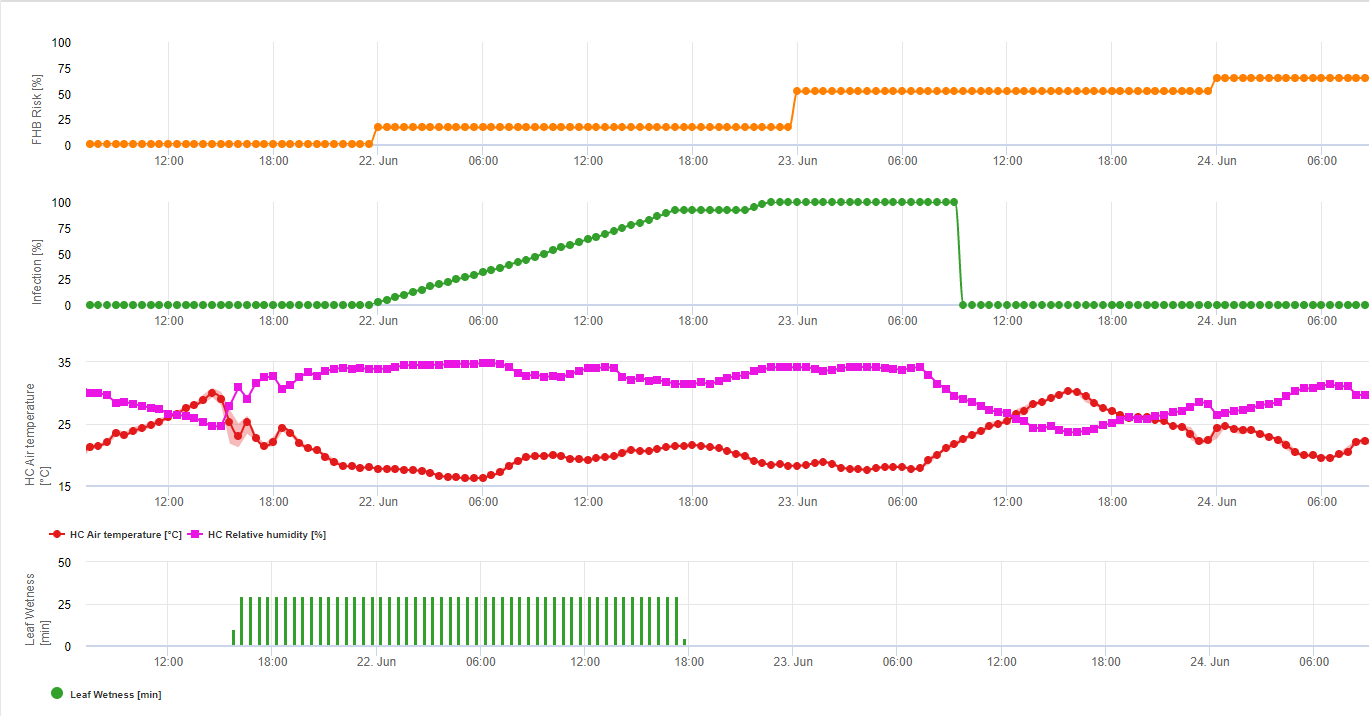
Infektionsversuche mit längeren Blattnässeperioden von Fusarium-Kopffäule führen zu hohen Mykotoxingehalten. Aus diesen Informationen geht hervor, dass eine Blattnässeperiode von 48 Stunden oder mehr während der Stadien 61 und 69 ein hohes Risiko für Mykotoxine darstellt. Die Erfahrungen bei der Analyse von DON in kommerziell angebautem Weizen haben gezeigt, dass ausreichend lange Blattnässeperioden für eine Infektion nach einer Erstinfektion in den Stadien 61 bis 69 auch die DON-Werte erhöhen können. Bei längeren Blattnässeperioden können Mykotoxine bis zum Stadium 85 ansteigen. FieldClimate akkumuliert einen Risikowert proportional zum Infektionsfortschritt für jede erfolgreiche Infektionsperiode über den Zeitraum, der für diese Berechnungen als geeignet ausgewählt wurde. 6 gerade abgeschlossene Infektionsperioden würden zu einem Risiko von 100% führen. Normalerweise ist die Blattnässeperiode, die zu einer Fusarieninfektion führt, länger als im Minimum erforderlich. Daher führen die meisten Fusarieninfektionen zu einer Risikoerhöhung von mehr als 17%. Der Risikowert, der auf eine problematische Mykotoxinsituation hinweist, hängt von der Feldgeschichte ab. Weizen, der nach Mais ohne Bodenbearbeitung oder Weizen ohne Bodenbearbeitung nach Mais ohne Bodenbearbeitung angebaut wird, kann nur ein geringes Risiko bergen, wenn er nicht zum optimalen Zeitpunkt gespritzt wird. Bei ungespritztem Weizen muss man bereits nach 35% mit erhöhten DON-Werten rechnen. Weizen nach unbestelltem Weizen, der auf eine andere Kultur als Mais oder Weizen folgt, kann ein höheres Risiko von 50% bergen. Bei Weizen nach Mais oder Weizen mit Bodenbearbeitung kann sich das Risiko auf 70% erhöhen. Weizen im ersten Jahr sollte auf DON getestet werden, wenn das Risiko über 100% liegt.
1) Fusarium Head Blight Risikomodell die die risikoreichen Zeiträume für eine Infektion angibt. Immer dann, wenn 100%-Infektionen (grüne Linie) erreicht werden, ist das Risiko (blaue Linie) sehr hoch und die Bedingungen für den Pilz waren günstig für eine Infektion. In Abhängigkeit von der Anwendungsmethode (kurativ, präventiv) wird der risikoreiche Zeitraum durch die blaue Linie dargestellt.
2) Fusarium Head Blight: In diesem Modell wird die Infektion des FHB durch Niederschlag (2 mm erforderlich), relative Luftfeuchtigkeit (über 85%) oder Blattnässe, Temperatur während des Prozesses berechnet. Wenn die Infektion 100% erreicht, sind optimale Bedingungen für den Pilzerreger gegeben. Des Weiteren berechnet das Modell das FHB-Mykotoxinrisiko.
Es gibt zwei wichtige Septoria-Krankheiten bei Weizen. Es handelt sich um die Septoria tritici-Fleckenkrankheit, die durch den Pilz ausgelöst wird Septoria tritici (Teleomorph: Mycophaerella graminicola), und Septoria nodorum Fleckenkrankheit, verursacht durch den Pilz Septoria nodorum (Teleomorph: Leptosphaeria nodorum). Beide Krankheiten verursachen schwere Ertragseinbußen, die Berichten zufolge zwischen 31 und 53 Prozent liegen (Eyal, 1981; Babadoost und Herbert, 1984; Polley und Thomas, 1991). Weltweit sind mehr als 50 Millionen Hektar Weizen betroffen, die hauptsächlich in niederschlagsreichen Gebieten wachsen. In den letzten 25 Jahren haben diese Krankheiten zugenommen und sind in bestimmten Gebieten zu einem wichtigen limitierenden Faktor für die Weizenproduktion geworden. Bei schweren Epidemien schrumpfen die Körner anfälliger Weizensorten und sind nicht mehr mahlfähig. Epidemien der Septoria tritici-Fleckenkrankheit und der Septoria nodorum-Fleckenkrankheit des Weizens stehen in Zusammenhang mit günstigen Witterungsbedingungen (häufige Regenfälle und gemäßigte Temperaturen), bestimmten Anbaumethoden, der Verfügbarkeit von Inokulum und dem Vorhandensein anfälliger Weizensorten (Eyal et al., 1987).
Septoria spp. Biologie
Nach Erick De Wolf, Septoria Tritici Blotch, Kansas State University, April 2008 Septoria tritici Fleck, der als gefleckter Blattfleck bekannt ist, wird durch den Pilz Septoria tritici. Sie ist in allen Weizenanbaugebieten der Welt verbreitet und stellt in vielen Regionen ein ernstes Problem dar. Der Septoria tritici-Fleck ist am schädlichsten, wenn die Krankheit die oberen Blätter und Köpfe anfälliger Sorten spät in der Saison befällt.
Symptome
Septoria tritici Die Fleckensymptome treten erstmals im Herbst auf. Die ersten Symptome sind kleine gelbe Flecken auf den Blättern. Diese Läsionen werden mit zunehmendem Alter oft hellbraun, und die Pilzfruchtkörper sind in den Läsionen an den Grannen eingebettet zu sehen. Die Läsionen sind unregelmäßig geformt und reichen von elliptisch bis lang und schmal (Abbildung 1). Die Läsionen enthalten kleine, runde, schwarze Sprenkel, die die Fruchtkörper des Pilzes darstellen. Die schwarzen Fruchtkörper sehen aus wie schwarze Pfefferkörner und sind in der Regel auch ohne Lupe zu erkennen. Die Krankheit beginnt an den unteren Blättern und breitet sich allmählich auf das Fahnenblatt aus. Auch die Blattscheiden sind anfällig für den Befall. In feuchten Jahren kann der gesprenkelte Blattfleckenpilz auf die Köpfe übergehen und braune Läsionen auf den Hüllspelzen und Grannen verursachen, die als Hüllspelzenfleck bekannt sind. Diese Läsionen werden mit zunehmendem Alter oft hellbraun, und die Pilzfruchtkörper sind oft in den Läsionen an den Grannen eingebettet.
Die Spelzenfäule kann zu erheblichen Ertragseinbußen führen, aber der Zusammenhang zwischen der Schwere der Krankheit und den Ertragseinbußen ist nicht genau bekannt. Septoria tritici Flecken können mit anderen Blattkrankheiten verwechselt werden, die sehr ähnliche Symptome aufweisen: Bräunungsflecken und Stagonspora nodorum Fleckenkrankheit, zum Beispiel. In der Regel sind die Pflanzen von mehr als einer dieser Blattkrankheiten befallen, und es kann eine Laboruntersuchung erforderlich sein, um genau zu bestimmen, welche Krankheiten am häufigsten vorkommen. Eine Laboruntersuchung ist fast immer erforderlich, um die Ursache der Spelzenfleckenkrankheit zu bestimmen. Die Kenntnis der Art ist für Spritzentscheidungen nicht wichtig, da alle drei Krankheiten ähnlich auf Fungizide ansprechen. Für die Sortenwahl ist es jedoch wichtig zu wissen, welche Krankheiten am häufigsten vorkommen, da die Resistenz gegen die Krankheiten von verschiedenen Genen gesteuert wird.
Die zuverlässigste Methode, den Septoria tritici-Fleck von den anderen Krankheiten zu unterscheiden, ist das Vorhandensein der schwarzen Pilzfruchtkörper. Der Pilz, der die Bräunungsfleckenkrankheit verursacht, bildet diese Art von Fortpflanzungsorganen nicht aus. Der Pilz, der den Stagonospora nodorum-Fleck verursacht, bildet jedoch unter feuchten Bedingungen hellbraune Fruchtkörper aus. Abgesehen von dem Farbunterschied sind diese Strukturen auch kleiner als die von Septoria tritici.
Lebenszyklus
Septoria tritici überlebt den Sommer über auf den Rückständen einer früheren Weizenernte und beginnt im Herbst mit der Infektion. Es gibt einige Hinweise darauf, dass der Pilz in Verbindung mit anderen Graswirten und Weizensamen überleben kann. Diese Quellen für den Pilz sind wahrscheinlich am wichtigsten, wenn keine Weizenrückstände vorhanden sind. Unabhängig von der Fruchtfolge oder der Rückstandsbewirtschaftung ist in der Regel genügend Inokulum vorhanden, um Herbstinfektionen auszulösen. Septoria tritici Fleckenkrankheit wird durch kühles, feuchtes Wetter begünstigt. Der optimale Temperaturbereich liegt bei 16 bis 21 °C; Infektionen können jedoch auch in den Wintermonaten bei Temperaturen von bis zu 5 °C auftreten. Die Infektion erfordert mindestens 6 Stunden Blattnässe, für eine maximale Infektion sind bis zu 48 Stunden Nässe erforderlich. Nach der Infektion benötigt der Pilz 21 bis 28 Tage, um die charakteristischen schwarzen Fruchtkörper zu entwickeln und eine neue Generation von Sporen zu produzieren. Die in diesen Fruchtkörpern produzierten Sporen werden in klebrigen Massen ausgeschieden und benötigen Regen, um sie auf die oberen Blätter und Köpfe zu spritzen.
Infektion durch Septoria tritici
Pyknidiensporen von S. tritici keimen in freiem Wasser aus beiden Enden der Spore oder aus Zwischenzellen (Weber, 1922). Die Sporenkeimung beginnt erst etwa 12 Stunden nach dem Kontakt mit dem Blatt. Die Keimschläuche wachsen willkürlich über die Blattoberfläche. Weber (1922) beobachtete nur die direkte Penetration zwischen den Epidermiszellen, andere Forscher kamen jedoch zu dem Schluss, dass die Penetration durch offene und geschlossene Spaltöffnungen das wichtigste Mittel für das Eindringen in den Wirt ist (Benedict, 1971; Cohen und Eyal, 1993; Hilu und Bever, 1957). Kema et al. (1996) beobachteten nur die Stomata-Penetration. Hyphen, die durch Spaltöffnungen wachsen, verengen sich auf einen Durchmesser von etwa 1 μm und werden dann breiter, wenn sie die substomatäre Höhle erreichen.
Die Hyphen wachsen parallel zur Blattoberfläche unter den Epidermiszellen, dann durch das Mesophyll zu den Zellen der unteren Epidermis, aber nicht in die Epidermis. Es werden keine Haustorien gebildet, und das Hyphenwachstum wird durch Sklerenchymzellen um die Gefäßbündel herum begrenzt, außer wenn die Hyphen sehr dicht sind. Gefäßbündel werden nicht durchdrungen. Die Hyphen wachsen interzellulär entlang der Zellwände durch das Mesophyll und verzweigen sich an einem Septum oder in der Mitte einer Zelle. Etwa 9 Tage lang treten keine makroskopischen Symptome auf, abgesehen von einer gelegentlichen toten Zelle, aber nach 11 Tagen sterben die Mesophyllzellen rasch ab. Die Pyknidien entwickeln sich in substomatalen Kammern. Die Hyphen wachsen nur selten in die Wirtszellen ein (Hilu und Bever, 1957; Kema et al, 1996; Weber, 1922).
Eine erfolgreiche Infektion erfolgt erst nach mindestens 20 Stunden hoher Luftfeuchtigkeit. Nur wenige braune Flecken entstanden, wenn die Blätter 5-10 Stunden nach der Sporenablage feucht blieben (Holmes und Colhoun, 1974) oder bis zu 24 Stunden (Kema et al., 1996). Die Wirt-Parasit-Beziehungen sind bei resistenten und anfälligen Weizenarten gleich. Die Sporenkeimung auf der Blattoberfläche ist unabhängig von der Anfälligkeit die gleiche. Die Zahl der erfolgreichen Penetrationen ist ungefähr gleich, aber das Hyphenwachstum ist bei den anfälligen Sorten schneller, was zu mehr Läsionen führt. Die Hyphen reichen bei allen Sorten über den nekrotischen Bereich hinaus. 44 Sitzung 2 - B.M. Cunfer Ein Toxin könnte bei der Pathogenese eine Rolle spielen (Cohen und Eyal, 1993; Hilu und Bever, 1957). Im Gegensatz dazu war die Kolonisierung bei einer resistenten Linie stark reduziert (Kema et al., 1996).
Stagonospora (Septoria) und Septoria-Pathogene des Getreides: Der Infektionsprozess
B.M. Cunfer, Abteilung für Pflanzenpathologie, Universität von Georgia, Griffin, GA
Der Infektionsprozess wurde am intensivsten für Stagonospora (Septoria) nodorum und Septoria tritici. Eine eingehende Studie über Septoria passerinii verfügbar ist. Fast alle Informationen beziehen sich auf die Infektion durch Pyknidiensporen. Der Infektionsprozess für andere Sporenformen ist jedoch recht ähnlich. Die vorliegenden Informationen beziehen sich meist auf die Infektion von Blättern unter optimalen Bedingungen. Einige Studien wurden an intakten Sämlingspflanzen durchgeführt, andere an abgetrennten Blättern. Infektion der Weizenkoleoptile und des Keimlings durch S. nodorum wurde von Baker (1971) ausführlich beschrieben und von Cunfer (1983) überprüft. Obwohl keine genauen Vergleiche angestellt wurden, hat es den Anschein, dass der Infektionsprozess in jedem Wirt-Parasit-System viele Ähnlichkeiten aufweist und typisch für viele nekrotrophe Krankheitserreger ist. Informationen über Faktoren, die die Entwicklung von Symptomen und die Ausprägung der Krankheit beeinflussen, sind nicht vorhanden, wurden aber von anderen Autoren geprüft (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Eine Zusammenfassung der Faktoren, die die Langlebigkeit der Sporen auf der Blattoberfläche beeinflussen, ist enthalten.
Die Rolle des Cirrus und das Überleben der Sporen auf der Blattoberfläche Die detailliertesten Informationen über die Funktion der Zirren, die die aus dem Pyknidium ausgeschiedenen Pyknidiensporen umhüllen, sind für S. nodorum. Der Zirrus ist ein Gel, das aus Protein- und Saccharidverbindungen besteht. Seine Zusammensetzung und Funktion sind ähnlich wie bei anderen Pilzen der Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths und Peverett, 1980). Die Hauptaufgaben der Zirruskomponenten sind der Schutz der Pyknidiensporen vor Austrocknung und die Verhinderung einer vorzeitigen Keimung.
Die Zirruslösung schützt die Pyknidiosporen, so dass einige von ihnen mindestens 28 Tage lang lebensfähig bleiben (Fournet, 1969). Wurde der Cirrus mit Wasser verdünnt, keimten bei einer Konzentration der Cirruslösung von >20% weniger als 10% der Pyknidiensporen. Bei einer niedrigeren Konzentration liefern die Komponenten Nährstoffe, die die Keimung der Sporen und die Verlängerung der Keimschläuche stimulieren. Die Länge der Keimschläuche nahm bis zu einer Zirruskonzentration von 15% zu und ging dann bei höheren Konzentrationen mäßig zurück (Harrower, 1976). Brennan et al. (1986) berichteten über eine stärkere Keimung in verdünnter Zirrusflüssigkeit. Zirruskomponenten verringerten die Keimung bei einer relativen Luftfeuchtigkeit von 10-60%. Sobald die Sporen verteilt sind, ist die stimulierende Wirkung der Zirrusflüssigkeit wahrscheinlich vernachlässigbar (Griffiths und Peverett, 1980).
Bei einer relativen Luftfeuchtigkeit von 35-45% werden Sporen von S. tritici in Zirren mindestens 60 Tage lang lebensfähig blieben (Gough und Lee, 1985). Die Bestandteile der Zirren können die Keimung der Sporen hemmen, oder das hohe osmotische Potential der Zirren kann die Keimung verhindern. Pyknidiensporen von S. nodorum überlebten keine 24 Stunden bei einer relativen Luftfeuchtigkeit von über 80% bei 20 C. Sporen überlebten zwei Wochen oder länger bei einer relativen Luftfeuchtigkeit von <10% (Griffiths und Peverett, 1980). Wenn die Zirrusflüssigkeit von S. nodorum mit Wasser verdünnt wurde, verloren etwa zwei Drittel der Pyknidiensporen innerhalb von 8 Stunden ihre Lebensfähigkeit, und nach 30 Stunden bei Tageslicht keimten nur 5%. Wurden die Sporen im Dunkeln gelagert, blieben 40% nach 30 Stunden lebensfähig (Brennan et al., 1986).
Trockene Konidien von S. nodorumim Schatten und bei direkter Sonneneinstrahlung mindestens 56 Stunden im Freien überlebten (Fernandes und Hendrix, 1986a). Die Keimung der Pyknidiensporen von S. nodorum wurde durch kontinuierliche UV-B-Bestrahlung (280-320 nm) gehemmt, während die Keimung von S. tritici war es nicht. Die Ausdehnung der Keimschläuche unter kontinuierlicher UV-B-Strahlung wurde bei beiden Pilzen im Vergleich zur Dunkelheit gehemmt (Rasanayagam et al., 1995).
Infektion durch Septoria nodorum
Der Prozess des Eindringens in den Wirt und die Entwicklung von S. nodorum innerhalb des Blattes wurde von mehreren Forschern eingehend untersucht (Baker und Smith, 1978, Bird und Ride 1981, Karjalainen und Lounatmaa, 1986; Keon und Hargreaves, 1984; Straley, 1979; Weber, 1922). Die Pyknidiensporen setzen sich in der Regel in den Vertiefungen zwischen zwei Epidermiszellen fest, und viele Versuche, in das Blatt einzudringen, beginnen dort. Die Sporen keimen auf der Blattoberfläche als Reaktion auf freie Feuchtigkeit (Fernandes und Hendrix, 1986b). Sie beginnen 2-3 Stunden nach der Ablagerung zu keimen, und nach 8 Stunden kann die Keimung 90% erreichen. Das Eindringen in die Blätter beginnt etwa 10 Stunden nach der Ablagerung der Sporen (Bird und Ride, 1981; Brönnimann et al., 1972; Holmes und Colhoun, 1974).
Zu Beginn der Keimung ist der Keimschlauch von einem amorphen Material umgeben, das mit dem Blatt verwachsen ist. Keimschläuche, die aus beiden Enden einer Spore und aus Zwischenzellen wachsen, neigen dazu, entlang der Vertiefungen zwischen den Zellen zu wachsen und sind oft entlang der Längsachse des Blattes ausgerichtet (O'Reilly und Downes, 1986). Die Hyphen von Sporen, die sich nicht in Vertiefungen befinden, wachsen willkürlich und verzweigen sich gelegentlich (Straley, 1979). Es bildet sich ein Appressorium mit einem Infektionszapfen, der die Kutikula und die periklinen Wände der Epidermiszellen direkt in das Zelllumen durchdringt, was zu einem schnellen Zelltod führt.
Viele Penetrationen erfolgen zunächst subkutikulär oder durch seitliches Wachstum der Hyphen innerhalb der Zellwand, bevor sie in das Zytoplasma eindringen (Bird und Ride, 1981; O'Reilly und Downes, 1986). Das Eindringen durch offene und geschlossene Spaltöffnungen erfolgt ebenfalls und kann schneller sein als das direkte Eindringen (Harrower, 1976; Jenkins, 1978; O'Reilly und Downes, 1986; Straley, 1979). Die Keimschläuche verzweigen sich an den Spaltöffnungen und den Verbindungsstellen der Epidermiszellen. Das Eindringen eines Keimschlauches in eine Spaltöffnung kann ohne Bildung eines Appressoriums erfolgen. Manchmal erfolgt das Eindringen durch Trichome (Straley, 1979). Offenbar scheitern die meisten Penetrationsversuche, wobei sich in den Zellen an der Stelle des versuchten Eindringens dichte Papillen bilden (Karjalainen und Lounatmaa, 1986; Bird und Ride, 1981).
Nach dem Eindringen sterben die Epidermiszellen schnell ab und verholzen, und die Hyphen wachsen in das Mesophyll. Die Mesophyllzellen verformen sich, und verholztes Material lagert sich außerhalb einiger Zellen ab, die dann kollabieren. Die Verholzung erfolgt, bevor die Hyphen die Zelle erreichen. Der Prozess ist bei allen Sorten gleich, entwickelt sich aber bei resistenten Sorten langsamer. Die Hyphen wachsen interzellulär zwischen den Epidermiszellen und dann in das Mesophyll. Wenn das Mesophyll durchdrungen ist, beginnt die Zerstörung der Chloroplasten innerhalb von 6-9 Tagen (Karjalainen und Lounatmaa, 1986).
Die Photosyntheserate beginnt jedoch innerhalb eines Tages nach der Infektion und vor dem Auftreten von Symptomen zu sinken (Krupinsky et al., 1973). Sklerenchymgewebe um die Gefäßbündel verhindert eine Infektion des Gefäßgewebes. Die Gefäßbündel blockieren die Ausbreitung der Hyphen durch das Mesophyll, es sei denn, das Sklerenchymgewebe ist noch jung und nicht vollständig ausgebildet (Baker und Smith, 1978).
Stagonospora nodorum setzt in vitro und während der Infektion von Weizenblättern ein breites Spektrum an zellwandabbauenden Enzymen frei, darunter Amylase, Pektinmethylesterase, Polygalacturonasen, Xylanasen und Cellulase (Baker, 1969; Lehtinen, 1993; Magro, 1984). Die Informationen über den Zellwandabbau durch Enzyme stimmen mit histologischen Beobachtungen überein, wobei diese Enzyme in Verbindung mit Toxinen wirken können. Die Enzymempfindlichkeit kann mit der Resistenz und der Geschwindigkeit der Pilzbesiedlung zusammenhängen (Magro, 1984). Wie viele Nekrotrophen produzieren auch Septoria- und Stagonospora-Pilze in vitro phytotoxische Verbindungen. Das Absterben und der Tod von Zellen vor dem Wachstum der Hyphen in das Mesophyllgewebe (Bird und Ride, 1981) steht im Einklang mit der Toxinproduktion. Eine endgültige Rolle der Toxine im Infektionsprozess und ihr Zusammenhang mit der Wirtsresistenz ist jedoch noch nicht nachgewiesen (Bethenod et al., 1982; Bousquet et al., 1980; Essad und Bousquet, 1981; King et al., 1983). Unterschiede im Wirtsspektrum zwischen weizen- und gerstenadaptierten Stämmen von S. nodorum kann mit der Toxinproduktion zusammenhängen (Bousquet und Kollmann, 1998). Der Beginn der Sporenkeimung und der prozentuale Anteil der gekeimten Sporen werden durch die Wirtsanfälligkeit nicht beeinflusst (Bird und Ride, 1981; Morgan 1974; Straley, 1979; Straley und Scharen, 1979; Baker und Smith, 1978).
Bird und Ride (1981) berichteten, dass die Ausdehnung der Keimschläuche auf der Blattoberfläche bei resistenten Sorten langsamer war als bei anfälligen Sorten. Dieser Mechanismus, der mindestens 48 Stunden nach der Sporenablage zum Tragen kommt, deutet auf eine Resistenz gegen die Ausdehnung der Keimschläuche vor dem Eindringen hin. Bei resistenten Sorten gab es weniger erfolgreiche Penetrationen, und die Penetration verlief bei resistenten Sorten langsamer (Baker und Smith, 1978; Bird und Ride, 1981). Es wurde angenommen, dass die Lignifizierung die Infektion sowohl bei resistenten als auch bei anfälligen Sorten einschränkt, aber andere Faktoren verlangsamten die Entwicklung des Pilzes bei resistenten Linien. Bei anfälligen Linien können schneller wachsende Hyphen der Lignifizierung der Wirtszellen entgehen. Vier Tage nach der Inokulation von Gerste mit einem Weizenbiotyp-Isolat von S. nodorumDie Hyphen wuchsen durch die Kutikula und manchmal durch die äußeren Zelluloseschichten der epidermalen Zellwände. Unter den Penetrationshyphen lagerten sich dicke Papillen ab, und die Zellen wurden nicht durchdrungen (Keon und Hargreaves, 1984).
Infektion durch Septoria passerinii
Green und Dickson (1957) legen eine detaillierte Beschreibung des Infektionsprozesses von S. passerinii auf Gerste. Der Infektionsprozess ist ähnlich wie bei S. tritici. Wie bei S. tritici ist die Zeitspanne, die für die Durchdringung der Blätter erforderlich ist, wesentlich länger als bei S. nodorum. Die Keimschläuche verzweigen sich und wachsen wahllos über die Blattoberfläche, manchmal aber auch entlang der Vertiefungen zwischen den Epidermiszellen. Das Eindringen in das Blatt erfolgt fast ausschließlich über die Spaltöffnungen. Die Keimhyphen schwellen an, und wenn die Penetration erfolglos bleibt, dehnen sich die Hyphen weiter aus. 48 Stunden nach der Sporenablage findet keine Penetration mehr statt. Nach 72 Stunden verdicken sich die Keimschläuche über den Spaltöffnungen, wachsen zwischen den Wächterzellen und auf den Stirnflächen der Nebenzellen sowie in die substomatären Hohlräume. Ein direktes Eindringen zwischen Epidermiszellen ist nur selten zu beobachten.
Die Sporenkeimung und das Eindringen in den Wirt sind bei resistenten und anfälligen Sorten gleich. Die Ausdehnung der Hyphen innerhalb der Blätter ist bei resistenten Sorten viel geringer, und Papillen werden auf vielen, aber nicht auf allen Zellwänden beobachtet. Die Hyphen wachsen unter der Epidermis von einem Stoma zum anderen, dringen aber nicht zwischen die Epidermiszellen ein. Das Mesophyll ist kolonisiert, aber es bilden sich keine Haustorien. Nachdem die Mesophyllzellen nekrotisch geworden sind, kollabieren die Epidermiszellen. Die Myzelentwicklung im Blatt ist spärlich und wird in der Regel durch Gefäßbündel blockiert. In jüngeren Blättern, wenn die Gefäßscheide weniger entwickelt ist, dringen die Hyphen zwischen dem Bündel und der Epidermis hindurch. Pyknidien bilden sich in substomatalen Hohlräumen, meist auf der Blattoberseite (Green und Dickson, 1957).
Faktoren, die die Langlebigkeit von Sporen beeinflussen auf der Blattoberfläche Von den Stagonospora- und Septoria-Erregern des Getreides sind endgültige Informationen über den Infektionsprozess nur für folgende Arten bekannt S. nodorum, S. triticiund S. passerinii. Wie viele andere nekrotrophe Krankheitserreger lösen auch diese beiden Erregergruppen keine Überempfindlichkeitsreaktion aus. Ein wesentlicher Unterschied im Infektionsprozess zwischen Septoria- und Stagonospora-Erregern besteht darin, dass die Sporenauskeimung und -penetration bei S. nodorum als für S. tritici und S. passerinii. Dies hat einen erheblichen Einfluss auf die Epidemiologie von Krankheiten.
Die Septoria-Erreger dringen hauptsächlich über die Spaltöffnungen in die Pflanze ein, während S. nodorum dringt sowohl direkt als auch über die Spaltöffnungen ein. S. nodorum dringt in die Epidermiszellen ein und tötet sie schnell ab, aber S. tritici und S. passerinii töten die Epidermiszellen erst ab, wenn sich die Hyphen durch das Blattmesophyll verzweigt haben und eine schnelle Nekrose einsetzt. Histologische Untersuchungen des Pilzwachstums nach dem Eindringen in den Wirt stimmen mit den Daten überein, die aus epidemiologischen Studien zur Wirtsresistenz gewonnen wurden. Die Resistenz verlangsamt die Geschwindigkeit der Wirtskolonisierung, hat aber keine nennenswerten Auswirkungen auf die Entwicklung der Läsionen.
Die Mechanismen, die die Reaktion des Wirts steuern, ob sie nun mit Enzymen und Toxinen oder anderen Stoffwechselprodukten zusammenhängen, die von den Erregern während der Infektion freigesetzt werden, sind noch unklar. Es gibt nur wenige Informationen über die Infektion durch Ascosporen. Der Infektionsprozess ist wahrscheinlich dem der Pyknidiensporen sehr ähnlich. Die Ascosporen von Phaeosphaeria nodorum keimen in einem breiten Temperaturbereich, und ihre Keimschläuche dringen direkt in das Blatt ein. Nach Rapilly et al. (1973) keimen die Ascosporen jedoch im Gegensatz zu den Pyknidiensporen nicht in freiem Wasser.
Septoria spp. Infektionsmodell
Septoria-Infektionen sind bei niedrigen Temperaturen möglich, wobei Temperaturen unter 7°C nicht unbedingt innerhalb von 2 Tagen zu einer Infektion führen. Das Temperaturoptimum der Krankheit wird im Bereich von 16 bis 21°C erreicht. Infektionen sind bei einer hohen relativen Luftfeuchtigkeit oder Blattnässe von 14 Stunden oder länger möglich. Um den Bedingungen gerecht zu werden, haben wir uns entschlossen, in Modelle für schwache, mittlere und schwere Infektionen zu unterteilen. Schwache Infektionen können gegeben sein, wenn es dem Erreger möglich ist, das Wirtsgewebe zu infizieren. Das bedeutet, dass eine schwache Infektion stattfinden kann, wenn die Temperaturen im Minimalbereich liegen und die Blattnässeperioden von kritischer Dauer sind. Eine mäßige Infektion findet unter Bedingungen statt, bei denen die meisten Infektionsversuche zu vernünftigen Ergebnissen führen, und eine schwere Infektion findet unter Bedingungen statt, bei denen der Erreger optimale Infektionsbedingungen vorfindet.
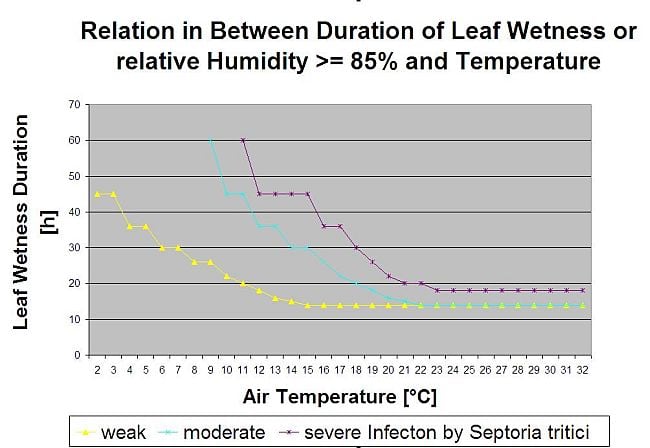

Die Infektion beginnt nach einem Regen von 0,5 mm. Wir haben beschlossen, kein Modell für die Pyknidienbildung zu verwenden. Als Voraussetzung für die Pyknidienbildung wird ein Zeitraum mit einer relativen Luftfeuchtigkeit von mehr als 85% angenommen. Die Lebensdauer der Pyknidien wird auf 24 Stunden geschätzt. In allen Klimazonen, in denen Septoria tritici eine Chance hat, sich zu infizieren, werden wir 2 Stunden finden, die diese Bedingungen an fast jedem Tag um den Sonnenaufgang herum erfüllen.
Bewertung des Schweregrads der Infektion
Um den Infektionsdruck durch Septoria tritici zwischen dem Stadium 10 (erstes Blatt durch die Koleoptile) und dem Stadium 32 (zweiter Knoten mindestens 2 cm über dem ersten Knoten) sowie zwischen 32 und 51 (Beginn des Blattaustriebs) beurteilen zu können, müssen wir den Schweregrad der Infektionen anhand der klimatischen Bedingungen bewerten. Diese Bewertung erfolgt auf einer Skala von 1 bis 5. Ein Schweregrad von 1 wird vergeben, wenn die Bedingungen für eine schwache Infektion erfüllt sind und es weniger als 5 mm geregnet hat, andernfalls ist der entsprechende Schweregrad 2. Ein Schweregrad von 3 wird vergeben, wenn eine mäßige Infektion erfüllt ist und es weniger als 5 mm geregnet hat. Wenn es bei einer mäßigen Infektion mehr als 5 mm oder bei einer schweren Infektion weniger als 5 mm geregnet hat, wird ein Schweregrad von 4 vergeben.
Eine schwere Infektion mit mehr als 5 mm Regen entspricht einem Schweregrad von 5.
Septoria tritici Bewertung des Krankheitsdrucks
Das Klima ist nur ein Faktor, der über den Krankheitsdruck im Feld entscheidet. Die beiden anderen Faktoren sind die Geschichte des Feldes und die Anfälligkeit der angebauten Sorte. Wenn wir die Krankheitsschwerewerte von Stadium 10 bis Stadium 32 auf einen Wert von 4 akkumulieren können, können wir einen schwachen Krankheitsdruck durch das Klima erwarten. Erreicht dieser Wert den Wert 6, ist mit einem mäßigen Krankheitsdruck zu rechnen, und wenn er den Wert 10 erreicht, ist mit einem hohen Krankheitsdruck durch das Klima zu rechnen. Wenn wir die Anfälligkeit der Sorte und die Vorgeschichte des Feldes kennen, können wir in dieser Situation bei schwachem oder mittlerem Krankheitsdruck spritzen oder nicht. Ein akkumulierter Wert von 10 kann in jedem Fall zu einer Spritzung im Stadium 32 führen. Die Entscheidung für eine Spritzung in einem späteren Stadium hängt eher vom Frühjahrsklima ab. Wenn es uns gelingt, die Schweregrade seit Stadium 10 auf einen Wert von 6 zu akkumulieren, können wir mit einem schwachen Krankheitsdruck rechnen. Erreicht dieser Wert 10, so ist mit einem mittleren Krankheitsdruck zu rechnen und erreicht dieser Wert 15, so ist aufgrund der klimatischen Situation mit einem hohen Krankheitsdruck zu rechnen.
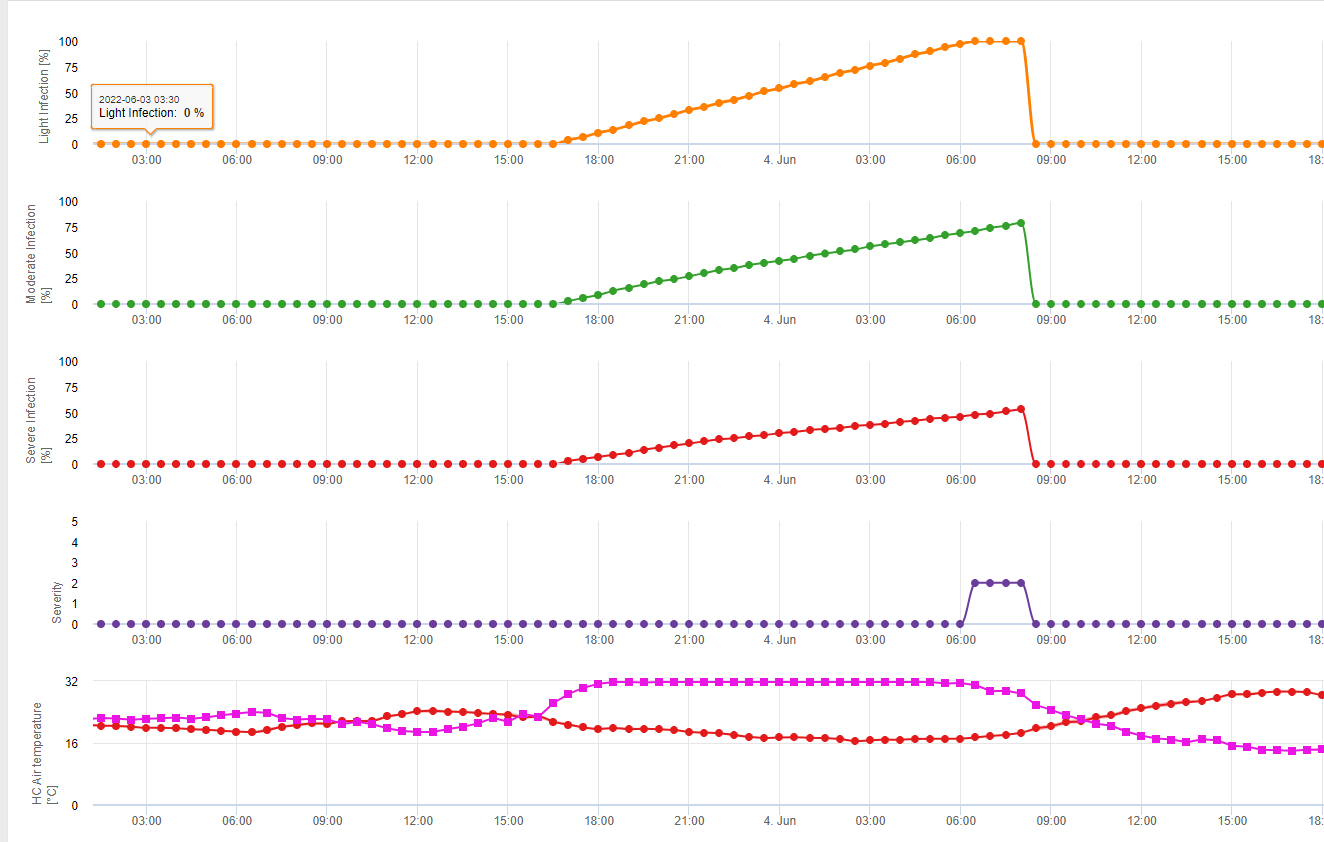
In FieldClimate zeigen wir die Septoria tritici Schweregrad zusammen mit den drei verschiedenen Infektionsschweregraden in einer Grafik (siehe oben). Aufgrund der Niederschläge und der langen Blattnässeperioden sind die Bedingungen für eine schwere Infektion durch S. tritici wurden am 14. und 16. Mai voll erfasst. Die Schweregrade erreichen am 14. Mai den höchsten Wert von 5, was bedeutet, dass jetzt ein hohes Infektionsrisiko besteht.
Die Infektionsbiologie von Stagonospora nodorum unterscheidet sich in gewissem Maße von der von S. tritici, aber dieser Unterschied ist nicht groß genug für ein eigenes Modell. Daher schlagen wir vor, dieses Modell für den gesamten Komplex der Stagnospora- und Septoria-Krankheiten bei Getreide zu verwenden, einschließlich S. passerinii. S. tritici und S. passerinii benötigen in der Regel längere Blattnässeperioden als S. nodorum. In Gebieten mit einem hohen Druck von S. nodorum Infektionen, die als schwach eingestuft werden und einen Schweregrad von 2 aufweisen, sollten ernster behandelt werden als in anderen Bereichen.
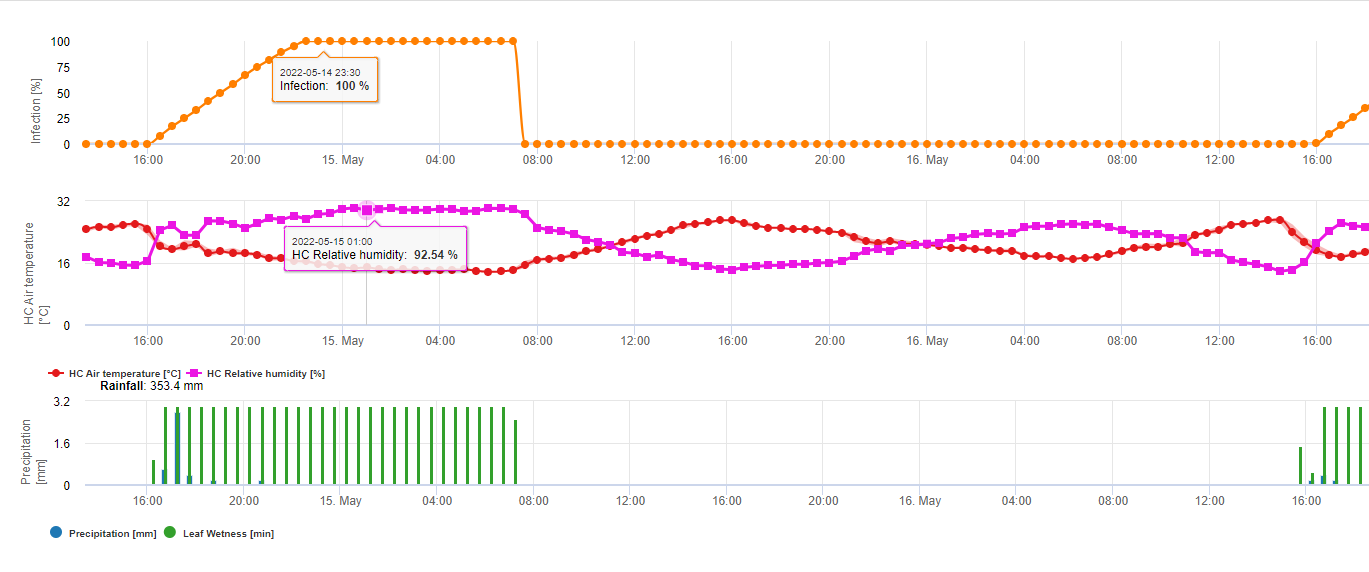
Für Septoria nodorum ein Risikomodell ist in FieldClimate dargestellt (siehe oben). Ein hohes Risiko wurde am 17. Juni und 7. Juli (100%) festgestellt. Je nach dem Stadium der anfälligen Pflanzen für eine Infektion müssen Pflanzenschutzmaßnahmen in Betracht gezogen werden, wenn das Risiko 80% erreicht (siehe auch Wettervorhersage, Pflanzenschutz). Liegt das Risiko bei 100% und wurde bereits eine Infektion festgestellt, müssen systemische Pflanzenschutzmaßnahmen (kurative Anwendung) ergriffen werden, um die Pflanze zu schützen.