Biologie der Plasmopara viticola (Echter Mehltau)
P. viticola ist ein obligater Parasit. Das bedeutet, dass er grüne, frische Rebenorgane zum Wachstum benötigt. Während der vegetationslosen Zeit überdauert er und bildet Fruchtkörper, so genannte Oosporen. Oosporen von Oomyceten können sehr lange im Boden überleben. Daher können wir den Falschen Mehltau an Orten finden, an denen eine Infektion nicht in jedem Jahr möglich ist. Im Frühjahr, wenn der Oberboden feucht und warm genug ist, bilden die Oosporen so genannte Makrosporangien, die bis zu 200 Zoosporen ins freie Wasser abgeben können. Die Zoosporen werden mit dem Wind in Wassertröpfchen zu den Blättern und Büscheln transportiert. Sie haben zwei Geißeln und bewegen sich in einem Wasserfilm auf der Unterseite der Blätter oder der Trauben und jungen Beeren, um ein Stoma zu finden und in das Pflanzengewebe einzudringen. Sie dringen in das Stoma ein und keimen dort aus, wobei sie innerhalb von weniger als einer Stunde ihr gesamtes Plasma übertragen. In mikroskopischen Untersuchungen waren das Auffinden des Stomas, die Zystierung und die Keimung im Stoma innerhalb von 90 Minuten abgeschlossen.
P. viticola wächst im Interzellularraum und ernährt sich von Haustorien, die die Epidermis- und Parenchymzellen durchdringen. In Abhängigkeit von Temperatur und relativer Luftfeuchtigkeit entwickelt er genügend interzelluläres Wachstum mit genügend Haustorien, um einen Substomatelkörper zu bilden, der den gesamten Substomatelbereich ausfüllt und das Epidermisgewebe vom Parenchymgewebe abhebt. Dies führt zu dem sichtbaren Symptom des Ölflecks.
Oomyceten sind Sporenbildung in Abwesenheit von Licht, wenn die relative Luftfeuchtigkeit ist sehr hoch. Unter P. viticola Bei Temperaturen unter 12 °C und einer relativen Luftfeuchtigkeit unter 95% findet keine Sporenbildung statt. Die Sporangiaphoren werden von den Substomatellbläschen gebildet und treten aus dem Stoma aus. Die frisch gebildeten Sporangien sind klebrig und können nur mit Wasser aus den Sporangiaphoren entfernt werden. Wenn die relative Luftfeuchtigkeit abnimmt, werden die Sporangien klebrig und können auch durch Wind entfernt werden.
Die Sporangien geben bis zu 20 Zoosporen in freies Wasser ab. Diese Zoosporen müssen durch den Wind in Wassertröpfchen verteilt werden, um auf frische Blätter zu gelangen, oder die Sporangien können durch Regen oder Wind selbst verteilt werden. Der Infektionsprozess der Primär- und Sekundärinfektion ist derselbe.
Aufgrund der großen Bedeutung des Sexualstadiums für die Überwinterung des Erregers kann man davon ausgehen, dass in allen Weinbergen, in denen der Falsche Mehltau auftritt, passende Paarungsformen vorkommen. Die Zoosporenbildung findet auf älteren Blättern im Sommer und Frühherbst statt.
Primäre Infektion im Detail
Infektionen, die von den Oosporen ausgehen, werden als Primärinfektionen bezeichnet. Dieser Begriff ist irreführend, da im Frühsommer mehrere Primärinfektionen auftreten können. Im epidemiologischen Geschehen spielt die Primärinfektion keine große Rolle, wenn genügend Ölflecken in den Weinbergen vorhanden sind und das Infektionspotenzial der Sommersporen (Sporangien) die Oosporen übersteigt.
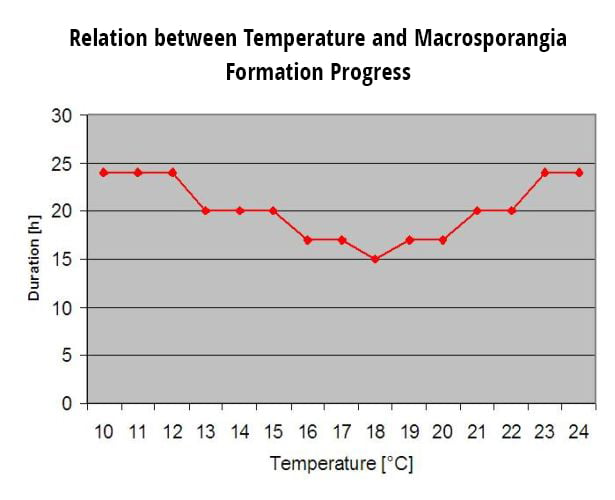
Die überwinternden Oosporen entwickeln sich zu sogenannten Makrosporangien, wenn etwa 24 Stunden lang eine ausreichende relative Luftfeuchtigkeit herrscht. Die Makrosporangien entlassen ihre Zoosporen in freies Wasser und starken Regen (wie bei einem Gewitter). Die Sporen werden zu den Blättern und/oder grünen Trieben der Rebe getragen. Primärinfektionen also, brauchen länger regnerisch Perioden oder mehrere aufeinander folgende Gewitter. Der erste Regen führt dazu, dass die abgefallenen Blätter mit Wasser gesättigt werden, und starker Regen an den folgenden Tagen bewirkt, dass die Makrosporangien ihre Zoosporen freisetzen, die die Weinblätter oder -triebe erreichen. Für eine erfolgreiche Infektion ist eine ausreichend lange Periode der Blattnässe erforderlich, damit die Zoosporen die Stomata der Blätter oder Triebe erreichen und diese infizieren können (die Abbildung oben zeigt den Entwicklungszyklus des Falschen Mehltaus der Rebe (Plasmopara viticola).
Das Modell für die Primärinfektion mit Falschem Mehltau prüft zunächst, ob das Wetter für die Entwicklung von Makrosporangien geeignet ist. Dies ist der Fall, solange die Blätter feucht sind oder die relative Luftfeuchtigkeit nach dem Regen nicht unter 70% fällt. Je nach Temperatur können sich die Makrosporangien innerhalb von 16 bis 24 Stunden entwickeln. Wenn reife Makrosporangien vorhanden sind, wird dies in der Anzeige der Primärinfektion des Falschen Mehltaus grafisch dargestellt. Wenn Makrosporangien vorhanden sind, kann ein starker Regenfall ihre Zoosporen verbreiten. Ein Dauerregen von 5 mm wird als Starkregen interpretiert und die Zoosporen werden verbreitet, eine Primärinfektion ist dann möglich, wenn die Blattnässe lange genug angedauert hat.
 Sekundäre Infektionen
Sekundäre Infektionen
 Sekundäre Infektionen
Sekundäre InfektionenSekundärinfektionen von Plasmopara viticola sind nur möglich, wenn in Ihrem Weinberg oder in dessen Nähe bereits reife Ölflecken vorhanden sind. Ölflecken sind reif, wenn sie in der Lage sind, zu sporulieren (Sporangien zu bilden). Sporangien werden nur gebildet von Nacht. Sonnenlicht hemmt die Sporenbildung. Sporangien werden produziert, wenn es wärmer als 12°C und dem relative Luftfeuchtigkeit ist sehr hoch. Die Sporangienproduktion nimmt mit der Temperatur bis zu 24 °C zu. Die optimale Temperatur für die Sporenbildung bei europäischen Rebsorten (Vitis vinifera) liegt bei etwa 24°C. Wenn die Temperaturen 29°C überschreiten, kann keine Sporenbildung stattfinden. In unserem Modell prüfen wir, ob Feuchtigkeit von mehr als 95% während der Nacht auftritt. Wenn diese Bedingung für eine akkumulierte stündliche Temperatur von mehr als 50°C andauert, ist die Sporulation beendet und neue Sporangien von Plasmopara im Weinberg existieren. Zum Beispiel entsprechen 50°C Stunden 4 Stunden mit 13°C oder 3 Stunden mit 17°C.
Sporangien von Plasmopara viticola haben eine begrenzte Lebensdauer. Je wärmer es ist und je trockener die Luft, desto schneller sterben sie ab (in fieldclimate.com setzen wir auf 0 zurück, wenn die relative Luftfeuchtigkeit unter 50% liegt). Sie sterben definitiv in der nächsten Tau- oder Blattnässeperiode ab, die für eine Infektion zu kurz ist. Die Fitness älterer Sporangien ist daher recht begrenzt. Unser Modell geht davon aus, dass die Lebensdauer der Sporangien auf einen Tag begrenzt ist.
Um neue Infektionen zu verursachen, müssen die Sporangien verbreitet werden. Für die Verbreitung der Sporen gibt es zwei Möglichkeiten: Wenn es unmittelbar nach der Bildung der Sporangien regnet, verbreiten sie sich mit den Regenspritzern. Bleiben die Weinblätter lange genug feucht, entsteht eine hohe Anzahl von Plasmopara viticola Infektionen stattfindet. Wenn der nächste Morgen ohne Regen und mit abnehmender Luftfeuchtigkeit beginnt, lösen sich die vertrockneten Sporangien von den Zweigen. Schon eine leichte Luftbewegung trägt sie auf gesunde Blätter. Wenn es nicht bald regnet, werden sie absterben.
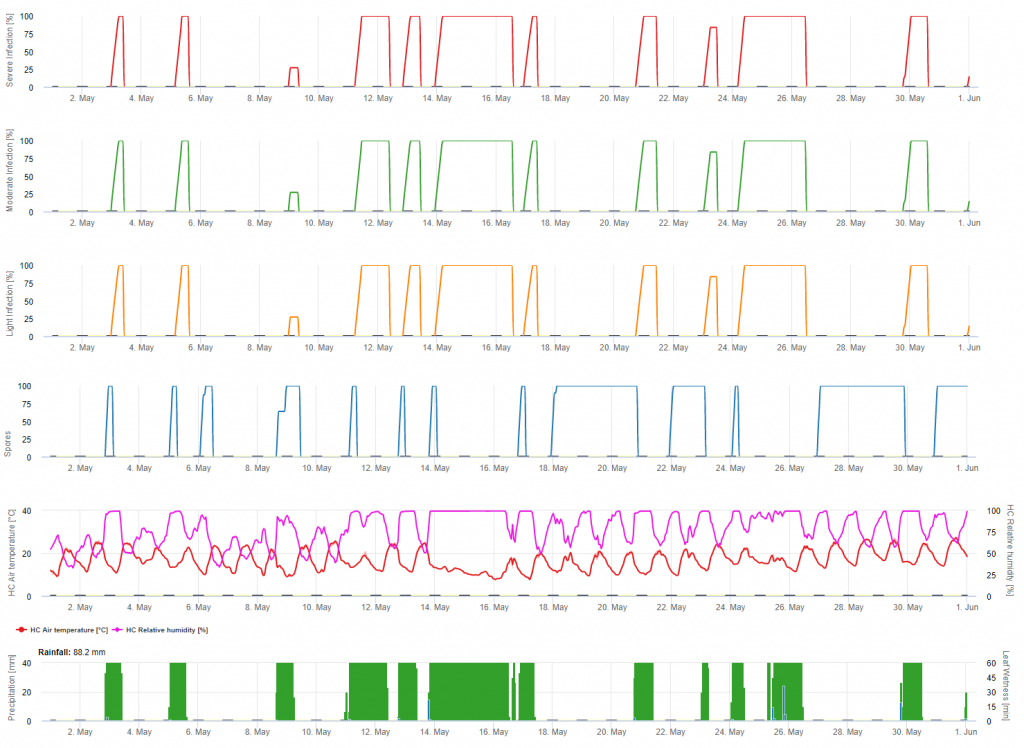
Falscher Mehltau Primärinfektion
Die Infektionen werden nach der Makrosporenentwicklung (orange, fünftes Diagramm) in drei verschiedenen Schweregraden dargestellt (schwach: orange, mäßig: grün, stark: rot), die von der Regenmenge und damit der Verteilung der Sporen im Weinberg abhängen. Wenn eine Infektion mit 100% festgestellt wird, sind optimale Bedingungen für das Eindringen in das Pflanzengewebe gegeben (Pflanzenschutzmittel) und die Inkubationskurve (erstes Diagramm) beginnt anzusteigen. Wenn die Inkubationszeit beendet ist (100%) und kein Pflanzenschutzmittel eingesetzt wurde, sollten die Symptome (Ölflecken) auf dem Feld kontrolliert werden.
1. Die Kurve des Infektionsverlaufs und nach vollständiger Infektion die sich daraus ergebende Inkubationszeit (Zeit zwischen Infektion und sichtbaren Symptomen auf dem Feld).
2. Eine Infektionsverlaufskurve wird ermittelt, wenn Makrosporangien oder Sporangien vorhanden sind und die Blätter feucht sind. Sie steigt mit zunehmender Blattnässe und Temperatur an. Erreicht die Infektionsverlaufskurve den Wert 100%, kann davon ausgegangen werden, dass die Infektion abgeschlossen ist. Die zur jeweiligen Infektion gehörende Inkubationsverlaufskurve wird mit dem Beginn einer Infektion berechnet. Wenn eine Infektion nicht abgeschlossen werden kann, wird die Inkubationskurve ebenfalls gestoppt.
3. Die Wetterbedingungen werden auf der gleichen Seite wie die Ergebnisse des Krankheitsmodells angezeigt. So können Sie mit einem Blick Temperatur, relative Luftfeuchtigkeit, Regen und Blattnässe während der Infektion überprüfen.
4. Das Modell zeigt die Infektionsdaten für die Primär- und Sekundärinfektionen mit Falschem Mehltau an Weinreben auf. Dies ist die wichtigste Information. Infektionen, die noch nicht durch präventive oder kurative Fungizide abgedeckt sind, können katastrophale Auswirkungen auf den Ertrag/ die Qualität haben. Weitere Informationen über die Schwere der Infektion (schwach/mittel/schwer) könnten hilfreich sein, um eine Pflanzenschutzstrategie in Abhängigkeit von der Geschichte des Weinbergs (Menge des Inokulums), der Sorte und der Erfahrung der letzten Jahre festzulegen.
Pflanzenschutzstrategien:
- Prophylaktische und systemische oder kurative Fungizide werden in großem Umfang zur Bekämpfung von Falschem Mehltau eingesetzt. Prophylaktische Chemikalien werden vor, aber so nah wie möglich an einem Infektionsereignis in der Zeit der größten Wirtsanfälligkeit, zwischen einer Trieblänge von 10 cm und erbsengroßen Beeren, eingesetzt.
- Prophylaktische Chemikalien können auch eingesetzt werden, indem so kurz wie möglich vor einem vorhergesagten Wetterereignis mit mehr als 2 mm Regen gesprüht wird (für den Primärkreis).
- Ein kuratives Fungizid sollte so schnell wie möglich nach einem Infektionsereignis (100%-Infektion) und vor dem Auftreten von Ölflecken angewendet werden. Achten Sie darauf, die Entwicklung einer Fungizidresistenz zu verhindern.
- Die Überwachung des Weinbergs auf das Auftreten von Falschem Mehltau (Ölflecken) sollte regelmäßig und unter Berücksichtigung von mindestens 200 Rebstöcken durchgeführt werden. Ein Risiko wird als gegeben angesehen, wenn mehr als zwei Ölflecken pro 50 Rebstöcke festgestellt werden.
Das Modell basiert auf den Veröffentlichungen von MUELLER-THURGAU, ARENS, MUELLER und SLEUMER, BLAESER, HERZOG, GEHMANN und vielen anderen Arbeitern, die seit dem frühen neunzehnten Jahrhundert mit dem Forschungsnetzwerk zur Epidemiologie des Falschen und Echten Mehltaus an Weinreben verbunden sind.
Biologie der Uncinula necator
Der Echte Mehltau der Weinrebe wird verursacht durch den Pilz Uncinula necator. Er ist ein obligater Parasit, das heißt, er braucht grünes Wirtsgewebe, um sich zu ernähren. U. necator Jahrhundert nach Europa und verbreitete sich bis zum Ende dieses Jahrhunderts in allen Weinbauländern. Er hat sehr starke Auswirkungen auf den Ertrag und kann zum Absterben der Reben führen, wenn er nicht chemisch bekämpft wird. Alle europäischen Rebsorten sind für diesen Krankheitserreger anfällig. Die Züchtung von Resistenzen gegen diesen Krankheitserreger ist recht neu, und die Ergebnisse werden noch mehrere Jahre Arbeit erfordern.
U. necator muss die vegetationslose Zeit in den meisten Weinbaugebieten überwinden. Dazu nutzt der Erreger zwei Mechanismen: a) er überlebt in seinen Fruchtkörpern, den so genannten Cleistothecien, oder b) er überlebt den Winter auch eingekapselt in schlafenden Knospen, wo er den ganzen Winter über lebendes Gewebe findet. Ob in einem Weinberg nur eine oder beide Überwinterungsformen vorkommen, hängt von der Krankheitsgeschichte dieses Weinbergs ab. Knospen können nur infiziert werden, solange sie nicht geschlossen und von einem Ring aus seneszentem Gewebe umgeben sind. Dies ist nur für einige Wochen nach der Bildung der Fall. Im normalen Weinbau werden die Reben auf Äste mit weniger als 14 Knospen oder auf Ausläufer zurückgeschnitten. Diese verbleibenden Knospen können nur bis zur Blüte infiziert werden. Auf der anderen Seite finden wir Fahnentriebe vom Stadium BBCH 15 bis 19. Fahnentriebe sind teilweise oder vollständig mit Mehltau bedeckte Triebe, die sich aus latent infizierten Knospen bilden. Diese Fahnentriebe sind eine frühe und sehr potente Inokulumquelle. Sie werden meist nicht innerhalb eines Weinbergs verteilt. Dies führt zu Stellen mit frühem und starkem Krankheitsbefall im Weinberg und zu Stellen mit geringem oder mäßigem Krankheitsbefall zu Beginn der Saison. Die Fahnentriebe repräsentieren aufgrund ihrer vegetativen Geschichte nur einen Paarungstyp. Dies kann zu Weinbergen führen, in denen wir aufgrund der Dominanz eines einzigen Paarungstyps nur wenige oder gar keine Cleistothecien finden.
Wenn Cleistothecien das einzige Inokulum sind, werden wir eine kontinuierliche Überwinterung von Cleistothecien feststellen, die nur in Jahren, in denen sich die Krankheit sehr früh entwickelt, in Fahnenschösslinge übergehen kann. Die Bildung von Cleistothecien ist aufgrund der Gelbfärbung des sexuellen Mischgewebes gut sichtbar. Sie kann in ungespritzten Weinbergen ab dem Hochsommer gefunden werden. In gespritzten Weinbergen dauert es länger, bis sich die Krankheit ausbreitet und die Paarungstypen aufeinander treffen können. Die Ausbreitung von Uncinula necator im Sommer wird vor allem durch die Temperatur beeinflusst.
Die optimalen Temperaturen für den Echten Mehltau liegen zwischen 21°C und 32°C. Je mehr Stunden täglich in diesem Temperaturbereich liegen, desto höher ist das Risiko für Echten Mehltau. Die Konidienbildung, die Infektion und die Geschwindigkeit der Inkubation benötigen ähnliche Temperaturbereiche. Aufgrund der Tatsache, dass U. necator kein freies Wasser für die Infektion und keine hohe relative Luftfeuchtigkeit für die Konidienbildung benötigt, gibt es keine verwertbaren Auswirkungen des Feuchtigkeitsparameters direkt auf den Erreger.
Das Vorhandensein von freier Feuchtigkeit steht in Wechselwirkung mit U. necator durch seinen gemeinsamen Hyperparasiten Ampelomyces quisqualis. Dieser pilzliche Hyperparasit wächst innerhalb der Hyphen des Mehltaupilzes. Er wandelt Konidien und Cleistothecien in eigene Pyknidien um. Er benötigt freie Feuchtigkeit oder eine sehr hohe relative Luftfeuchtigkeit, um zu infizieren und mit dünnen, schnell wachsenden Hyphen auf den Blättern zu wachsen, um Mehltau-Läsionen zu finden. Starker Befall U. necator Die Läsionen verfärben sich in ein sehr dunkles Braun. Während der Regenzeit sind sie von weißem Sporenexsudat bedeckt, das aus den Pyknidien des Hyperparasiten stammt. Ampelomyces quisqualis ist auch als kommerzielles Produkt zur Bekämpfung von Mehltau erhältlich.
Praktische Arbeit
Wir haben verschiedene Modelle für den Infektionsprozess in FieldClimate.
a) der Pilz überwintert hauptsächlich in Form eines Myzels. Im Frühjahr beginnt das Myzel zu wachsen und bildet Konidiophoren mit Konidien (also keine Primärinfektion).
b) Wenn sich der Pilz im Herbst auf Beeren entwickelt, könnte die geschlechtliche Form vorhanden sein (Bildung von Cleistothecien), so dass im Frühjahr die Freisetzung von Ascosporen zu einer Primärinfektion führen könnte (Verwendung: Modell POWDERY MILDEW/ASCOSPORE INFECTION).
Kalifornisches Risikomodell (von Thomas Gubler)
Das Modell der UC Davis zur Risikobewertung des Echten Mehltaus (Gubler-Thomas-Modell) basiert auf epidemiologischen Labor- und Feldstudien über den Echten Mehltau an Weinreben in Kalifornien.
Index der Risikobewertung:
Nach der Freisetzung von Ascosporen und der Keimung (für dieses Modell ASCOSPORE INFECTION verwenden) wird die anschließende Entwicklung und Vermehrung des Echten Mehltaus in erster Linie durch die Umgebungstemperaturen beeinflusst. Das Modell der UC Davis bewertet die Umgebungstemperaturen und schätzt das Risiko der Mehltauentwicklung anhand eines Index von 0-100 Punkten ein.
Um den Index für die Risikobewertung einzuleiten, müssen nach dem Austrieb und nach dem Ausbruch der Krankheit drei aufeinanderfolgende Tage mit mindestens sechs aufeinanderfolgenden Stunden mit Temperaturen zwischen 21 und 30 °C vorhanden sein, damit die Mehltauepidemie beginnt. Während der gesamten Berechnung des Index erhöht sich der Index für jeden Tag mit sechs oder mehr aufeinanderfolgenden Stunden zwischen 21 und 32 °C um 20 Punkte. Das Modell schwankt dann je nach Dauer der Tagestemperaturen zwischen 0 und 100 Indexpunkten. Der Index erhöht sich um 20 Punkte für jeden Tag, an dem die Anforderung von sechs aufeinanderfolgenden Stunden mit Temperaturen zwischen 21 und 32 °C erfüllt wird, und er verliert 10 Punkte für jeden Tag, an dem die Anforderung von sechs Stunden nicht erfüllt wird, oder wenn die Temperatur zu irgendeinem Zeitpunkt während des Tages über 32 °C oder unter 21 °C steigt.
Interpretation des Indexes:
Niedrige Indexwerte von 0-30 bedeuten, dass sich der Erreger nicht vermehrt. Landwirte können in Betracht ziehen, Fungizidanwendungen während längerer Zeiträume mit niedrigen Indexwerten zu verschieben. Ein Index von 40-50 gilt als moderat und würde eine Vermehrungsrate des Mehltaus von etwa 15 Tagen bedeuten. Indexwerte von 60-100 deuten darauf hin, dass sich der Erreger schnell vermehrt (alle 5 Tage) und dass das Risiko einer Krankheitsepidemie groß ist.
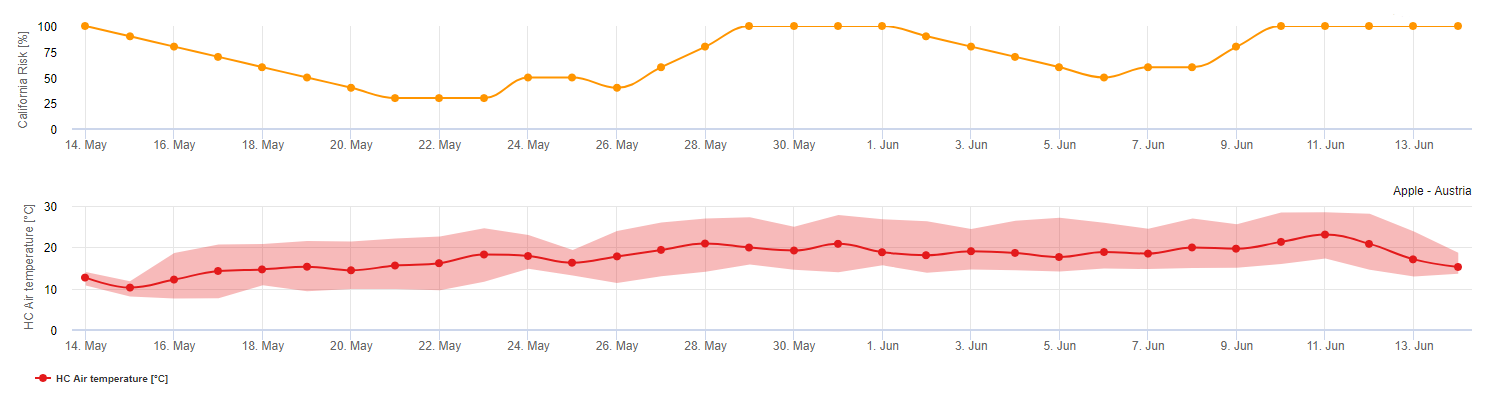
Pessl Instruments-Risikomodell
Die Ausbreitung von Uncinula necator im Sommer wird in erster Linie beeinflusst durch Temperatur. Der Antagonist von U. necator genannt. Ampelomyces quisqualis wird hauptsächlich durch freie Feuchtigkeit begünstigt. Die optimalen Temperaturen für den Echten Mehltau liegen zwischen 21°C und 32°C. Je mehr Stunden täglich die Temperatur in diesem Bereich liegt, desto höher ist das Risiko für Echten Mehltau (siehe Modell Gubler). Tage, an denen die Temperatur mehr als 6 Stunden in diesem Bereich liegt, erhöhen das Risiko um 20 Punkte.. Tage, an denen die Temperatur nicht in diesem optimalen Bereich liegt, verringern das Risiko um 10 Punkte. Dies sind Tage, an denen die Temperatur den ganzen Tag über unter 21 °C liegt oder an denen die Temperatur 6 Stunden lang 32 °C überschreitet. Jedoch Tage mit lange Blattnässe Perioden führen zu einer Etablierung des antagonistischen Pilzes A. quisqualis. Diese werden zu einem Rückgang der U. necator.
Für dieses Modell empfehlen wir, diese Regeln zu befolgen: Bei einem Mehltaustrisiko von weniger als 20 Punkten kann das Spritzintervall verlängert werden. Bei 20 bis 60 Punkten gilt das normale Sprühintervall. Bei einem Risiko von mehr als 60 Punkten sollten Sie das Spritzintervall verkürzen.
Das Risiko beginnt: mit 3 Tagen mit gleich oder mehr als 6 Stunden von: 21°C <= Temperatur 60 Punkte
Das Risiko steigt: Mit jedem Tag mit gleich oder mehr als 6 Stunden von: 21°C <= Temperatur +20 Punkte
Das Risiko sinkt: Mit jedem Tag, an dem die Temperatur nicht 21°C erreicht oder jeden Tag mit 6 Stunden von: 32°C -10 Punkte
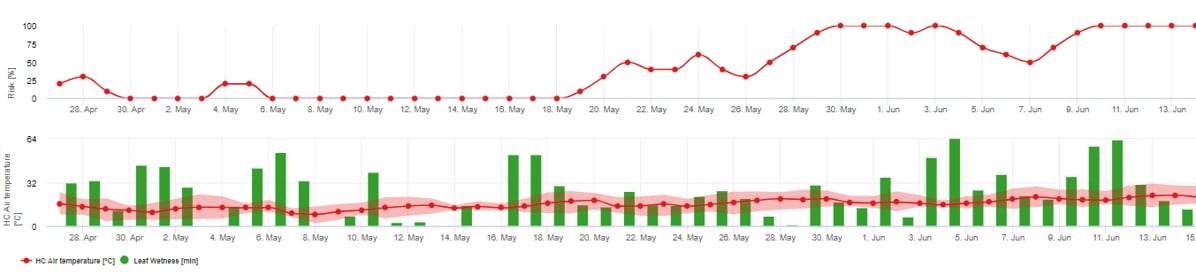
Drittes Risikomodell: Primäres Inokulum von Ascosporen
Wir empfehlen, dieses Modell zu verwenden, wenn die Ascosporen während der kalten Winterzeit gebildet werden (also keine Infektion durch das Myzel ("Fahnensprossen").
Die Ascosporeninfektion prognostiziert die Ascosporenfreisetzung und die Erstinfektionen, die auf den Durchschnittstemperaturen während längerer Blattnässeperioden beruhen.
Für die Berechnung in fieldclimate.com etwa 2,5 mm Niederschlag sind erforderlich, um Ascosporen freizusetzen, gefolgt von at mindestens 8-12 Stunden Blattnässe nach Freisetzung der Ascosporen bei Temperaturen zwischen 10 -15°C für die Infektion. Sobald die Infektion erfolgt ist, geht das Modell in die Phase der Bewertung des Krankheitsrisikos über (Verwendung des Modells Gubler/Risk model) und basiert ausschließlich auf den Auswirkungen der Temperatur auf die Vermehrungsrate des Erregers.
Praktische Anwendung
In diesem Modell zeigen wir die Infektion durch Uncinula necator in einer Linie zwischen 0 und 100%.
Wir zeigen zwei verschiedene Modelle in FieldClimate:
1. Mögliche Ascosporen-Infektionen nach den von David Gadoury und Doug Gubler vorgeschlagenen Modellen werden in der Grafik für Ascosporen-Infektionen dargestellt. Wenn die Verlaufskurve für das Infektionsmodell den Wert 100% erreicht, besteht die Möglichkeit einer Ascosporeninfektion. 3 bis 5 Tage nach diesem Ereignis können erste Symptome des Echten Mehltaus auf Blättern in der Nähe des alten Kordons sichtbar werden.
2. Das Mehltaurisiko nach dem Kalifornien-Modell und dem Modell für gemäßigte Klimazonen ist in der obigen Grafik dargestellt. Perioden mit Risiko müssen mit Fungizidanwendungen abgedeckt werden. In Perioden mit einem Risiko von mehr als 60 Punkten wird empfohlen, häufiger zu spritzen, insbesondere um und kurz nach der Blüte.
Die Schwarzfäule der Trauben betrifft viele Weinbauern in den Vereinigten Staaten und den meisten anderen warmen und trockenen Weinbaugebieten weltweit. Um die Krankheit in den Griff zu bekommen, ist es wichtig, den Lebenszyklus der Krankheit und die Umweltbedingungen zu kennen. Sobald die Infektion erfolgt ist, gibt es verschiedene Methoden zur Bekämpfung der Krankheit.
Biologie
Der Erreger der Traubenschwärze überwintert in vielen Teilen der Weinrebe und kann auch am Boden überwintern, insbesondere in Mumien. Darüber hinaus kann der Erreger "mindestens 2 Jahre lang in Läsionen infizierter Triebe, die als Stöcke oder Ausläufer erhalten bleiben, überwintern".
Wenn es einmal regnet, können die überwinternden Sporen (Ascosporen) freigesetzt werden, die sich in den Mumien auf dem Boden bilden und durch den Wind verbreitet werden können. Einige der Mumien auf dem Boden können einen beträchtlichen Ausstoß von Ascosporen aufweisen, der etwa 2 bis 3 Wochen nach dem Knospenaufbruch beginnt und 1 bis 2 Wochen nach Beginn der Blüte reift. Eine zweite Art von Sporen (Konidien) kann sich auch in Rohrläsionen oder Mumien bilden, die im Spalier verblieben sind, und diese werden durch aufspritzende Regentropfen über kurze Entfernungen (Zentimeter bis Fuß) verbreitet." Die Infektion erfolgt, wenn eine der beiden Sporenarten auf grünem Traubengewebe landet und das Gewebe für eine "ausreichende Zeitspanne, die von der Temperatur abhängt, feucht bleibt." Der Zeitraum, in dem diese überwinternden Sporen eine Infektion verursachen können, hängt von der Quelle ab. Wenn es eine große Infektionsquelle gibt, setzt die Infektion früh ein. Bei Vorhandensein von Feuchtigkeit keimen diese Ascosporen langsam aus, was 36 bis 48 Stunden dauert, dringen aber schließlich in die jungen Blätter und Fruchtstiele (Stiele) ein. Die Infektionen werden nach 8 bis 25 Tagen sichtbar. Bei feuchter Witterung werden Ascosporen produziert und während des gesamten Frühjahrs und Sommers freigesetzt, was zu einer kontinuierlichen Primärinfektion führt. (Wilcox, Wayne F. "Schwarzfäule Guignardia bidwellii." Krankheitsdatenblatt Nr. 102GFSG-D4. 2003. Cornell. 24 Oct. 2010).
Der Schwarzfäulepilz benötigt also warmes Wetter für ein optimales Wachstum; kühles Wetter verlangsamt sein Wachstum. Eine Periode von 2 bis 3 Tagen mit Regen, Nieselregen oder Nebel ist ebenfalls für eine Infektion erforderlich." (Ries, Stephen M. "IPM : Berichte über Pflanzenkrankheiten : Schwarzfäule der Weintraube". Integrierte Schädlingsbekämpfung an der Universität von Illinois. Dez. 1999. 24 Oct. 2010). Regentropfen übertragen diese Sporen, indem sie die Sporen auf verschiedene Pflanzenteile, insbesondere anfällige junge Blätter, bringen. Wenn Wasser vorhanden ist, keimen die Konidien innerhalb von 10 bis 15 Stunden und dringen in junges Gewebe ein. Neue Schwarzfäule-Infektionen setzen sich bis ins späte Frühjahr und in den Sommer hinein fort, wenn es über längere Zeiträume warm und regnerisch ist. Die Konidien sind in der Lage, mehrere Monate nach ihrer Bildung zu keimen und Infektionen zu verursachen. Im August verwandeln sich die Pyknidien in ein Überwinterungsstadium, aus dem wiederum Pseudothecien hervorgehen, in denen die Frühjahrssporen (Ascosporen) gebildet werden. Die Forschung hat gezeigt, dass Ascosporen eine wichtige Quelle für Primärinfektionen im Frühjahr sind. Im Frühjahr, bei feuchtem Wetter, nehmen die Pyknidien auf dem infizierten Gewebe Wasser auf, und die Konidien werden ausgepresst. Die Konidien werden durch Regen wahllos aufgespritzt und können bei Temperaturen zwischen 60 und 90 Grad in weniger als 12 Stunden jedes junge Gewebe infizieren (Ellis, Michael A. "Fact sheet Agricultural and Natural Resources: Grape Black Rot." Abteilung für Pflanzenpathologie. Die Universität von Ohio State Extension. 2008).
Praktische Anwendung
Die Grundlage für unser Modell bildet die von Spotts veröffentlichte Literatur (Spotts. R. A. 1977. Einfluss von Blattnässedauer und Temperatur auf die Infektiosität von Guignardia). In Gesprächen mit Daniel Molitor haben wir das ursprüngliche Spotts-Modell dahingehend geändert, dass es in leichte, mittelschwere und schwere Infektionen unterteilt wird, ähnlich wie bei den Modellen für Apfelschorf. (Daniel Molitor: Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Dissertation Justus-Liebig-Universität Gießen, 2009).
Eine Infektion, die die "Spotts-Kriterien" erfüllt, wird als leichte Infektion eingestuft. Infektionen, die die "Spotts-Kriterien" von 150% erfüllen, werden als mäßig eingestuft, und Infektionen, die die "Spotts-Kriterien" von 200% erfüllen, werden als schwer eingestuft.
Das Modell zeigt die Infektionsdaten für leichte, mittlere und schwere Infektionen. Wenn 100% einer Infektion erfüllt ist, sind die Bedingungen für Sporenausstoß, Keimung und Infektion des Pflanzengewebes gegeben. Der Schweregrad der Infektion hängt von der Zeitspanne der optimalen Temperatur und der Nässeperiode des Pilzes ab. Je nach Infektionsdruck sollten die Infektionen präventiv abgedeckt werden oder es muss kurz nach der Infektion eine kurative Spritzung durchgeführt werden. Im gemäßigten halbtrockenen Klima der Mosel oder der meisten österreichischen Weinbaugebiete wäre eine Spritzung nach Feststellung eines leichten Befalls nicht zu empfehlen.
Lobesia botrana (Totricidae, Olethreutinae) wurde erstmals aus Österreich beschrieben, stammt aber ursprünglich aus Süditalien und ist in ganz Europa, Nord- und Westafrika, dem Nahen Osten und Ostrussland verbreitet. In jüngerer Zeit wurde sie nach Japan eingeschleppt, und 2008 wurde sie erstmals in Chile gemeldet.
Hosts
Traube (Vitis vinifera) und Wolfsmilch-Lorbeer (Daphne gnidium) sind bevorzugte Wirte, aber es wurde auch von Brombeeren (Rubus fruticosus), Stachelbeere (Ribes sp.), schwarze und rote Johannisbeeren (Ribes nigurm), Oliven (Olea europaea), Kirsche (Prunus avium), beschneiden (Prunus domestica), Kakipflaume (Diospyrus kakis), Kiwi (Actinidia chinensis), Granatapfel (Punica granatum), Nelke (Dianthus spp.), und eine Reihe anderer wilder Wirte.
Schaden
Im Mai und Juni nisten sich die Larven der ersten Generation in den Blütenbüscheln ein und ernähren sich von ihnen. Die Larven der zweiten Generation (Juli-August) ernähren sich von grünen Beeren. Die jungen Larven dringen in die Beeren ein, höhlen sie aus und hinterlassen die Schale und die Samen. Die Larven der dritten Generation (August-September) richten den größten Schaden an, indem sie sich in die Beeren und in die Trauben einspinnen und fressen, die dadurch mit Exkrementen verunreinigt werden. Außerdem begünstigt der Fraßschaden an den Beeren die Infektion durch Botrytis und andere sekundäre Pilze wie Aspergillus, Alternaria, Rhizopus, Cladosporium, und Penicillium.
Biologie und Lebenszyklus
Der erwachsene Falter ist etwa 6-8 mm lang und hat eine Flügelspannweite von 11-13 mm. Das Weibchen ist etwas größer. Sowohl die Männchen als auch die Weibchen haben ähnliche mosaikartig gemusterte Flügel. Das erste Flügelpaar (Vorderflügel) ist hellbraun-cremefarben und mit graublauen, braunen und schwarzen Flecken gesprenkelt. Das zweite Flügelpaar ist grau und hat einen fransigen Rand. Die Flügel sind im Ruhezustand glockenförmig über dem Hinterleib gehalten. Die Weibchen legen ihre Eier getrennt ab. Die Eier sind elliptisch und flach, mit einem Durchmesser von etwa 0,6-0,8 mm. Diese linsenförmigen Eier sind sichtbar und schillern cremeweiß, färben sich gelb, wenn sich der Embryo entwickelt, und später schwarz, wenn sich der Kopf der sich entwickelnden Larve bildet. Es gibt fünf Larvenstadien. Das erste Stadium ist etwa 1 mm und das letzte Stadium etwa 12-15 mm groß. Die Larven haben einen schwarzen Kopf und färben sich gelblich-braun, während der Körper die Farbe der Nahrung (Darminhalt) annimmt. Das fünftletzte Larvenstadium spinnt einen gräulichen Silikonkokon, um sich zu verpuppen. Die Puppe ist etwa 4-9 mm lang.
In unserer Klimazone hat der Europäische Traubenwickler zwei bis drei Generationen pro Jahr. Aus wärmeren Regionen Spaniens, Griechenlands und Ägyptens werden vier Generationen berichtet. Das Puppenstadium überwintert in den Silikonkokons (Diapause) unter der Rinde oder in Erdspalten oder an versteckten, geschützten Stellen. Wenn die Lufttemperaturen für etwa 10 bis 12 Tage über 10 °C steigen, schlüpfen die Erwachsenen. Die ersten Männchen werden gefunden und nach einer Woche schlüpfen die Weibchen. Der erste Flug der Männchen kann also bereits mit dem Knospenaufbruch beginnen, und der Flug dauert etwa 4 bis 5 Wochen an. Die Flugaktivitäten finden statt, wenn die Sonne aufgeht und die Temperaturen etwa 12 °C betragen. Während des Fluges paaren sich die Insekten, und die meisten Weibchen paaren sich nur einmal. Die Eiablage beginnt kurz nach der Paarung (etwa ein bis zwei Tage). Sie legen die Eier einzeln auf oder in der Nähe der Blüten ab. Ein Weibchen legt etwa 100 Eier und ist je nach den klimatischen Bedingungen 1 bis 3 Wochen lang erwachsen.
Im Frühjahr dauert der Schlupf der Eier 10 Tage oder länger, während sie im Sommer je nach Temperatur innerhalb von 3 bis 4 Tagen schlüpfen. Die Larven der ersten Generation nisten sich in den Blüten ein und ernähren sich von ihnen, sie können in den Blütenstiel eindringen und die Traube vertrocknen lassen. Die Entwicklung der Larven dauert je nach Witterungsbedingungen etwa 20 bis 30 Tage. Dann verpuppen sie sich unter der Rinde, in der Erde oder auf der Blüte, und die erwachsenen Tiere schlüpfen nach 6 bis 14 Tagen wieder.
Die weiblichen Falter des zweiten und dritten Fluges legen ihre Eier einzeln direkt auf beschatteten Beeren ab. Kurz nachdem die Larve geschlüpft ist, dringt sie in die Beere ein und höhlt sie beim Fressen aus. Eine einzige Traube kann von mehreren Larven befallen sein. Gespinste, Frass und Pilzbefall können zu einer starken Kontamination der Traube führen.
Die untere Entwicklungsschwelle liegt bei etwa 10 °C und die obere Entwicklungsschwelle bei 30 °C. Optimale Entwicklungsbedingungen herrschen bei 26-29 °C und einer Luftfeuchtigkeit von 40 bis 70%. Wenn die Tageslänge abnimmt und es kälter wird, wird das Diapausenstadium eingeleitet. Die Larvenstadien sterben ab, wenn die Temperaturen unter 8 °C sinken, aber die Diapause-Puppe kann sogar die kalten nordeuropäischen Winter überstehen.
Die erste Generation ist kürzer als die Sommergenerationen. Die Eier schlüpfen in etwa 66 Grad Celsius-Tagen (DDC). Larven, die sich von Blütentrauben ernähren, entwickeln sich Berichten zufolge schneller als solche, die sich später in der Saison von Traubenbeeren ernähren, was die Generationszeit beeinflusst. Nicht-diapausierende Puppen benötigen etwa 130 DDC für ihre Entwicklung. Erwachsene Weibchen können etwa 61 Tage nach dem Schlüpfen Eier legen.
Die Schätzungen der DD für eine Generation variieren in der Literatur beträchtlich, von 427 DDC bis 577 DDC in der ersten Generation bis 482 DDC bis 577 DDC in späteren Generationen.
Überwachung und Verwaltung
Die Männchen werden durch Pheromone angelockt, die zur Überwachung des Fluges der Männchen verwendet werden. Stellen Sie vor dem Knospenbruch Fallen mit L. botrana Köder hoch in den Baumkronen (iScout-Pheromone). Stellen Sie mindestens eine Falle pro 30 Hektar oder, falls kleiner, pro Weinbergsblock auf. Wechseln Sie die Köder entsprechend den Empfehlungen des Herstellers aus. Kontrollieren Sie die Fallen wöchentlich, indem Sie die Anzahl der gefangenen Motten aufzeichnen und die gefangenen Motten vom klebrigen Fallenboden entfernen. Zeichnen Sie die wöchentlichen Fänge auf, um den Beginn und den Höhepunkt der männlichen Flüge in jeder Generation zu bestimmen. Setzen Sie die Überwachung mit Fallen bis zum Höhepunkt des dritten Fluges fort.
Der Einsatz von Insektiziden sollte zeitlich auf das Auftauchen der Larven abgestimmt werden, weshalb die Überwachung der Eiablage und die Bestimmung des Schlupfes der Eier für die Bekämpfung dieses Schädlings von entscheidender Bedeutung sind. Bei der ersten Generation sollte die Eiablage vom Höhepunkt bis zum Ende des Fluges überwacht werden. Suchen Sie am Stiel von 100 Trauben nach Eiern, wobei Sie eine Traube pro Rebe auswählen. Notieren Sie das Stadium der meisten gefundenen Eier. Die Eier sind weiß, wenn sie gerade gelegt wurden, und färben sich gelb und später schwarz, wenn die Larven kurz vor dem Schlüpfen stehen. Ein durchscheinendes Eichorion zeigt an, dass die Larve geschlüpft ist. Achten Sie nach dem Schlüpfen der Eier auf Gespinste an Blütenteilen. Öffnen Sie das Gespinst und suchen Sie nach Fraßschäden und Larven.
Beginnen Sie mit der Überwachung von Eiern der zweiten und dritten Generation auf Beeren eine Woche, nachdem die ersten Falter des jeweiligen Fluges in den Fallen gefangen wurden. Setzen Sie die Überwachung auf Eier wöchentlich fort, bis eine Woche nach dem Hauptflug. Untersuchen Sie 100 Trauben, wobei Sie eine pro Rebstock auswählen. Weitere Überwachung der Trauben auf Fraßschäden (Löcher oder hohle Beeren), Gespinste und das Vorhandensein von Larven.
In Ländern, in denen L. botrana etabliert ist, werden die Bekämpfungsmaßnahmen auf die zweite Generation ausgerichtet. Dies ist zum Teil auf das verlängerte Auftreten der ersten Generation und auf den möglichen erneuten Befall durch unbehandelte benachbarte Weinberge zurückzuführen. Eine Behandlung der ersten Generation wird jedoch empfohlen, wenn die Populationen hoch sind oder wenn Behandlungen flächendeckend durchgeführt werden. Insektizide sind nach dem Traubenschluss weniger wirksam. Mehrere Insektizide mit reduziertem Risiko sind zur Bekämpfung von Tortricid-Larven im Weinbau zugelassen. Dazu gehören Insektenwachstumsregulatoren, Spinosyns und Bacillus thuringiensis.
Die Störung der Paarung wird in Europa seit mehreren Jahren untersucht. Sie hat sich als besonders wirksam erwiesen, wenn die Populationen des Traubenwicklers gering sind und wenn sie auf großen Flächen von mehr als 10 Hektar oder flächendeckend angewendet wird.
In der europäischen Literatur wird von zahlreichen Räubern und Parasitoiden berichtet. Zu den Parasitoiden gehören 4 Arten von Tachiniden und fast 100 Arten von Schlupfwespen aus den Familien der Ichneumoniden, Braconiden, Pteromaliden und Chalicididen. Die Parasiten, die den Berichten zufolge die größten Auswirkungen haben, sind diejenigen, die die überwinternden Puppen befallen. In Spanien sind dies die Pteromaliden Dibrachys affinis und D. cavusdie Berichten zufolge eine Puppensterblichkeit von bis zu 70% verursachen, während in Italien die Schlupfwespen Dicaelotus inflexus und Campoplex-Kapitator sind die wichtigsten.
Literatur:
- Briere JF, Pracros P. 1998. Vergleich von temperaturabhängigen Wachstumsmodellen mit der Entwicklung von Lobesia botrana (Lepidoptera: Tortricidae). Environ. Entomol. 27: 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. en las comarcas vitícolas valencianas. Bol. Serv. Plagas 7: 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). In Los parasitos de la vid, estrategias de proteccion razonada. Madrid, Spanien. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33: 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. Studie über die Beziehung zwischen den Fängen von Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) mit Sexualpheromonfallen und der Akkumulation von Gradtagen in Sherry-Weinbergen (Südwest-Spanien). J. Appl. Ent. 125: 9-14.
- Gabel B, Mocko V. 1986. Eine funktionelle Simulation des Europäischen Rebenwicklers Lobesia botrana Den. Et Schiff. (Lep., Torticidae) in der Populationsentwicklung. J. Appl. Ent. 101: 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. Vorhersage der Flugaktivität von Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) im Südwesten Spaniens. J. Appl. Entomol. 133: 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. Control of Sparganothis pilleriana Schiff. and Lobesia botrana (Den. & Schiff). in German vineyards using sex pheromone-mediated mating disruption. Bull. IOBC/WPRS 25: 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera: Tortricidae): influence de l'infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thése N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. Daphne gnidium, eine mögliche einheimische Wirtspflanze des Europäischen Traubenwicklers Lobesia botrana, stimuliert dessen Eiablage. Ist eine Wirtsverlagerung relevant? Chemoecol. 16: 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. Die Anziehungskraft des Traubenwicklers auf Wirts- und Nicht-Wirts-Pflanzenteile im Windkanal: Auswirkungen von Pflanzenphänologie, Geschlecht und Paarungsstatus. Entomol. Exp. Appl. 122: 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Ent. 125: 515-518.
- Moreau J, Benrey B, Thiéry. 2006. Die Rebsorte beeinflusst die Larvenleistung und auch die Reproduktionsleistung der Weibchen des Europäischen Traubenwicklers Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 96: 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. Effects of methoxyfenozide on Lobesia botrana (Den & Schiff) (Lepidoptera: Torticidae) egg, larval and adult stages.
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. In Les ravageurs de la vigne. Féret, Bordeaux. S. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. Die Beziehung zwischen Ausbreitung und Überleben der Larven von Lobesia botrana und ihrer Dichte in den Blütenständen von Weinstöcken. Entomol. Exp. Appl. 84: 109-114.
- Xuéreb A, Thiéry D. 2006. Variiert der natürliche Larvenparasitismus von Lobesia botrana (Lepidoptera: Torticidae) zwischen den Jahren, der Generation, der Dichte des Wirts und dem Rebkultivar? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). In Lepidotteri dei fruttiferi e della vite. Mailand, Italien. Pp. 85-88.
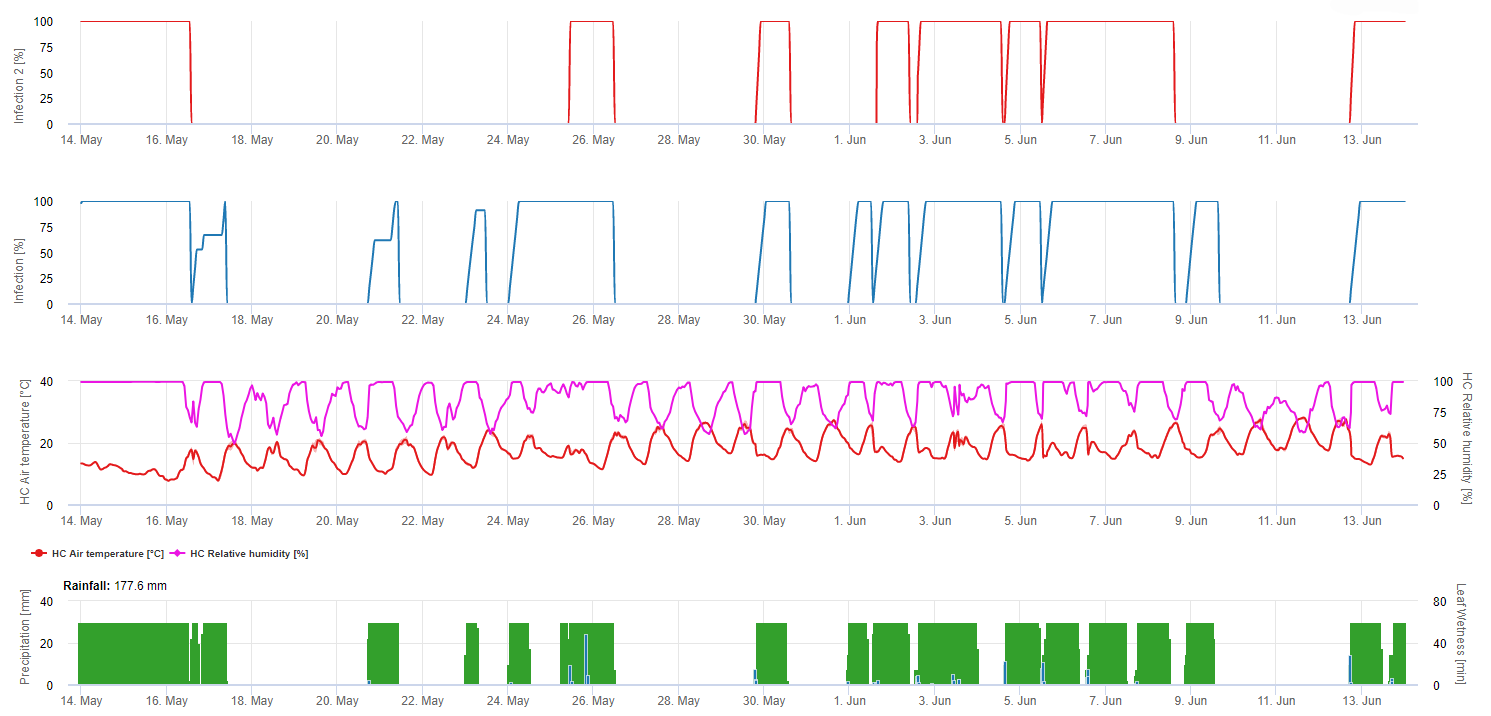
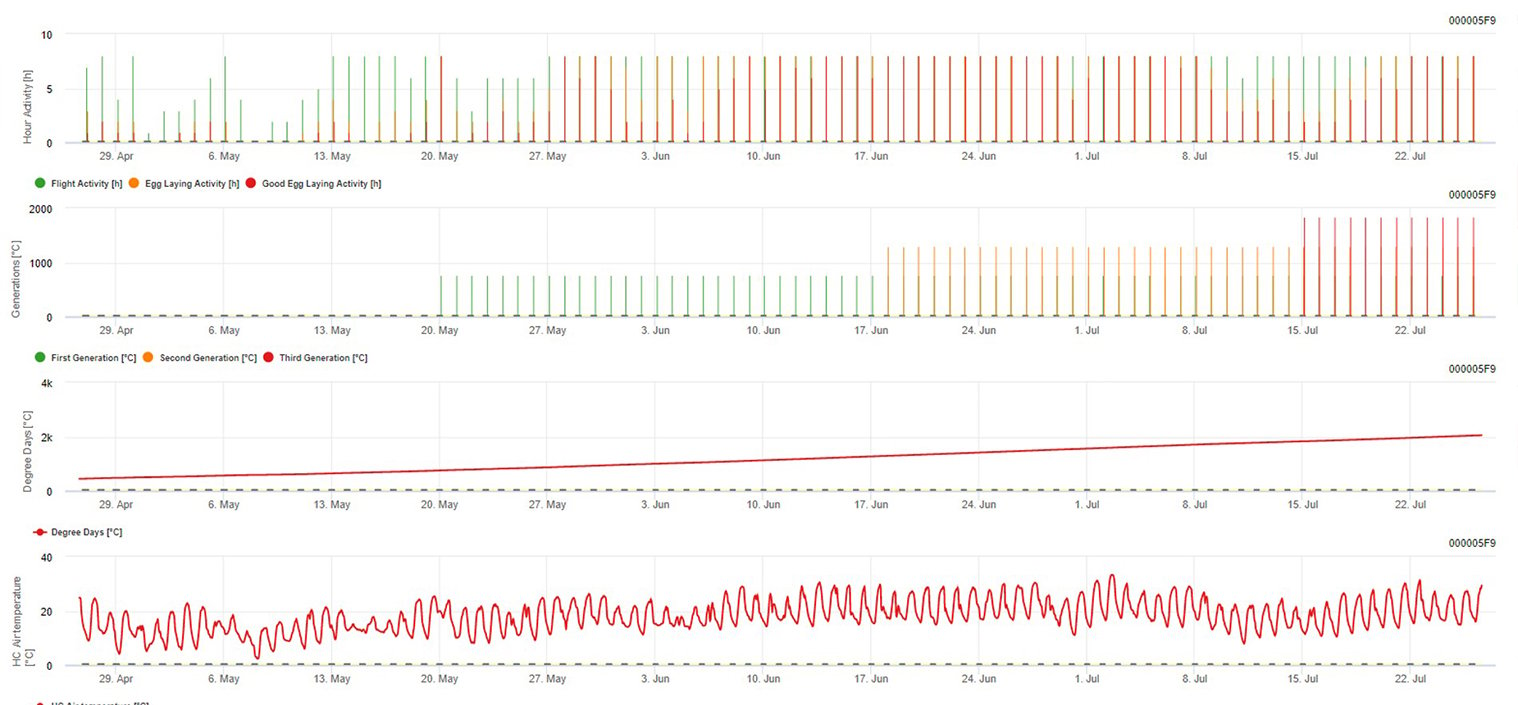
Modellierung des Europäischen Traubenwicklers in der FieldClimate-Plattform
Sensor benötigt: Lufttemperatur
Stündliche Daten Ausgabe: Flugaktivität, Legeaktivität, erste Generation, zweite Generation und dritte Generation, akkumulierte Gradtage
Für Flugaktivität, Eier und gute Legetätigkeit: Wir prüfen die Daten der letzten 8 Stunden (zwischen 19:00 und 03:00 Uhr). Der Schwellenwert für die Flugaktivität liegt bei >12°C, für die Eiablageaktivität bei > 15°C und für eine gute Eiablageaktivität bei > 17°C. Wenn alle Bedingungen für die Eiablageaktivität erfüllt sind, erzeugen wir die erste Generation (= akkumulierte Gradtage). Die erste Generation + 530 DD ist die zweite Generation (wenn nicht aufgrund der vorherigen Bedingungen vorhergesagt). Zweite Generation + 530 DD ist die dritte Generation.
In der Grafik sehen Sie das Auftreten der ersten Generation (gute Bedingungen für alle drei Aktivitäten - Flug, Eiablage und gute Eiablage) an etwa 469 Grad-Tagen akkumuliert. Das Fliegen der zweiten Generation beginnt Mitte Juni bei etwa 1010 akkumulierten Gradtagen und guten Temperaturbedingungen für die Eiablage.
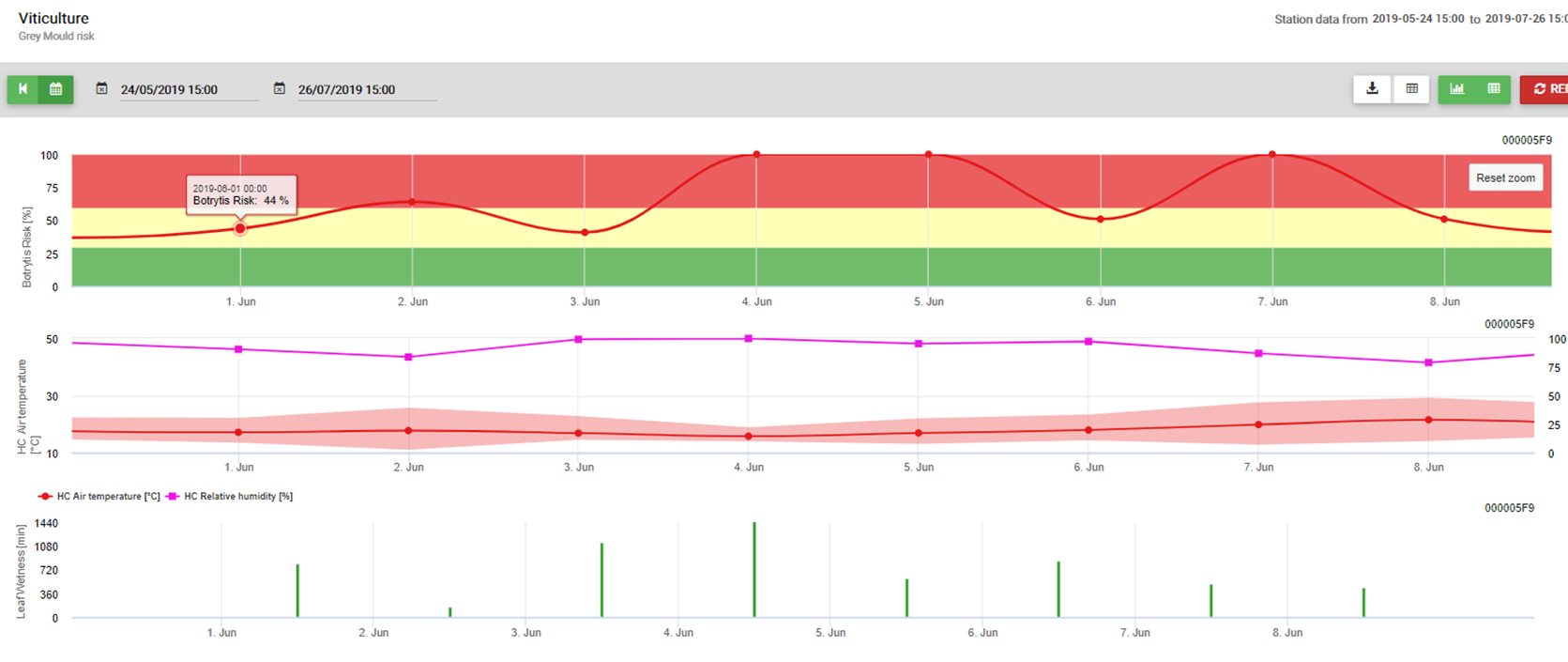
Botrytis cinerea ist ein nekrotropher Pilz, der viele Pflanzenarten befällt, dessen bekanntester Wirt jedoch die Weinrebe sein dürfte. Im Weinbau ist er allgemein als Botrytis-Traubenfäule bekannt; im Gartenbau wird er üblicherweise als Grauschimmel oder Graufäule bezeichnet.
Der Pilz führt zu zwei verschiedenen Arten von Infektionen bei Trauben. Die erste, die Graufäule, ist die Folge von anhaltend feuchten oder nassen Bedingungen und führt in der Regel zum Verlust der befallenen Trauben. Die zweite Art, die Edelfäule, tritt auf, wenn trockenere Bedingungen auf feuchtere folgen, und kann zu ausgeprägten süßen Dessertweinen wie dem Sauternes oder dem Aszú von Tokaj führen. Der Name der Art Botrytis cinerea ist vom lateinischen Wort für "Trauben wie Asche" abgeleitet; obwohl poetisch, bezieht sich das "Trauben" auf die Bündelung der Pilzsporen an ihren Konidiophoren, und "Asche" bezieht sich lediglich auf die gräuliche Farbe der Sporenmassen. Der Pilz wird in der Regel als Anamorph (ungeschlechtliche Form) bezeichnet, da die sexuelle Phase nur selten beobachtet wird. Die teleomorphe (geschlechtliche) Form ist ein Ascomycet, Botryotinia cinerea.
Biologie von B. cinerea
Bei Obst und Weintrauben haben Blüteninfektionen mit anschließender Latenz einen großen Einfluss auf die Epidemiologie von Grauschimmel. Es können mehrere Infektionswege von der Blüte zur Frucht unterschieden werden. Bei Weintrauben, Kiwis und Himbeeren wurde eine Infektion über den Griffel in die Samenanlage postuliert. Im Ovulus bleibt der Erreger latent, was auf eine vorformatierte Wirtsabwehrstrategie zurückzuführen sein dürfte (ähnlich dem Resveratrol-Gehalt der jungen Weinbeere). Bei Weintrauben, Kiwis und Erdbeeren wurden Infektionen über die Staubblätter, Blütenblätter oder Kelchblätter festgestellt. Bei Weintrauben haben Studien gezeigt, dass B. cinerea die Staubblätter infizieren kann und sich basal ausbreitet, um den Rezeptor zu infizieren, und dann systemisch auf den Stiel und das Gefäßgewebe in den Beeren wächst.
Ein sechsjähriges Forschungsprojekt in der Kapregion hat gezeigt, dass Traubenbeeren über das Stoma und die Lentizellen des Blütenstiels und im unteren Bereich der Rachis infiziert werden können. Stielinfektionen sind auch während der Blütezeit möglich. Später erhöht dieses Gewebe die Resistenz gegen B. cinerea Infektionen.
Andere Infektionswege postulieren das saprophytische Wachstum des Erregers auf Blütenresten und die spätere Infektion von Beeren, wenn die Anfälligkeit mit der Reifung oder durch Insekten- oder Hagelschäden an den Beeren zunimmt. Eine weitere These ist die Anhäufung von Konidien in der Frucht während des Sommers und die Infektion anfälliger Beeren später in der Saison. Eine Konidieninfektion von reifenden Früchten ist von jeder Inokulumquelle möglich. Höchstwahrscheinlich wird eine geringe Anzahl latent infizierter Beeren gebildet, die bei zunehmender Anfälligkeit der Beeren mit zunehmender Reife eine umfangreiche Sporenbildung aufweisen. Man kann davon ausgehen, dass Beeren ab einem Zuckergehalt von 6% anfällig werden.
Bei Kiwis haben die Pflückbedingungen einen großen Einfluss auf das Auftreten von B. cinerea. Früchte, die mit einer feuchten Oberfläche gepflückt werden, können infiziert werden durch B. cinerea an den Mikroverletzungen, die von den Fingern der Pflücker verursacht werden.
Bei der praktischen Kontrolle von B. cinerea müssen wir zwei wichtige Infektionsperioden unterscheiden: Blüte und Seneszenz. 1) Während der Blütezeit haben wir anfällige junge Früchte, bei denen die Infektion von einer Latenzzeit gefolgt wird. 2) Die Infektion reifer (seneszenter) Früchte führt zu Symptomen ohne Latenzzeit. Die Bedeutung der Infektion während der Blütezeit bei Weintrauben kann von Saison zu Saison und von Region zu Region unterschiedlich sein. Bei Früchten, bei denen eine gewisse Haltbarkeit zu erwarten ist (Tafeltrauben, Kiwis oder Erdbeeren), treten die Symptome auf, wenn sie unter gekühlten Bedingungen in Geschäften oder Lagern gelagert werden. Eine chemische Bekämpfung von Weintrauben, die eine gute Resistenz gegen B. cinerea während der Blütezeit keine wirtschaftlichen Ergebnisse zeigen wird. Daher müssen alle Risikobedingungen und die Wahrscheinlichkeit einer Infektion, die Anfälligkeit der Früchte und die Lagerfähigkeit sowie die Lagerbedingungen bei der Entscheidung über einen Antrag gegen Botrytis cinerea während der Blütezeit.
Bei Steinobst Infektion durch B. cinerea tritt meist während der Blütezeit auf. Zu diesem Zeitpunkt sind Behandlungen gegen Monilina spp. in Betracht gezogen werden, die auch die Botrytis cinerea Infektion.
Modell der B. cinerea und praktische Anwendung
Benötigte Sensoren: Blattnässe, Temperatur, relative Luftfeuchtigkeit
Botrytis cinerea ist ein fakultativer Parasit. Er wächst auch auf abgestorbenem Pflanzenmaterial. Aus diesem Grund ist er in Weinbergen und Obstplantagen immer präsent. Botrytis cinerea ist an ein feuchtes Klima gebunden. Für eine Infektion benötigt er eine sehr hohe relative Luftfeuchtigkeit oder das Vorhandensein von freiem Wasser (Sensor: Blattnässe, relative Luftfeuchtigkeit). Der Pilz ist nicht in der Lage, gesundes erwachsenes Pflanzenmaterial durch Sporen zu infizieren. Die Infektion erfolgt an jungen Trieben der Rebe während längerer feuchter Perioden oder an durch Hagelstürme beschädigten Trieben.
Das Modell verwendet die folgende Korrelation zwischen der Dauer der Blattnässe und der Temperatur, um das Risiko einer Infektion zu berechnen.
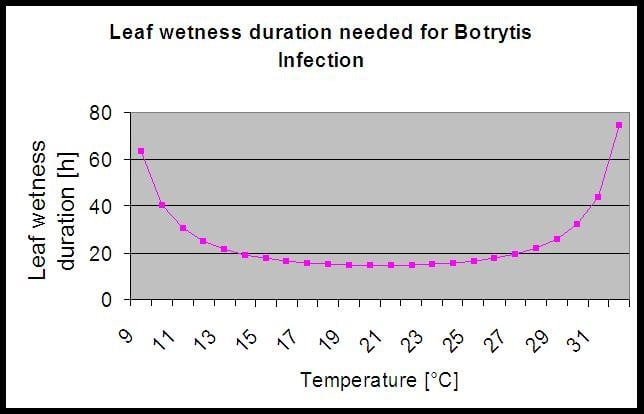
Die Botrytis Risikomodell ergibt einen Risikowert von 0 bis 100%. Dieser Wert gibt den Druck von B. cinerea zu der Zeit. Wenn wir einen Wert von 100% haben, bedeutet dies, dass es mehrere Male eine Nässeperiode gab, die lang genug war, um das anfällige Gewebe zu infizieren (wir berechnen so genannte "nasse Punkte" (Array zwischen Blattnässe, Temperatur mit einem Maximum von anfänglich 38400 Punkten (Anfang der Saison, was 30% Risiko anzeigt). Nach diesem Zeitraum erhöht jede feuchte Periode mit etwa 4000 feuchten Punkten (Array) das Risiko um 10% oder umgekehrt reduziert jede trockene Periode das Risiko um 1/5 des vorherigen Wertes.
Eine Klage gegen B. cinerea ist abhängig von der Frucht und dem Produktionsziel.
Anthraknose, Schwarzfleckenkrankheit, Vogelaugenfäule, Vogelaugenfleckenkrankheit ist weltweit verbreitet. Die Krankheit mindert die Fruchtqualität und den Ertrag und schwächt die Rebe durch eine erhebliche Zerstörung der neuen Triebe und Blätter. Die meisten Vitis-Arten sind für Anthraknose anfällig.
In feuchten Weinbaujahren ist die Krankheit gut etabliert und verursacht nach frühen Regenfällen Schäden an sehr anfälligen Sorten.
Hat sich die Krankheit erst einmal in den Weinbergen etabliert, kann sie sehr schwer zu bekämpfen sein. In Europa ist die Krankheit jedoch nahezu vernachlässigbar, da Fungizide mit Kupferverbindungen, Dithiocarbamaten und Phthalimiden zur Bekämpfung des Falschen Mehltaus eingesetzt werden und es in den trockenen Klimazonen kaum Niederschläge gibt.
Symptome
Die Krankheit befällt die oberirdischen, saftigen Teile der Rebe, einschließlich junger Triebe, Blätter, Blattstiele, Ranken und Trauben; am häufigsten treten Läsionen an Trieben und Beeren auf. Die ersten Symptome treten als vereinzelte, kleine, kreisförmige Flecken auf, die sich bräunlich verfärben, mit grauen Zentren und dunklen, runden oder eckigen Rändern eingesunken sind. Manchmal sehen die Symptome wie Hagelverletzungen aus. Auf den Blättern erscheinen zahlreiche kleine, braune Flecken, die in der Mitte grau sind; das nekrotische Zentrum fällt in der Regel heraus, wodurch ein "Einschussloch" entsteht. Junge Blätter sind anfälliger für Infektionen. Auf Beeren bilden sich kleine rötlich-braune, kreisförmige Flecken, die leicht eingesunken sind. Das Zentrum wird grau und ist von rötlich-braunen bis schwarzen Rändern umgeben (Vogelauge).
Lebenszyklus des Erregers
Verursacht wird die Krankheit durch den Erreger Elsinöe Ampelina (de Bary) Schere.
Infizierte Stöcke sind die Hauptquelle der Krankheit. Sklerotien oder Myzel, die in den Läsionen überleben, werden im Frühjahr aktiv und produzieren Konidien unter feuchten Bedingungen (Regen oder Tau für 24 Stunden) bei einer Temperatur von 2°C-40°C. Die Konidien werden durch Regen auf neues Gewebe geschleudert. Sie keimen aus und erzeugen neue Infektionen. Warmes Wetter verkürzt die für die Erstinfektion erforderliche Benetzungszeit und die Inkubationszeit, bevor Symptome auf den Blättern sichtbar werden. Die optimale Sporenkeimung findet bei 25-30 °C statt, wobei die erforderliche Blattnässe mindestens 3 bis 4 Stunden beträgt. Auch die Inkubationszeit ist in diesem Temperaturbereich am kürzesten (3-4 Tage unter ideal feuchten Bedingungen). Bei Infektionen bei Temperaturen um 10 °C benötigen die Krankheitssymptome etwa 14 Tage, um auf den Blättern sichtbar zu werden.
Konidien oder Ascosporen, die sich auf infizierten Beeren bilden, die an der Rebe oder am Boden überwintern, können ebenfalls Primärinfektionen verursachen. Die Sporen infizieren neue Blätter, Triebe, Ranken und junge Beeren und bilden bei feuchten Bedingungen Läsionen. Diese Konidien dienen als sekundäres Inokulum und sind für weitere Infektionen im Laufe der Saison verantwortlich. Die Konidien werden durch Regentropfen aufgewirbelt und im Weinberg verteilt.
Die Entwicklung der Epidemie hängt von der Anfälligkeit des Rebengewebes, der Ausbreitung der Konidien und den vorherrschenden Wetterbedingungen ab. Der wichtigste Faktor ist die Häufigkeit von Regenperioden.
Modellierung in Fieldclimate
Entwicklung der überwinternden Sporen: Temperatur: 2-40C°, relative Luftfeuchtigkeit über 90% oder Blattnässe- wenn die Sporen entwickelt sind (100%) und noch Bedingungen der Blattnässe und Temperatur, beginnt die Infektion zu berechnen. Die Sporenentwicklung (r.h. unter 50% ) und die Infektion werden nicht mehr berechnet, wenn die Feuchtigkeit zu niedrig ist. Die Schwere der Infektion hängt von den feuchten Bedingungen (Regenereignis) ab.