Sojarost, insbesondere der Asiatische Sojarost, ist eine schwere Krankheit der Sojabohne, die durch den Pilzerreger Phakopsora pachyrhizi. Verbreitung der Krankheit: Asien und Australien, in jüngerer Zeit auch in Afrika, Nord- und Südamerika. Die Krankheit wurde erstmals 1902 in Japan beobachtet. Der Erreger verbreitete sich über Asien und Australien bis nach Amerika. Der Sojarost kann in Gebieten mit Frosttemperaturen nicht überwintern, aber er kann sich durch Wind schnell über so große Entfernungen ausbreiten, sich so explosionsartig entwickeln und einen so schnellen Blattverlust verursachen, dass er heute zu den am meisten gefürchteten Krankheiten in den Sojabohnenanbaugebieten der Welt gehört.
Symptome und Anzeichen
Die ersten Symptome von Sojarost, verursacht durch Phakopsora pachyrhizi beginnen als sehr kleine braune oder ziegelrote Flecken auf den Blättern. Im Freiland beginnen diese Flecken in der Regel im unteren Kronendach bei oder nach der Blüte, obwohl auch Sämlinge unter bestimmten Umständen infiziert werden können. Häufig treten die ersten Läsionen an der Basis der Blattfieder in der Nähe des Blattstiels und der Blattadern auf. Dieser Teil der Blattfieder hält den Tau wahrscheinlich länger zurück, was die Bedingungen für eine Infektion begünstigt. Die Läsionen bleiben klein (2-5 mm im Durchmesser), nehmen aber mit dem Fortschreiten der Krankheit zu. In diesen Läsionen bilden sich Pusteln, so genannte Uredinia, meist auf der unteren Blattoberfläche, die viele Urediniosporen produzieren können.
Die erhabenen Pusteln sind mit bloßem Auge zu erkennen, insbesondere wenn sie sporulieren. Auch wenn die Läsionen klein sind, weist jede Läsion oft mehrere Pusteln (Uredinien) auf. Wenn die Pusteln aktiv sind, können die Läsionen vollständig mit Urediniospora bedeckt sein. Die Uredinosporen des Sojarosts sind blass gelbbraun bis farblos und weisen eine stachelige (kurzstachelige) Oberflächenverzierung auf. Diese Färbung unterscheidet sich von vielen anderen Rostpathogenen, deren Sporen oft rötlich-braun (rostfarben) sind. Die Keimung von P. pachyrhizi Die Bildung von Urediniosporen erfolgt durch eine äquatoriale (zentrale) Pore, die einen Keimschlauch bildet, der in einem Appressorium endet, mit dem der Pilz direkt oder über ein Stoma in den Wirt eindringt.
Je mehr Läsionen sich auf einem Blatt bilden, desto mehr vergilbt der befallene Bereich, und schließlich fällt das Blatt von der Pflanze ab. Der Sojarost beginnt in der Regel in der unteren Baumkrone, breitet sich aber schnell an der Pflanze aus, bis alle Blätter befallen sind. Schwer erkrankte Pflanzen können vollständig entlaubt werden. Der Verlust an effektivem Blattgewebe führt zu Ertragsminderungen durch weniger und kleinere Samen. Es wurden Ertragseinbußen von 30 bis 80% berichtet, aber die Höhe der Verluste hängt davon ab, wann die Krankheit beginnt und wie schnell sie fortschreitet. Neben Blättern kann Sojarost auch auf Blattstielen, Stängeln und sogar Keimblättern auftreten, die meisten Rostläsionen treten jedoch auf Blättern auf.
Die Läsionen können entweder hellbraun oder rotbraun sein. Bräunliche Läsionen weisen viele Pusteln auf, die zahlreiche Urediniospora produzieren. Rotbraune Läsionen, die als mäßige Resistenzreaktion gelten, haben nur einige wenige Pusteln, die nur wenige Urediniosporen produzieren. Wie im Abschnitt Krankheitsmanagement erörtert wird, hängt dieser Läsionstyp vom Erregerstamm ab und kann auf demselben Blatt wie lohfarbene Läsionen auftreten, oder lohfarbene Läsionen können sich mit zunehmendem Alter rotbraun verfärben. Die Symptome und Anzeichen auf anderen Wirten, wie z. B. Kudzu, sind ähnlich, obwohl die Größe der Läsionen unterschiedlich sein kann. Wenn die Pusteln altern, können sie schwarz werden. Dies wird durch die Bildung einer Schicht von Teliosporen in den Pusteln verursacht, die die Pusteln von Uredinien in Telien verwandeln.
Teliosporen haben zwei Funktionen: das Überleben des Pilzes in Abwesenheit eines lebenden Wirtes (Overseasoning) und die sexuelle Vermehrung. Die dicken Wände der Teliosporen schützen den Pilz vor der Umwelt und dem Angriff anderer Organismen. Bei Rostpilzen keimen die Teliosporen aus und bilden ein Basidium und vier Basidiosporen, bei denen eine sexuelle Rekombination stattfindet. Die Keimung von P. pachyrhizi Die Verbreitung von Teliosporen wurde nur im Labor beobachtet und scheint nicht wesentlich zum Fortbestehen der Krankheit auf dem Feld beizutragen.
Biologie der Krankheitserreger
Es gibt zwei eng verwandte Pilze, die Rost an Sojabohnen verursachen: Phakopsora pachyrhizider manchmal als asiatischer oder australasiatischer Sojarost-Erreger bezeichnet wird, der aber inzwischen auch in der westlichen Hemisphäre vorkommt, und P. meibomiaeden sogenannten Neuwelt-Sojarost, der nur in der westlichen Hemisphäre vorkommt. Abgesehen von einigen wenigen Merkmalen scheinen die beiden Pilze morphologisch identisch zu sein, aber P. pachyrhizi ist bei Sojabohnen viel aggressiver als P. meibomiae. Bis heute, P. meibomiae hat in Mittel- und Südamerika nachweislich keine nennenswerten Ertragseinbußen verursacht. Die beiden Arten können anhand von DNA-Analyseprotokollen unterschieden werden.
Wie andere Roste sind auch die Sojarost-Erreger obligate Parasiten, die einen lebenden Wirt benötigen, um zu wachsen und sich zu vermehren. Unter natürlichen Bedingungen können sie außerhalb des Wirts nur wenige Tage lang als Urediniospora überleben. Soweit wir wissen, produzieren beide Sojarost-Erreger nur zwei Arten von Sporen: Urediosporen und Teliosporen (Abbildung 15). Dies steht im Gegensatz zu anderen Rostarten, die bis zu fünf Sporenstadien haben können (z. B. Weizenstammrost). Beim Sojarost ist, wie bei den meisten Rostarten, das Uredinialstadium das sich wiederholende Stadium. Das bedeutet, dass die Urediniosporen denselben Wirt, auf dem sie entstanden sind (Sojabohne), noch in derselben Saison infizieren können. Epidemien können sich schnell aus nur wenigen Pusteln entwickeln, da die sporenproduzierenden Pusteln bereits 7 bis 10 Tage nach der Infektion entstehen und jede Pustel Hunderte von Urediosporen produzieren kann. Teliosporen werden in alten Läsionen gebildet, scheinen aber in der Natur nicht zu keimen, und es sind weder ein alternativer Wirt noch Aecien oder Spermogonien bekannt.
Ohne die Keimung der Teliosporen kann keine sexuelle Vermehrung stattfinden. Das Fehlen der sexuellen Vermehrung sollte die Variabilität des Rostpilzes begrenzen, aber dennoch gibt es eine erhebliche Variabilität in P. pachyrhizi in Bezug auf die Virulenz. Dies hat die Verwendung einzelner Gene für die Resistenz bei Sojabohnen eingeschränkt, da in kurzer Zeit neue Isolate des Erregers entstehen, die das Resistenzgen überwinden. Es ist nicht bekannt, wie diese Variabilität zustande kommt P. pachyrhizi. Weizen-Streifenrost, Puccinia striiformishat einen ähnlichen Lebenszyklus wie P. pachyrhizi ohne funktionsfähiges Telialstadium und daher ohne sexuelle Vermehrung, aber mit vielen Rassen. Es könnte sein, dass jedes Resistenzgen so spezifisch ist, dass eine einzige Mutation im richtigen Gen des Pilzes es ihm ermöglicht, auf Wirten mit dem neuen Resistenzgen virulent zu werden.
Epidemiologie
Sojarost-Epidemien beginnen mit dem Eintreffen von Inokulum (Uredinosporen) aus der Luft. Dieser Erreger ist unter den Rostarten einzigartig, weil er viele alternative Wirte hat, die als Inokulumquellen dienen können. Alternativwirte sind andere Pflanzen, die mit demselben Erreger infiziert werden können, aber nicht erforderlich sind, um den Lebenszyklus des Erregers zu vollenden. Ausweichwirte sind nicht zu verwechseln mit Alternativwirten, bei denen es sich um eine andere Pflanze als den Hauptwirt handelt, die ein Erreger für die Vollendung seines Lebenszyklus benötigt. In frostfreien Gebieten wie Südamerika, Mittelamerika, dem karibischen Becken, Südtexas und Florida könnte die Inokulumquelle in der Nähe von Sojabohnen-Durchwuchspflanzen, Kudzu oder einem anderen Alternativwirt liegen. In Gebieten, in denen Frost herrscht, wie im Mittleren Westen der USA, muss das Inokulum von Überwinterungsquellen, die Hunderte von Kilometern entfernt sein können, eingeblasen werden.
Die Wiedereinschleppung von obligaten Krankheitserregern in eine weit entfernte Region kommt bei verschiedenen anderen Krankheiten vor, z. B. bei Weizenstängelrost und Falschem Mehltau, z. B. Blauschimmel bei Tabak. Da Sporen von P. pachyrhizi empfindlich auf ultraviolette Strahlung reagieren, werden diese Rostsporen wahrscheinlich durch Gewitter, bei denen Wolken die Sporen vor der Sonne schützen, über weite Strecken verbreitet. Sobald lebensfähige Sporen auf der Blattoberfläche eines geeigneten Wirts gelandet sind, hängen die Infektion und die anschließende Entwicklung der Epidemie von den Umweltbedingungen ab. Im Allgemeinen erfolgt die Infektion, wenn die Blätter feucht sind und die Temperatur zwischen 8°C und 28°C liegt, mit einem Optimum von 16°C bis 28°C. Bei 25 °C erfolgt die Infektion teilweise schon nach 6 Stunden Blattnässe, optimal sind jedoch 12 Stunden. Nach der Infektion können innerhalb von 7 oder 8 Tagen Läsionen und Pusteln mit Urediniospora auftreten, und der nächste Infektionszyklus beginnt.
Dieser kurze Lebenszyklus bedeutet, dass Sojarost-Epidemien unter den richtigen Bedingungen schnell von fast nicht nachweisbaren Werten auf sehr hohe Werte ansteigen können. Sojarost-Epidemien können innerhalb eines Monats von einem nicht nachweisbaren Niveau bis zur Entlaubung fortschreiten. Die Epidemien können scheinbar noch schneller voranschreiten, da frühe Infektionen im unteren Kronendach auftreten und schwer zu finden sind. Neben der Umwelt beeinflusst auch das Alter der Pflanzen die Sojarostepidemien. In der Regel werden Rostläsionen an Sojabohnen erst zur Blütezeit festgestellt, es sei denn, es gibt zu Beginn der Saison hohe Inokulumwerte. Dies kann darauf zurückzuführen sein, dass die Pflanzen anfälliger für Rost sind, wenn der Wirt in die Reproduktionsphase eintritt, oder darauf, dass die Sporen in den unteren Teilen der Baumkronen besser vor UV-Strahlung geschützt sind, oder darauf, dass die Bedingungen in den Baumkronen feuchter werden, wenn sich die Baumkronen schließen. In jedem Fall können sich in jedem Wachstumsstadium Läsionen bilden, aber eine größere Zunahme der Krankheit tritt erst nach der Blüte auf.
Zwei Modelle für Sojarost werden in FieldClimate beschrieben und berechnet:
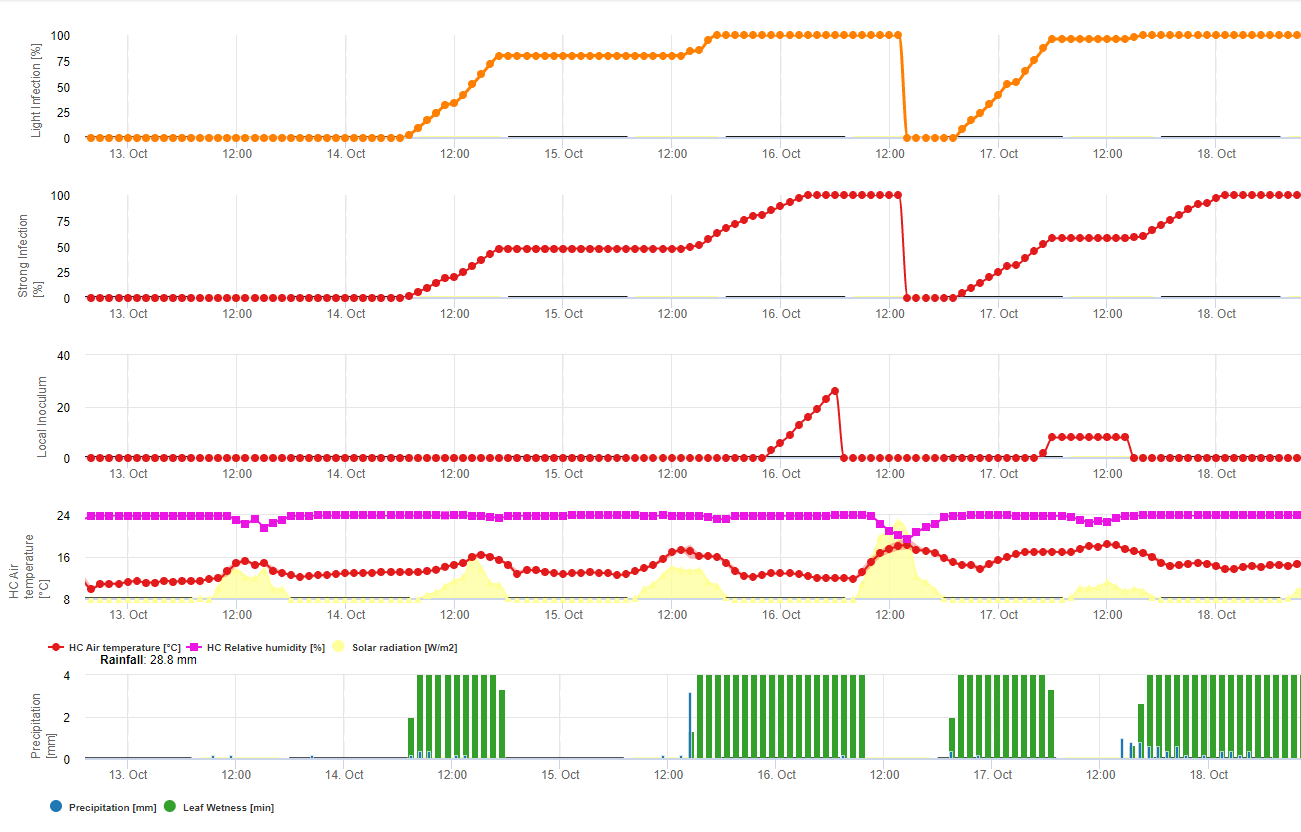
Modell 1: Hauptfaktoren für die Berechnung der Infektion und des Schweregrads des Sojarosts sind die Blattnässe und die Temperatur (Achtung: Die Infektion muss 100% betragen, bevor die Schweregrade (1-3) berechnet werden können). Die Blätter müssen nass sein und die Temperatur muss zwischen 8 und 28°C liegen - dann beginnt die Berechnung der Infektion (die Summe der 7200 °Minuten ergibt 100%). Wenn die Infektion 100% beträgt, werden in Abhängigkeit von der Temperatur und der Dauer der Blattnässe die Schweregrade von 1 bis 3 errechnet.
Modell 2: Hier werden viel mehr Faktoren in die Berechnung von Sojarost einbezogen: Temperatur, Blattnässe, relative Luftfeuchtigkeit, Niederschlag und Sonneneinstrahlung (die Pilzhyphen können tagsüber in die Spaltöffnungen eindringen).
Wenn die Infektion mit Sojarost 100% erreicht, wird der Schweregrad berechnet. Wenn beides (Infektion und Schweregrad) 100% beträgt, wird das Auftreten (die Etablierung?) eines lokalen Inokulums vorhergesagt.
Literatur:
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), S. 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L., und Royer, M. H. 1989. Auswirkungen von Dauer, Häufigkeit und Temperatur von Blattnässeperioden auf Sojarost. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte, und A.P. Dias. Das Risiko von Sojarost durch einen Vergleich mit Brasilien. Abteilung für Pflanzenpathologie, Iowa State University
- Yang, X. B. 1995. Assessment and management of the risk of soybean rust. Proceeding of the soybean rust workshop, 9-11 August 1995. J. B. Sinclair und G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL
Sclerotinia-Stängelfäule oder Weißschimmel bei Sojabohnen ist eine Krankheit, die durch den Pilz Sclerotinia sclerotiorum. Die Krankheit kann zu erheblichen Ertragseinbußen oder sogar zur völligen Vernichtung der Ernte führen, wenn Sojabohnen in befallenen Böden gepflanzt werden und ein dichtes Pflanzendach mit längeren feuchten Wetterperioden vorhanden ist. Ertragseinbußen treten in der Regel auf, wenn das Auftreten der Krankheit 15 % oder mehr beträgt. In den meisten Jahren in North Dakota stellt die Sklerotinia-Stängelfäule nur ein geringes bis mittleres Problem für die Landwirte dar, und die Krankheit wird in Dürrejahren nur selten beobachtet. Nasses Wetter ist ein wichtiger Faktor für die Entwicklung der Krankheit. Bei bewässerten Sojabohnen ist sie ein anhaltendes Problem. Neben einer Verringerung des Saatgutertrags führt die Krankheit auch zu einer verminderten Saatgutqualität und zu Saatgut, das mit den schwarzen Sklerotien des Pilzes kontaminiert ist. Die Kontamination des Saatguts kann ein ernsthaftes Problem für exportiertes Saatgut darstellen, da sie zur Zurückweisung der Saatgutpartie an ausländischen Einfuhrhäfen führen kann. Außerdem können Sklerotien, die in den Boden gelangen, andere Kulturen in der Fruchtfolge beeinträchtigen. Die Sklerotinia-Stammfäule kann von den Landwirten durch Kenntnis des Erregers und des Krankheitszyklus in den Griff bekommen werden.
Symptome
Die Symptome werden in der Regel erst beobachtet, wenn sich das Blätterdach zwischen den Reihen geschlossen hat und ein feuchtes Mikroklima entstanden ist. Das Welken und Verwelken der Blätter und das anschließende Absterben der Pflanzen sind in der Regel die ersten beobachteten Symptome. Bei einer genauen Inspektion unter dem Pflanzendach ist ein watteartiges, weißes Myzelwachstum (Pilzfäden) an Stängeln, Blättern oder Schoten zu erkennen. Die Läsionen entwickeln sich an Hauptstängeln und Seitenzweigen. Schließlich umhüllen die Läsionen die Stängel und die darüber liegenden Pflanzenteile sterben ab. Die Stängel erscheinen gebleicht und manchmal durch die fortgeschrittene Fäulnis zerfetzt. Aus dem weißen Myzel, das auf dem Pflanzengewebe wächst, bilden sich große, schwarze Sklerotien in verschiedenen Formen und Größen. Sklerotien bilden sich auch im Mark des Stängels und haben eine charakteristische zylindrische Form. Die Samen in befallenen Schoten sind in der Regel geschrumpft und können vom Pilz befallen oder durch schwarze Sklerotien ersetzt werden. Das Saatgut ist in der Regel mit Sklerotien kontaminiert, wenn infizierte Pflanzen geerntet werden.
Biologie der Krankheitserreger
Der Pilz hat ein umfangreiches Wirtsspektrum von über 370 Pflanzenarten und verursacht Krankheiten an einer Vielzahl von Kulturen wie Sonnenblumen, Trockenbohnen, Raps, Kartoffeln, Luzerne, Buchweizen, Lupinen, Senf, Topinambur, Saflor, Linsen, Flachs, Felderbsen und vielen Gemüsearten. In North Dakota ist es selten, dass dieser Erreger bei einigen dieser Kulturen wie Flachs und Kartoffeln ernsthafte Schäden verursacht. Es gibt auch viele verbreitete Wirtspflanzen für breitblättrige Unkräuter, wie z. B. Sumpfholunder, Lammfuchsschwanz, Schweinegras, Kanadadistel und Ackersenf. Der Pilz, der den Weißschimmel an Sojabohnen verursacht, ist derselbe, der auch den Weißschimmel oder die Sclerotinia-Krankheit an Sonnenblumen, Trockenbohnen, Raps und anderen Kulturen verursacht. Sclerotinia sclerotiorum überwintert hauptsächlich als Sklerotien im Boden. Die Sklerotien keimen aus und bilden kleine hellbraune bis braune Pilze, die Apothecien genannt werden (etwa ein Achtel bis ein Viertel Zoll im Durchmesser). Diese produzieren Sporen, die als Ascosporen bezeichnet werden und die Krankheit auf Sojabohnen und anderen anfälligen Pflanzen auslösen.
Krankheitszyklus
Feuchtigkeit und Blüte sind entscheidende Faktoren für die Krankheitsentwicklung. Krankheiten treten in der Regel nicht vor dem Schließen des Blätterdachs auf, da ein dichtes Blätterdach kühle Temperaturen und ein feuchtes Mikroklima um die Stängel herum begünstigt und die Bodenfeuchtigkeit nach Regen oder Bewässerung hoch hält. Der Ausbruch der Krankheit ist auch eng mit der Blüte verbunden. Nach sieben bis 14 Tagen hoher Bodenfeuchtigkeit keimen die Sklerotien in den oberen Zentimetern des Bodens und bilden die pilzartigen Apothecien. Ein einziges Sklerotium kann mehrere Apothecien hervorbringen. Die Apothecien schleudern ihre Ascosporen mit Gewalt in die Luft, wo sie von Luftströmungen zu den Sojapflanzen getragen werden. Die wichtigste Quelle für Ascosporen sind Apothecien, die innerhalb des Feldes produziert werden, aber Ascosporen können auch von benachbarten oder nahe gelegenen Feldern eingeblasen werden. Eine Apothecie kann über mehrere Tage hinweg enorme Mengen an Ascosporen produzieren. Die Ascosporen überleben für kurze Zeit auf dem Pflanzengewebe, überwintern aber nicht. Ascosporen benötigen einen Wasserfilm und eine Nahrungsgrundlage wie abgestorbenes oder alterndes Blütengewebe, um zu keimen und zu wachsen, bevor sie die Pflanze infizieren. Blütengewebe ist die wichtigste Nahrungsgrundlage für die Auslösung von Infektionen. Häufig beginnen die Infektionen in den Achseln der Stängel, wo das alternde Blütengewebe abgefallen ist und sich festgesetzt hat. Infektionen können auch durch Wunden auftreten, die durch Hagel oder andere Verletzungen entstanden sind. Ein Wasserfilm auf der Pflanzenoberfläche fördert die Entwicklung von Läsionen und vergrößert das Ausmaß der Gewebeschäden. Die anfängliche Krankheitsentwicklung erfordert in der Regel mehr als 40 Stunden ununterbrochene Nässe auf der Pflanzenoberfläche, aber wenn die Krankheit erst einmal ausgebrochen ist, ermöglichen auch kürzere Nässeperioden die Entwicklung von Läsionen. Dies ist der Grund, warum die Krankheit mit längeren Perioden bewölkten, feuchten und regnerischen Wetters in Verbindung gebracht wird. Wenn die Pflanzenoberflächen abtrocknen, wird das Fortschreiten der Krankheit verlangsamt. Kühle Temperaturen zwischen 59 und 75 Grad Celsius begünstigen die Entwicklung der Krankheit. Je dichter das Pflanzendach ist, desto günstiger sind die Umweltbedingungen für die Krankheit. Sojabohnen mit fester Aussaat und gelagerte Pflanzen sind daher anfälliger für die Entwicklung von Krankheiten. Sklerotien werden gebildet, wenn das Myzel in und auf dem Pflanzengewebe wächst. Diese Sklerotien keimen nicht aus, um während der Saison weitere Apothecien zu bilden, sondern werden bei der Ernte und Bodenbearbeitung in den Boden zurückgebracht und überwintern als Inokulum (Quelle des infektiösen Pilzes) für eine künftige anfällige Kultur. Sklerotien sind sehr widerstandsfähige Strukturen und überleben lange Zeit im Boden.
Quelle: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
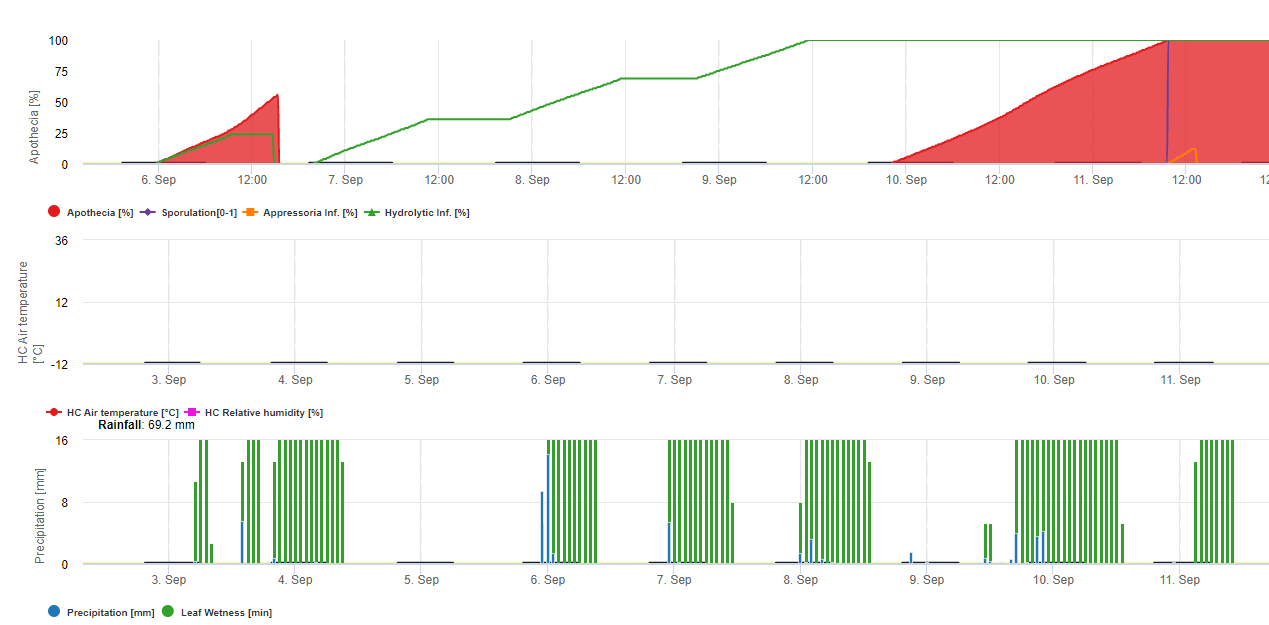
In FieldClimate wird das Modell für Sclerotinia in Abhängigkeit von einer Regenperiode, der relativen Luftfeuchtigkeit und Temperatur sowie der Blattnässe berechnet. Bei langen Regenperioden wird eine Infektion durch die Bildung eines Appressoriums durch den Pilzerreger empfohlen. Eine andere Art der Infektion wird als "hydrolytische Infektion" bezeichnet. Sie beruht auf der Freisetzung von hydrolytischen Enzymen, die nacheinander die Kutikula, die Mittellamellen sowie die primären und sekundären Zellwände der Pflanze abbauen und die gesamte Pflanze zerstören. Modell im Detail beschrieben auf dieser Seite.