





Kirsche Krankheitsmodelle
Die wichtigste Krankheit bei Kirschen ist Braunfäule, verursacht durch den Pilzerreger Monilia laxa. Eine weitere wichtige Krankheit, die ebenfalls bei hoher Feuchtigkeit auftritt, ist die Krankheit Einschussloch, verursacht durch den Pilzerreger Wilsonmyces carpophilu und die Krankheit der Kirschblattfleck, verursacht durch Blumeriella jaapii.
Regen Akkumulation
Starker Regen wäscht die auf den Blättern von Reben oder anderen Pflanzen verbliebenen Pestizide ab. Die Regenhärte moderner Fungizide hat sich seit 1980 stark verbessert. Heute können wir davon ausgehen, dass die modernsten Fungizide bis zu 30 mm Regen aushalten, wenn sie die Möglichkeit hatten, auf dem Blatt zu trocknen. Wenn der Regen unmittelbar nach dem Sprühen oder während des Sprühens einsetzt, kann die Regenhärte stark reduziert sein.
Bei altmodischen Kontaktfungizidformulierungen müssen wir mit einer Regenhärte von weniger als 12 mm rechnen. So wie wir es in den 1970er Jahren gewohnt waren. Um die Blätter in einem Weinberg zu benetzen, braucht es etwa 2 mm Regen. Deshalb kumulieren wir in diesem Modul nur Niederschläge, die größer als 2 mm innerhalb einer Blattbefeuchtungsperiode sind. Das bedeutet, dass es an einem einzigen Tag insgesamt 6 mm Regen geben könnte, aber dieses Modul akkumuliert nichts davon, weil die Blätter schon wieder trocken sind, bevor es 2 mm geregnet hat.
Der Regen wird für 3, 5 und 7 Tage akkumuliert. Über einen längeren Zeitraum ist das Pflanzenwachstum viel wichtiger für die Wirkung von Kontaktfungiziden als die Regenresistenz der Mittel.
Modell des Steinobstschorfs
Steinobstschorf wird durch einen pflanzenpathogenen Pilz ausgelöst Cladosporium carpophilum. Der Erreger tritt an Pfirsichen, Nektarinen, Aprikosen und Pflaumen auf, wobei die Verluste bei Pfirsichen im Allgemeinen größer sind als bei den anderen Früchten.
Die Krankheit befällt Zweige, Blätter und Früchte. Die schwerwiegendsten Schäden entstehen bei Infektionen der Früchte.
Symptome
Die Fruchtläsionen beginnen als kleine, runde, grünliche Flecken. Diese Flecken treten in der Regel erst auf, wenn die Früchte halb ausgewachsen sind, auch wenn die Infektion schon früher in der Saison erfolgt ist (etwa sechs bis sieben Wochen nach dem Blütenblattfall). Ältere Läsionen haben einen Durchmesser von etwa 1/4 Zoll und sehen staubig oder samtig grün aus. Die zahlreichen Läsionen befinden sich in der Regel in der Nähe des Stielendes der Frucht (diese Stelle ist der Sonne ausgesetzt). Ausgedehnte Fleckenbildung kann zu Fruchtrissen führen, die als Eintrittspforten für verschiedene Fruchtfäulepilze dienen. Die Früchte können auch vorzeitig abfallen oder nicht gut gelagert werden.
Auch die Blätter können infiziert sein. Kleine, runde und gelblich-grüne Flecken treten auf der Unterseite des Blattes auf. Das Pflanzengewebe kann eintrocknen und abfallen, wobei Einschusslöcher zurückbleiben. In der Regenzeit fallen die infizierten Blätter in der Regel früh ab.
An Zweigen beginnen Krebsgeschwüre als kleine, rötliche Läsionen am Wachstum der laufenden Saison. Sie breiten sich langsam aus und sind möglicherweise erst im Hochsommer sichtbar. Die kleinen Krebsgeschwüre haben unregelmäßige Ränder, verursachen aber keine eingesunkenen Stellen auf der Rinde.
Auf den Zweigen der das Myzel (oder die Konidien) überwintert in Form von dunkelbraunen, kugelförmigen Zellen. Aus dem überwinternden Myzel werden im Frühjahr Konidien produziert, die auf die Blätter und Früchte übertragen werden durch Wind oder durch Regen. Die Bedingungen, die die Entwicklung der Krankheit begünstigen, sind Temperaturen über 16°C für die Sporenproduktion, über 10°C (optimal 22°C bis 27°C) für die Sporenkeimung und zwischen2°C und 35°C für die Krankheitsentwicklung. Die Keimung und das Eindringen in das Pflanzengewebe erfolgen in Kürze. Inokulationen und Infektionen finden bis etwa einen Monat vor der Fruchtreife statt. Während der Pilz auf der Frucht wächst, setzt sich das Myzel dicht an der Oberfläche zwischen den Haaren fest und bildet eine Matte aus kurzen, plumpen Zellen, aus denen sich Konidiophoren und Konidien bilden. Das Fruchtfleisch des Pfirsichs wird nicht durchdrungen, aber der enge Kontakt des Pilzes mit den äußeren Zellen ermöglicht die Aufnahme von Nährstoffen aus der Frucht durch die unversehrten Wände. Offensichtlich sind die äußeren Zellen in gewissem Maße geschädigt.
Bei FieldClimate wird das Risiko einer Caldosporium carpophilum-Infektion bestimmt durch feuchte Bedingungen im Frühjahr und Frühsommer nach dem Abfallen der Blütenblätter. Die Krankheit ist in der Regel schwerwiegender bei tiefliegende, schattige und feuchte Gebiete mit geringe Luftbewegung.
Bei FieldClimate stellen wir Infektionen in einem Temperaturbereich von 7 bis 24°C fest, mit einem Temperaturoptimum um 20°C.
Das FieldClimate Modell berechnet in Abhängigkeit von Blattnässedauer und Temperatur ein Risikomodell von Cladosporium carpohilum.
Einschussloch
Das Einschussloch wird durch den Pilzerreger Wilsonmyces carpophilu (Kraut- und Knollenfäule).
Am stärksten bei Aprikosen, tritt aber bei allen Steinfrüchten auf. Der Pilz befällt die Blätter, Zweige und Früchte.
Symptome
Befallene Blätter zeigen kleine braune Flecken mit rötlichen Rändern (etwa 1 mm Durchmesser), die sich zu größeren kreisförmigen Läsionen (etwa 3 mm Durchmesser) ausweiten. Diese Flecken trocknen ein und fallen aus dem Blatt heraus, was wie ein Einschussloch aussieht. Die befallenen Zweige weisen deutlich geschnittene braune Ränder mit einem nekrotischen Zentrum auf, das nicht abfällt, sondern große Mengen von Schleim absondert. Im weiteren Verlauf wird die Verholzung der infizierten Zweige behindert, und die Läsionen wachsen zu Krebsen heran. In schweren Fällen kann es zum vorzeitigen Absterben des Baumes kommen.
Die Früchte zeigen zunächst kleine kreisförmige, tiefviolette Flecken. Wenn die Krankheit fortschreitet, unterscheiden sich die Symptome je nach Fruchtart. Bei Aprikosen werden die Flecken braun, erhaben und rau, wodurch die Früchte eine schorfige Oberfläche bekommen. Bei Pfirsichen und Nektarinen entwickelt sich der Schorf zu tiefen Einbuchtungen.
Befallene Früchte weisen Flecken und in schweren Fällen Risse in der Schale auf.
Der Schrotschusspilz überlebt in infizierten Knospen. Er ist in der Lage, Blätter, Stängel und Früchte zu infizieren, während kaltes, regnerisches Wetter Perioden im Frühjahr und Herbst. Es sind Regenperioden erforderlich, um gesunde Pflanzenorgane zu infizieren.
Der Pilz ist in der Lage, mehrere Jahre in den Kanälen oder Knospen der infizierten Zweige zu überdauern. Wenn die Bedingungen günstig sind, kann er weiter wachsen, auch im Winter. Im Frühjahr werden die Konidien durch Rinde auf Blüten und junge Blätter geschleudert und infizieren diese. In ungünstigen Perioden (Trockenheit) sind die Konidien noch mehrere Monate lang lebensfähig. Regen ist notwendig für die Ausbreitung und feuchte Bedingungen sind für die Keimung erforderlich. Der Pilz ist in der Lage, über 2 °C zu wachsen.
Literatur:
- Adaskaveg JE, Ogawa JM, Butler EE (1990) Morphology and ontogeny of conidia in Wilsonomyces carpophilus, gen. nov. and comb. nov., causal
Erreger der Schrotschusskrankheit von Prunuss-Arten. Mycotaxon 37, 275-290. - Ashkan M, Asadi P (1971) Shot hole of stone fruits in Iran. Iranian Journal of Plant Pathology 7, 39-63.Esfandiari E (1947) Les maladies des plantes cultivées et des arbres
fruitiers des régions subtropicales du nord de l'Iran. Entomologie et Phytopathologie Appliquées 5, 2. - Grove GG (2002) Einfluss von Temperatur und Feuchtigkeitsdauer auf die Infektion von Kirschen- und Pfirsichblättern durch Wilsonomyces carpophilus. Kanadische
Zeitschrift für Pflanzenpathologie 24, 40-45. - Ogawa JM, Zehr EI, Bird GW, Ritchie DF, Uriu K, Uyemoto JK (1995) Compendium of stone fruit diseases. (Die Amerikanische Phytopathologische
Society Press: St Paul, MN)
Echter Mehltau
Echter Mehltau ist eine häufige Krankheit bei vielen Pflanzenarten. Mehrere Mehltaupilze verursachen ähnliche Krankheiten an verschiedenen Pflanzen (wie z. B. Podosphaera Arten auf Apfel und Steinobst; Sphaerotheca-Arten auf Beeren und Steinobst; Erysiphe necator an Weinstöcken). Echte Mehltaupilze benötigen in der Regel feuchte Bedingungen um überwinternde Sporen freizusetzen, die dann keimen und Pflanzengewebe infizieren können. Der Pilz benötigt jedoch keine Feuchtigkeit, um sich zu etablieren und nach der Infektion der Pflanze zu wachsen. Echter Mehltau bevorzugt normalerweise warmmediterranes Klima.
Echter Mehltau ist an den meisten Pflanzen leicht zu erkennen durch das weiße bis graue pulverförmige Myzel und Sporenwachstum, das sich auf beiden Seiten von Blättern, Blüten, Früchten und an Trieben bildet. An Baumfrüchten bildet sich ein rauer, korkiger Fleck auf der Schale, wo die Infektion stattgefunden hat.
Alle Mehltaupilze benötigen lebendes Pflanzengewebe, um zu wachsen. Auf laubabwerfenden, mehrjährigen Wirtspflanzen wie Weinreben, Himbeeren und Obstbäumen ist der Echte Mehltau überlebt von einer Saison zur nächsten in infizierte Knospen oder als Fruchtkörper Chasmothecien genannt, die sich auf der Rinde von Kordonen, Ästen und Stämmen befinden.
Die meisten Mehltaupilze wachsen als dünne Myzelschicht auf der Oberfläche des befallenen Pflanzenteils. Die Sporen, die das Hauptverbreitungsmittel sind, machen den Großteil des pulverförmigen Wachstums aus und werden in Ketten produziert, die mit einer Handlinse gesehen werden können. Im Gegensatz dazu wachsen die Sporen des Falschen Mehltaus an verzweigten Stängeln, die wie kleine Bäume aussehen. Auch die Kolonien des Falschen Mehltaus sind grau statt weiß und treten meist auf der unteren Blattoberfläche auf.
Die Sporen des Echten Mehltaus werden übertragen durch Wind auf Wirtspflanzen. Obwohl die Feuchtigkeitsanforderungen für die Keimung variieren, können viele Mehltauarten keimen und infizieren in Abwesenheit von Wasser. Tatsächlich werden die Sporen einiger Mehltaupilze abgetötet, und die Keimung und das Myzelwachstum werden durch Wasser auf Pflanzenoberflächen gehemmt. Gemäßigte Temperaturen und Schatten sind im Allgemeinen die günstigsten Bedingungen für die Entwicklung von Mehltau, da Sporen und Myzel empfindlich auf extreme Hitze und direkte Sonneneinstrahlung reagieren.
Der Pilz überwintert als Myzel in den Knospenschuppen, die Primärinfektion erfolgt beim Austrieb der Blätter aus diesen infizierten Knospen. Sekundärinfektionen treten auf, wenn Konidien, die durch Primär- und nachfolgende Sekundärinfektionen gebildet werden, auf anfälliges Gewebe geweht oder durch Regen aufgespritzt werden. Früchte (vor der Aushärtung der Kerne) und sukkulentes Endwachstum sind anfällig für Infektionen.
Die durchschnittlichen Minimal-, Optimal- und Maximaltemperaturen für S. pannosa liegen bei 5°, 24° und 24°C. In trockener Luft werden bei allen Temperaturen viel mehr Konidien gebildet als in feuchter Luft (C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli; 1954)
Echter Mehltau tritt bei ähnlicher relativer Luftfeuchtigkeit und ähnlichen Temperaturen wie der Kirschenmehltau auf.
Literatur:
- C.E. Yarwood, Soliman Sidky, Morris Cohen, Vincent Santilli (1954): Temperature relations of Powdery Mildews. HILGARDIA. A Journal of Agricultural Science Published by the California Agricultural Experiment Station. Universität von Kalifornien. Band 22/Nummer 17.
Taphrina-Blattrolligkeit
Pfirsichblattwucherung (Pilzerreger: Taphrina deformans) ist eine Pilzkrankheit, die bei fast allen Pfirsich- und Nektarinensorten zu schwerem, frühem Blattverlust und Ernteeinbußen führen kann.
Symptome
Das häufigste und auffälligste Symptom der Blattrolligkeit tritt an der Blätter (Blattwerk). Befallene Blätter sind stark deformiert und weisen oft eine Vielzahl von Farben auf (hellgrün und gelb bis hin zu Rot- und Violett-Tönen). Der Pilz bewirkt, dass sich die meristematischen Zellen an den Blatträndern schnell und wahllos vermehren, was dazu führt, dass die Blätter unterschiedlich faltig, runzlig und eingerollt werden (Foto 2). Wenn diese infizierten Blätter reifen, bilden sich auf der Oberfläche nackte Asci, die Ascosporen des Erregers enthalten und ihnen ein staubiges Aussehen verleihen, woraufhin die Blätter braun werden, verschrumpeln und vom Baum abfallen.
Viele befallene Früchte früh abfallen und unbemerkt bleiben; die, die übrig bleiben, können zu schief am Stielende wie ein kleiner gelber Kürbis, während andere rötlich bis violett werden und "warzenartige" Verformungen auf der Oberfläche haben.
Krankheitszyklus
Der Erreger kommt fast überall vor, wo Pfirsiche angebaut werden. Der pilzliche Erreger überwintert als Konidien (Blastosporen, "hyphenartige" Sporen) an geschützten Stellen in der Rinde und um die Knospen herum. Primärinfektionen treten im zeitigen Frühjahr auf. Ab dem Anschwellen der Knospen bis zum Erscheinen der ersten Blätter an den Knospen. Infektionen an jungen Pfirsichblättern treten auf bei Temperaturen von 10°C bis 21°C. Unter 7°C treten nur wenige Infektionen auf. Infektionen treten hauptsächlich auf, wenn Regen die überwinterten Sporen in die Knospen waschen und kalte Temperaturen die Entwicklungszeit der Blätter zu verlängern (sie sind dem Krankheitserreger lange ausgesetzt, bevor sie voll entwickelt sind und dem Eindringen des Pilzes widerstehen können). Wenn Temperaturen nach dem Anschwellen der Knospen sind warm und die Blätter entwickeln sich schnellInfektionen selten sich etablieren, auch wenn es im Frühjahr regnet. Nässe durch Regen (oder andere Faktoren) über mehr als 12,5 Stunden ist für eine Blattinfektion erforderlich, allerdings nur, wenn die Temperatur während der nassen Periode unter 16 °C liegt. Die maximale Infektion tritt auf, wenn die Bäume 2 Tage oder länger nass sind, was westlich der Kaskaden häufig der Fall ist. Wenn die Temperaturen über 21 °C (69 °F) bleiben, treten trotz der Infektion möglicherweise keine Symptome auf. Die Früchte sind nach dem Abfallen der Blütenblätter anfällig, bis die Lufttemperatur über 19 °C liegt. Für eine Infektion der Früchte sind Niederschläge von 0,5 Zoll und eine Nässe von 24 Stunden erforderlich.
Das Risiko einer Taphrina defomans In FieldClimate wird die Infektion auf zwei Arten berechnet:
- anhand von Temperaturwerten (altes Modell).
- unter Verwendung der Niederschlagsmenge der letzten Stunden und der Temperatur während dieses Zeitraums. Darüber hinaus umfasst dieses Modell auch die Inkubationszeit (Zeit, in der die Symptome auf dem Feld zu sehen sind), wobei für die Berechnungen Temperaturen unter 19 °C verwendet werden.
Beide Modelle sind in FieldClimate unter "Taphrina Leaf Curl disease" erhältlich.
Blattfleckenkrankheit
Die Kirschblattfleckenkrankheit wird verursacht durch den Pilz Blumeriella jaapii. Die Krankheit reduziert die Blüte und schwächt den Baum. Die Kirschblattfleckenkrankheit entblättert den Baum oft schon im Hochsommer. Wiederholtes Entblättern macht den Baum anfälliger für Winterschäden und kann ihn schließlich abtöten.
Symptome
Die Blätter zeigen im Frühsommer kleine violette bis braune Flecken mit deutlichen Grenzen. Die Blätter werden gelb und fallen ab. Im Juli fallen die Zentren der befallenen Flecken häufig aus, was ein schusslochartiges Aussehen ergibt.
Ursache
Der Pilz überwintert auf Laub. Im Frühjahr nach feuchtem Wetter bilden sich Sporen, die vom Wind verweht werden und Blätter infizieren. Wenn die Temperaturen günstig sind (16°C bis 24°C), erfordert die Infektion eine nasse Periode von nur wenigen Stunden. Die Blätter sind anfällig, wenn sie vollständig entfaltet sind - dies geschieht in der Regel kurz vor dem Blattfall. Die ersten Blattinfektionen bilden Flecken, und in den Flecken werden weitere Sporen produziert. Diese Sporen werden durch Regen auf andere Blätter gespritzt und infizieren sie. Die sekundäre Ausbreitung und Infektion durch Sporen setzt sich bei feuchtwarmem Wetter immer wieder fort, bis die Blätter im Herbst fallen.
Lebenszyklus
Die Kirschblattfleckenkrankheit wird durch den Ascomycetenpilz Blumeriella jaapii (früher bekannt als Coccomyces hiemalis) anamorph Phloeosporella padi (Lib.) Arx. Dieser Pilz überwintert in totem Laub auf dem Boden als seine Hauptüberlebensform. Im Frühjahr entwickeln sich auf diesen Blättern die Apothecien. In diesen Pilzfruchtkörpern werden Ascosporen gebildet, die gewaltsam ausgestoßen werden während Regenperioden für etwa 6-8 Wochen, beginnend mit dem Blattfall. Dies ist das telemorphe oder sexuelle Stadium und führt zu einer neuen genetischen Rekombination und Variation der Krankheit. Dieser Krankheitserreger befällt vor allem die Blätter von Kirschbäumen und ist dort am auffälligsten. Am auffälligsten ist die Symptom dieser Krankheit ist die Vergilbung der Blätter vor dem Abfallen im zeitigen Frühjahr. Befallene Blätter sind gefüllt mit zahlreichen schwarze Läsionen die einen deutlichen Teil des Blattes bedecken. Diese Läsionen erscheinen zunächst als kleine violette Flecken, die sich rot bis braun und dann schwarz verfärben. Die meisten Flecken sind kreisförmig, können aber auch zu großen, unregelmäßigen toten Flecken zusammenwachsen. Diese Flecken neigen dazu, sich zu vergrößern und sich vom gesunden Gewebe zu lösen. Schließlich fallen sie aus dem Blatt heraus, was ein "Einschussloch"-Erscheinungsbild ergibt. Dieser "Einschussloch"-Effekt tritt bei Sauerkirschen häufiger auf als bei Süßkirschen. In schwereren und fortgeschrittenen Fällen finden sich Läsionen an den Früchten, Blattstielen und Stängeln. Blätter mit Läsionen fallen in der Regel früh in der Vegetationsperiode ab, und es ist üblich, dass stark befallene Bäume im Hochsommer entlaubt sind. Stark befallene Blätter werden sehr gelb, bevor sie abfallen.
Wenn die Krankheit nicht bekämpft wird und über mehrere Jahre anhält, kann es zu verschiedenen negativen Auswirkungen auf die gesamte Kirschproduktion eines Baumes kommen, die in erster Linie auf den Zyklus der frühzeitigen und wiederholten Entlaubung zurückzuführen sind. Zu diesen Auswirkungen gehören kleinwüchsige und ungleichmäßig gereifte Früchte mit schlechtem Geschmack, eine höhere Anfälligkeit der Bäume für Winterschäden, das Absterben der Fruchttriebe, kleine und schwache Fruchtknospen, eine geringere Fruchtgröße und ein geringerer Fruchtansatz und schließlich das Absterben des Baumes. Wenn die Krankheit die Blätter im zeitigen Frühjahr stark genug infiziert hat, ist es möglich, dass die Früchte in diesem Jahr nicht ausreifen. Diese Früchte bleiben in der Regel hell, weich und haben einen sehr geringen Gehalt an löslichen Feststoffen und Zucker.
Krankheitszyklus
Primäre Infektion
Dieser Pilz überwintert in abgestorbenen Blättern auf dem Boden. Im zeitigen Frühjahr (um den Blattfall) entwickeln sich in diesen Blättern Pilzfruchtkörper, Apothecien genannt. In den Apothecien werden Sporen (Ascosporen) gebildet, die ab dem Blattfall gewaltsam ausgestoßen werden. Diese Ascosporen werden durch Wind oder Regen auf die gesunden, neuen, grünen Blätter verbreitet und dienen so als primäres Inokulum in einer neuen Wachstumssaison. Die Ascosporen können auf dem Blatt haften bleiben und keimen, wenn ein Wasserfilm und eine optimale Temperatur vorhanden sind, innerhalb von Stunden nach dem Anhaften. Nach der Keimung können die Ascosporen durch die Spaltöffnungen (natürliche Öffnungen) an der Blattunterseite in das Blatt eindringen. In unreife Spaltöffnungen können die Ascosporen nicht sehr gut eindringen. Diese Ascosporen wachsen in den Zellzwischenräumen des Blattes. Die Inkubationszeit hängt von der verfügbaren Feuchtigkeit und den Temperaturen ab. Die kleinen violetten Läsionen können innerhalb von 5 Tagen auftreten, wenn die Bedingungen feucht sind und die Temperatur konstant zwischen 60 (15°C) und 68 Grad F (20°C) bleibt. Die Inkubationszeit kann jedoch bis zu 15 Tage dauern, wenn niedrigere Temperaturen und trockenere Bedingungen herrschen. Im Allgemeinen werden die Blätter weniger anfällig für die Krankheit, je älter sie im Laufe der Vegetationsperiode werden. Die ersten Anzeichen der Krankheit können sich an den bodennahen Trieben zeigen. Dies gilt als der primäre Reproduktionszyklus der Kirschblattfleckenkrankheit. Die Infektion durch diesen primären Zyklus ist jedoch relativ gering, aber für den Erreger unerlässlich, um sekundäre Sporen produzieren zu können.
Sekundärinfektion
Sobald sich diese Läsionen der Primärinfektion gebildet haben, besteht die Möglichkeit einer Sekundärinfektion. Sekundär- oder Sommersporen, sogenannte Konidien, bilden sich auf der Unterseite des Blattes aus leicht konkav geformten Ausstülpungen, den Acervuli. Diese Konidien sind hornförmig und gut wasserlöslich. Die Sporen haben auf der Unterseite des Blattes ein weißliches Aussehen. Sie sind Verbreitung durch Wind oder Regen wenn sie aufgebrochen werden, und können weitere Infektionen verursachen. Jede dieser Sporen kann sich vermehren und in relativ kurzer Zeit Tausende von zusätzlichen Konidien produzieren. Alle diese Konidien enthalten genau dieselbe genetische Information und sind nichts anderes als die Replikation der DNA des Pilzes. In diesem Stadium überwältigt die Pilzinfektion die Wirtspflanze und führt dazu, dass die Pflanze ihre Blätter vorzeitig abwirft. Die dabei entstehende Laubstreu wird dann zum Überwinterungsort des Pilzes, und der Zyklus beginnt im nächsten Frühjahr. Durch diese Sekundärinfektionen kann sich die Krankheit sehr schnell zu einer Epidemie auswachsen.
Umwelt
Die Krankheit ist allgemein in den USA, Kanada und verschiedenen Teilen der Welt verbreitet, wo feuchte Bedingungen auftreten. Der Kirschblattfleckenkrankheitspilz bevorzugt mäßig feuchte Bedingungen mit Temperaturen von über 15°C (60 Grad Fahrenheit). Der optimale Temperaturbereich für die Ausbreitung dieses Pilzes liegt zwischen 60 (15) und 68 Grad Celsius (20°C). Eine ernsthafte Infektion eines Baumes tritt in Jahren mit vielen Regenperioden und kühleren Sommern auf. Bei Temperaturen unter 7 °C (46 °F) werden nur sehr wenige Ascosporen aus den Apothecien ausgestoßen. Diese Krankheit gedeiht in Gegenden, in denen keine Hygienemaßnahmen zur Verhinderung der Ausbreitung der Krankheit angewandt werden. Bei der Auswahl eines Standorts für eine Obstplantage sollten Sie Gebiete mit gut entwässerten Böden, viel Sonnenlicht und guter Luftzirkulation wählen.
Modellierung der Infektion durch Blumeriella jaapii
Das Ascopsore-Reifungsmodell: Bei optimalen Temperaturen reifen die Ascosporen innerhalb von 2 Wochen. Optimale Temperaturen liegen über 16°C. Bei suboptimalen Bedingungen kann der Zeitraum der Ascosporenentladung bis zu 6 Wochen dauern. Das Modell zeigt den Zeitraum des möglichen Ascosporenausstoßes. In diesem Zeitraum ist mit Primärinfektionen der Krankheit zu rechnen.
Das Modell des Ascosporenausstoßes:
Der Ascosporenauswurf ist abhängig von der Befeuchtung der Apothecien und dem anschließenden Austrocknen. Er ist größer, wenn mehr Apothecien benetzt werden (stärkerer Regen oder mehr Regen) und wenn die Temperatur während und nach dem Regen höher ist. Er beginnt bei Temperaturen von 16°C.
Das Infektionsmodell für Blummeriella jaapii:
Die Infektion wird anhand der Tabelle von Eisensmith und Jones (Tabelle 1) vorhergesagt, die Folgendes verwendet Dauer der Blattnässe und Lufttemperatur Daten. In dem Modell wird eine Benetzungsperiode eingeleitet, wenn der Blattnässesensor nass wird. Die Benetzungsperiode endet, wenn die rF unter 90% fällt und der Blattnässesensor trocken ist. Wird der Blattnässesensor jedoch innerhalb von 8 Stunden nach dem Zeitpunkt, an dem die relative Luftfeuchtigkeit unter 90% gefallen ist, erneut nass, so wird dies als dieselbe Nässeperiode betrachtet, wobei die Trockenstunden ebenfalls hinzugefügt werden. Eine Nässeperiode kann dazu führen, dass keine Infektion oder eine Infektion mit dem Erreger der Kirschfleckenkrankheit vorliegt.
Verwendung des Modells:
Das Kirschblattfleckenmodell kann als Leitfaden dienen, um den Bedarf an Ausrottungssprays, wenn die Schutzmittelabdeckung während einer Regenepisode schwach war. Das Modell sollte nicht als routinemäßiger Leitfaden für die Anwendung von Ausrottungsspritzen verwendet werden - ein Schutzspritzprogramm ist der zuverlässigste Ansatz zur Bekämpfung der Kirschblattflecken. Das Modell ist nützlich für den Vergleich der vorhergesagten Blattfleckeninfektionsaktivität zwischen den Jahren.
Ascosporen- und Konidieninfektionsmodell auf der Grundlage von:
- EISENSMITH, S.P. und A. L. Jones (1981): Infection Model for timing fungicide applications to control cherry leaf spot. Plant Dis. 65
- Eisensmith, S. P. und A. L. Jones. 1981. A model for detecting infection periods of Coccomyces hiemalis on sour cherry. Phytopathology.71:728-732.
- Jones, A. L. und T. B. Sutton. 1996. Krankheiten von Baumfrüchten im Osten. Northcentral Regional Bulletin 45, Michigan State University
- Shane, B. (2011): Über Enviro-weather's Cherry Leaf Spot Report, siehe dort hier.
Monilia spp.
Braunfäule, verursacht durch Monilia spp. (Monilia laxa, Monilia fructigena und Monilia fructicola) gehören zu den zerstörerischsten Krankheiten an Steinobst in Europa. Besonders in der Kirschenproduktion sind die wirtschaftlichen Auswirkungen erheblich.
Symptome
Die Symptome der Braunfäule sind die Fäulnis der Kirschblüte und der grünen Zweigspitze, die durch das Eindringen des Erregers in die geöffnete Blüte durch die Narbe des Stempels oder der Staubbeutel entsteht. Dies führt in der Regel dazu, dass der gesamte Teil eines einjährigen Zweiges welk wird. Die Blätter beginnen herabzuhängen, später werden sie braun und starr, fallen aber normalerweise nicht auf den Boden, sondern bleiben bis zum Frühjahr des nächsten Jahres am Baum. Manchmal, vor allem unter feuchten Bedingungen, sind Schleimtropfen zu sehen, die ein Zeichen für die Besiedlung mit dem Pilz sind, ebenso wie die etablierten Krebsgeschwüre. Die befallenen Früchte sind mit Fäulnisflecken bedeckt, aus denen warzige Sporodochien (Hyphe) mit Konidien der "Sommerform" erscheinen. Im Spätherbst und Winter produziert der Pilz außerdem Sporodochien der "Winter"-Form auf infizierten Zweigen. Mit der Zeit werden stark befallene Früchte mumifiziert. Das in solchen Mumien wachsende Myzel verdichtet sich allmählich zu Sklerotien. Solche Früchte bleiben den Winter über am Baum.
Temperatur und Nässedauer sind wichtige Umweltfaktoren, die die Häufigkeit der Infektion mit M. laxa auf Süßkirschblüten bestimmen. Monilia laxa ist gut geeignet für die relativ niedrige Temperaturen im Frühjahr und verursachen Infektionen bei Temperaturen von bis zu 5°C innerhalb eines sehr kurzen Zeitraums der Nässedauer. Die Infektion der aktiven Blüte durch die Stima braucht nicht sehr viel Blattnässe. Blattnässe wird nur für die Keimung der Konidien benötigt. Daher sind für die Infektion der jungen Früchte längere Perioden mit Blattnässe erforderlich. Um die junge Frucht zu infizieren, muss eine Appressorie gebildet werden, und es wird freie Feuchtigkeit benötigt, um den Druck zur Bildung des Infektionsstifts aufzubauen, der in die Epidermiszelle eindringt. Mit zunehmender Reife der Früchte ermöglichen kleine Narben auf den Früchten wieder eine Infektion ohne Infektionszapfen und die benötigte Blattnässedauer wird wieder kürzer.
Keine Sporulation ist möglich, wenn die relative Luftfeuchtigkeit nicht über 85% relativer Luftfeuchtigkeit liegt. Das M.laxa-Modell in fieldclimate.com berechnet schwache und schwere Infektionen. Schwache Infektionen werden für Bedingungen modelliert, unter denen hoch anfällige Stadien wie Blüten und verletzte Früchte kurz vor der Reife infiziert werden. Für die Infektion dieses Gewebes haben sich sehr kurze Blattnässeperioden als ausreichend erwiesen. Mehrere Studien, zum Beispiel:" Phänologische Analyse der Braunfäule der Süßkirsche, verursacht durch Monilinia laxa"(L. Tamm, Chr. E. Minder, and W. Flickiger; 1994) oder "Effects of wounding, fruit age and wetness duration on the development of cherry brown rot in the UK." (X.-M. Xu*, C. Bertone und A. Berrie; 2003) bestätigten, dass für Infektionen nur geringe Nässeperioden erforderlich sind. Im weiteren Verlauf sind unreife Früchte widerstandsfähiger gegen Infektionen, aber kurz vor der Reife werden sie anfälliger und die Bedingungen für schwere Infektionen sind gegeben.
Ausbreitungsmuster im Frühjahr von Monilinia laxa Konidien in Aprikosen-, Pfirsich-, Pflaumen- und Mandelbäumen. Kanadische Zeitschrift für Botanik (1974), 52: 167-176
Modellierung des Feldklimas: Es ist wahrscheinlich, dass die Zeit, die für die Infektion während der Blüte benötigt wird, verkürzt werden muss. Daher verkürzt das Modell die Infektionen im Bereich von 2000 bis 4800 Gradstunden über 5°C.
Westlicher Blütenthrips
Westliche Blütenthripse sind winzige Insekten, etwa 0,03 Zoll lang, mit zwei Paar gefransten Flügeln. Die erwachsenen Tiere haben drei Farbformen, die je nach Jahreszeit unterschiedlich stark ausgeprägt sind. Es gibt eine helle Form, die weiß und gelb ist, abgesehen von leichten braunen Flecken oder Flecken auf der Oberseite des Hinterleibs; eine Zwischenform mit orangefarbenem Thorax und braunem Hinterleib; und eine dunkle Form, die dunkelbraun ist. Die Zwischenform kommt das ganze Jahr über vor, aber im Frühjahr überwiegt die dunkle Form, während die helle Form zu anderen Zeiten des Jahres am häufigsten vorkommt.
Die Nymphen des ersten Stadiums sind undurchsichtig oder hellgelb und färben sich nach der ersten Häutung goldgelb. Das Nymphenstadium dauert zwischen 5 und 20 Tagen.
Schaden
Die Nymphen schlüpfen und ernähren sich in großer Zahl von den Früchten, oft unter dem trocknenden Kelch oder den Blütenteilen. Ihr Fraß hinterlässt Narben auf der Oberfläche der Früchte. Diese Narben vergrößern sich, wenn die Früchte wachsen, und können zu Fruchtdeformationen führen. Thripse können auch kurz vor der Reife der Früchte eine Versilberung verursachen.
Obwohl ein gewisser Fraß an den Blüten stattfindet, entstehen bis zur Fruchtbildung kaum Schäden. Thripse können Endtriebe schädigen und deren Wachstum stoppen. In der Regel bleiben ein bis zwei kleine tote Blätter an der Triebspitze hängen. Knospen wachsen direkt unter dem Endtrieb und geben dem Zweig ein buschiges Aussehen.
Verwaltung
Westliche Blütenthripse überwintern als erwachsene Tiere in Unkräutern, Gräsern, Luzerne und anderen Wirtspflanzen, entweder auf dem Boden von Obstplantagen oder in der Nähe. Im zeitigen Frühjahr, wenn die Überwinterungsplätze gestört werden oder austrocknen, wandern die Thripse zu blühenden Bäumen und Pflanzen und legen ihre Eier in den zarten Teilen der Wirtspflanze ab, z. B. in den Trieben, Knospen und Blütenteilen.
Kulturelle Kontrolle
Thripse werden oft von blühenden Unkräutern auf dem Obstgartenboden angezogen. Um zu verhindern, dass Thripse in die Bäume eindringen, sollten Sie die Deckfrüchte nicht während der Blütezeit der Bäume entfernen. Offene, verunkrautete Flächen in der Nähe von Obstplantagen sollten so früh wie möglich gerodet werden, um die Entwicklung von Thripsen und die Einwanderung erwachsener Tiere in die Obstplantagen zu verhindern.
Organisch akzeptable Methoden
Kulturkontrollen, sauberer Anbau und das Besprühen mit der Entrust-Formulierung von Spinosad sind ökologisch vertretbare Mittel.
Überwachung und Behandlungsentscheidungen
Beginnen Sie mit der Überwachung der Thripse, sobald die Bäume zu blühen beginnen. Untersuchen Sie die Blüten von Bäumen auf Thripse, indem Sie einen Trieb mit fünf bis zehn Blüten gegen eine gelbe Karte klatschen oder nach den unreifen Stadien in den Blüten suchen. Oft lassen sich die Nymphen durch das Klatschen nicht entfernen. Daher sollten Sie auch einzelne Blüten aufschneiden und mit einer Handlinse auf Nymphen untersuchen. Die Nymphen des ersten Larvenstadiums sind weiß und oft schwer zu erkennen, daher sollten Sie genau hinsehen. Kontrollieren Sie mindestens 50 Bäume pro Obstgarten auf Nymphen. In warmen Frühjahren wandern die erwachsenen Tiere oft unbemerkt in einen Block hinein und wieder heraus, so dass es wichtig ist, immer nach Nymphen zu suchen.
Wenn zwei oder mehr erwachsene Thripse vorhanden sind oder Nymphen gefunden werden, ist eine Behandlung angebracht. Wenn eine Behandlung durchgeführt wird, sollte sie erfolgen, bevor sich der Kelch um den sich entwickelnden Fruchtknoten schließt. Wenn Nymphen unter dem Mantel gefunden werden, nachdem dieser sich um die Frucht gelegt hat, sollte Methomyl verwendet werden.
Quelle: Literatur von UC IPM Davis:
- K. R. Day, UC Cooperative Extension, Bezirk Tulare
- K. Tollerup, UC IPM-Programm, Kearney Agricultural Center, Parlier
In FieldClimate verwenden wir folgende Bedingungen für die Berechnung des Risikomodells:
Risikomodell für die Westlicher Blütenthrips Frankliniella occidentalis (Pergande) auf der Grundlage von Tageswerten:
- Das Risiko steigt, wenn die Lufttemperaturen über 18°C und unter 32°C liegen und die relative Luftfeuchtigkeit über 70% bis 20% liegt.
- Das Risiko sinkt, wenn die Lufttemperatur länger als 8 Stunden über 32 °C liegt oder
- Wenn die relative Luftfeuchtigkeit nie höher als 70% ist oder wenn alle Temperaturen unter 15°C liegen, wird das Risiko um 10% reduziert.
Bakterielle Krebserkrankung
Bakterienkrebs ist eine Krankheit der Stämme und Blätter von Prunus, insbesondere von Pflaumen und Kirschen, aber auch von Aprikosen, Pfirsichen und Prunus-Zierpflanzen. Er verursacht eingesunkene Flecken mit abgestorbener Rinde und kleine Löcher in den Blättern.
Die Arten P. syringae existiert als eine große Anzahl von Pathovaren (abgekürzt pv.), die so genannt werden, weil sie zwar alle gleich aussehen, aber unterschiedliche, spezifische Wirte haben. Der Pathovar morsprunorum ist auf Prunus-Arten beschränkt, pv. syringae hat ein viel größeres Wirtsspektrum, aber beide verursachen ähnliche Symptome auf Prunus.
Die Bakterien leben als Oberflächenbewohner (Epiphyten) auf den Blättern und können bei feuchtem Wetter im Frühjahr oder Frühsommer durch die Blattporen (Spaltöffnungen) eindringen und Infektionen in den jungen Blättern hervorrufen. Wenn das Blatt heranreift, dehnen sich diese Infektionen nicht mehr aus und werden als kleine Flecken mit abgestorbenem Gewebe sichtbar. Wenn sich das Blatt vollständig ausdehnt, zieht sich das lebende Gewebe von dem toten Fleck ab, der herausfällt und ein "Einschussloch" hinterlässt.
Schimmelpilze entwickeln sich, wenn die Bakterienzellen durch Wunden oder Blattnarben zum Zeitpunkt des Blattfalls eindringen. Im Sommer, wenn das Gewebe widerstandsfähig ist, und im Herbst und Winter, wenn die Temperaturen niedrig sind, bleibt die Rindenkrankheit mehr oder weniger untätig. Im Frühjahr breiten sich die Infektionen rasch aus und töten die Rinde ab.
Symptome
Die folgenden Symptome können auftreten:
X) An Stämmen und Ausläufern: Im Frühjahr und Frühsommer bilden sich eingesunkene, abgestorbene Rindenstellen, oft begleitet von einem schleimigen Schleim. Breitet sich die Infektion auf den ganzen Zweig aus, stirbt er schnell ab. Es ist jedoch anzumerken, dass die Bildung von Gummi (Gumminose) an der Rinde von Prunus-Arten recht häufig vorkommt, und wenn keine abgestorbene, eingesunkene Rinde vorhanden ist, ist es wahrscheinlich, dass andere Ursachen als Bakterienkrebs vorliegen, z. B. physische Schäden oder Umweltbelastungen.
Auf Blättern: Es entstehen kleine braune Flecken, die oft rund sind und später herausfallen und Löcher hinterlassen - als ob das Blatt von Schrotkugeln getroffen worden wäre, was zu dem volkstümlichen Namen "Schießscharte" führt.
Nicht-chemische Kontrolle
Führen Sie nach Möglichkeit alle Beschneidungen im Juli oder August durch, wenn das Gewebe am widerstandsfähigsten ist. Dies ist auch der beste Zeitpunkt für den Rückschnitt, um das Risiko einer Infektion durch Sporen des Pilzes, der die Silberblattkrankheit verursacht, zu minimieren. Schneiden Sie alle verkrüppelten Stellen aus, indem Sie bis zum gesunden Holz zurückschneiden, und streichen Sie die Wunde umgehend mit einem Wundanstrich, um sie vor Neuinfektionen zu schützen.
Quelle: Königliche Gartenbaugesellschaft
Modell FieldClimate
Risikomodell auf der Grundlage von Tageswerten:
- wenn die Bodenwasserspannung den ganzen Tag über kleiner als 25 ist und der Regen mehr als 2 mm beträgt und die Dauer der Blattnässe länger als 6 Stunden ist, erhöht sich das Risiko um 20%.
- wenn die Bodenwasserspannung ganztägig höher als 40 ist, sinkt das Risiko um 20% oder
- wenn den ganzen Tag keine Blattnässe auftritt, sinkt das Risiko um 10% oder
- wenn keine Stunde mit mehr als 70% rel. Feuchtigkeitsrisiko um 10% abnimmt
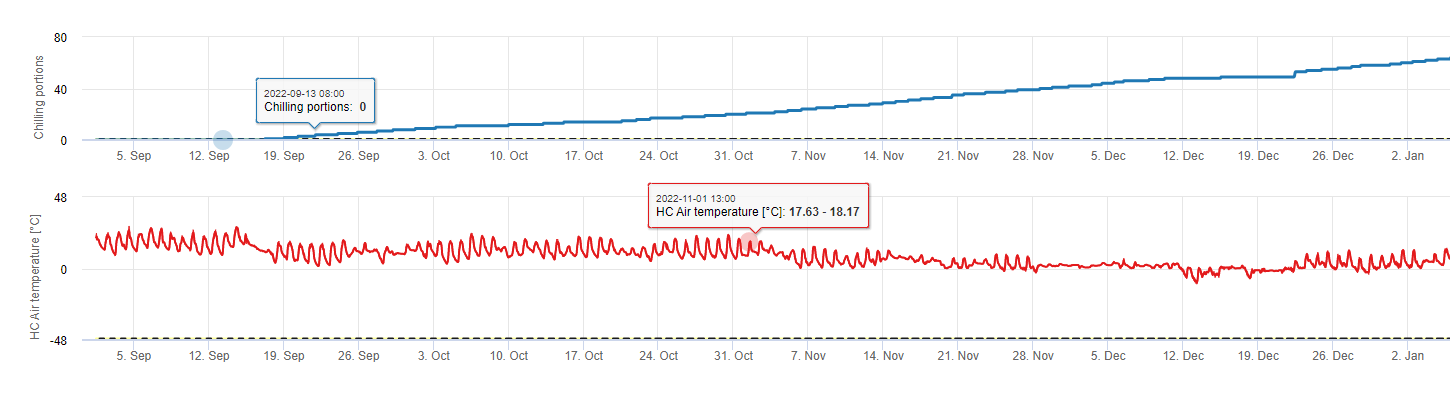
Kühlende Portionen
Kühlung
Steinobstbäume entwickeln ihre vegetativen und fruchttragenden Knospen im Sommer, und wenn der Winter naht, gehen die bereits entwickelten Knospen als Reaktion auf die kürzere Tageslänge und die kühleren Temperaturen in eine Ruhephase über. Diese Ruhephase schützt die Knospen vor der bevorstehenden kalten Witterung. Sobald die Knospen in die Ruhephase eingetreten sind, vertragen sie Temperaturen weit unter dem Gefrierpunkt und wachsen nicht mehr als Reaktion auf Wärmeperioden in der Mitte des Winters. Diese Knospen bleiben in der Ruhephase, bis sie genügend Kälteeinheiten (CU) angesammelt haben. Wenn genug gekühlt akkumuliert, sind die Knospen als Reaktion auf die warmen Temperaturen bereit zu wachsen. Solange genügend CUs vorhanden sind, entwickeln sich die Blüten- und Blattknospen normal. Wenn die Knospen im Winter nicht genügend kühle Temperaturen erhalten um die Keimruhe vollständig aufzuheben, entwickeln die Bäume eines oder mehrere der physiologischen Symptome, die mit einer unzureichenden Abkühlung einhergehen: 1) verzögerte Belaubung, 2) verminderter Fruchtansatz und vermehrte Knospung und 3) verminderte Fruchtqualität.
Unzureichende Abschreckungssymptome
Verspätete Foliation:
Ein klassisches Symptom für unzureichende Abkühlung ist eine verzögerte Belaubung. Ein Baum kann in der Nähe der Stammspitzen ein kleines Blattbüschel haben und 12 bis 20 Zoll unterhalb der Spitzen blattlos sein. Die unteren Knospen brechen mit der Zeit auf, aber die vollständige Belaubung verzögert sich erheblich, der Fruchtansatz ist reduziert und der Baum ist geschwächt. Darüber hinaus verursacht ein starker Austrieb an den unteren Teilen des Baumes Probleme bei der Bewirtschaftung, und die normale Entwicklung der Fruchtknospen des nächsten Jahres kann beeinträchtigt werden.
Reduzierter Fruchtansatz und Knospung:
Die Blüte als Reaktion auf eine unzureichende Abkühlung folgt oft dem Muster, das bei der Blattentwicklung zu beobachten ist. Die Blüte verzögert und verlängert sich, und aufgrund von Anomalien bei der Entwicklung von Stempeln und Pollen ist der Fruchtansatz reduziert. Bei vielen Pfirsichsorten fallen die Blüten vor oder um den Schalenaufbruch ab, aber bei anderen wie 'Jersey Queen' und 'Harvester' bilden sich Knöpfe. Die Knospen entstehen aus Blüten, die zwar scheinbar geblüht haben, sich aber nie zu vollwertigen Früchten entwickeln. Die Früchte bleiben klein und unförmig, während sie reifen. Wenn man diese Früchte aufschneidet, ist der Samen tot. Da die Knöpfchenbildung zu Beginn der Saison nicht sichtbar ist, können die Anbauer die anormalen Früchte nicht ausdünnen, und die sich entwickelnden Knöpfchen dienen als Nahrungsquelle und Überwinterungsort für Insekten und Krankheiten.
Verminderte Fruchtqualität:
Die Auswirkungen unzureichender Kühlung auf die Fruchtqualität werden wahrscheinlich am wenigsten diskutiert, scheinen aber besonders in Zentral- und Südtexas sehr verbreitet zu sein. Die Auswirkungen auf das Blattwachstum und den Fruchtansatz sind dramatisch, aber die Auswirkungen einer unzureichenden Kühlung auf die Fruchtqualität sind subtil und können auftreten, wenn andere Symptome nicht auftreten. Eine unzureichende Kühlung führt bei vielen Sorten zu einer vergrößerten Spitze und einer geringeren Festigkeit. Außerdem kann die Grundfärbung der Früchte grüner als gewöhnlich sein, was möglicherweise darauf zurückzuführen ist, dass die Früchte an Festigkeit verlieren, bevor sich die Grundfarbe vollständig von grün nach gelb verändern kann. Das Ausmaß dieser Qualitätsprobleme hängt von der Sorte und dem Grad des Kältemangels ab.
Modelle
Es gibt verschiedene Modelle zur Berechnung der Kühlung, wobei jedes Modell definiert, was eine Kühleinheit ist. Die drei gebräuchlichsten Modelle sind das Modell der Anzahl der Stunden unter 7°C (45 Grad F), das Modell der Anzahl der Stunden zwischen 2 und 7°C (32 und 45 Grad F) und das Utah-Modell. Die ersten beiden Modelle sind einfach und definieren eine Kühleinheit als eine Stunde unter oder zwischen bestimmten Temperaturen. Die Utah-Methode ist komplexer, da sie das Konzept der relativen Kühleffektivität und der negativen Kühlakkumulation (oder Kältenegation) einführt.
In FieldClimate verwenden wir die Modell zur Berechnung von Kühlanteilen (CP). Die Kühlakkumulationen werden als Kühlaufwand berechnet, wobei ein Temperaturbereich von 2 bis 7°C verwendet wird. Die Berechnung der Chill-Anteile endet nach 96 Stunden bei gleicher oder höherer Temperatur als 15°C (sie liegt zwischen 7 und 15°C).
Die Berechnungen beruhen auf den Arbeiten von Erez A, Fishman S, Linsley- Noakes GC, Allan P (1990) The dynamic model for rest completion in peach buds. Acta Hortic 276: 165-174.
Empfohlene Ausrüstung
Prüfen Sie, welcher Sensorensatz für die Überwachung potenzieller Krankheiten dieser Kultur benötigt wird.