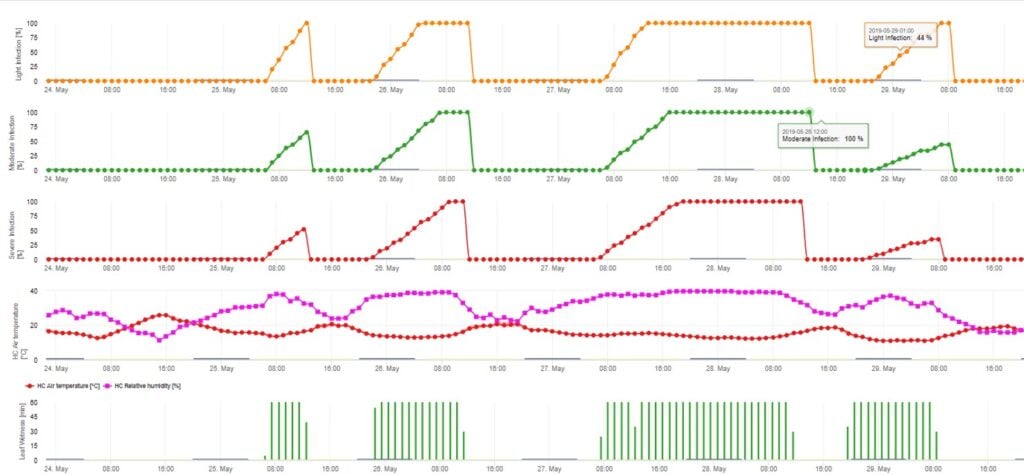
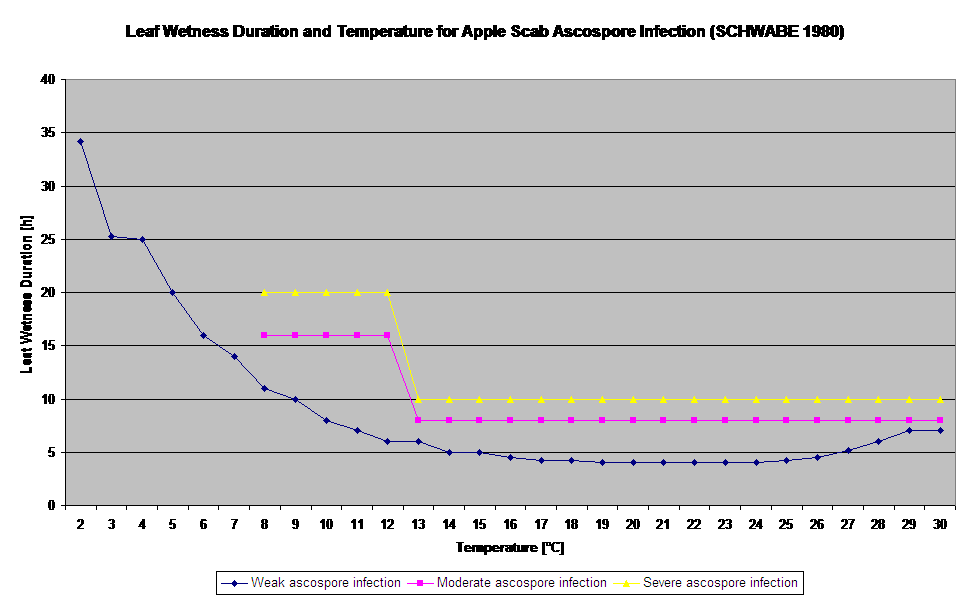
Keimung und Penetration von Ascosporen von Venturia inaequalis auf Blättern oder Früchten hängen von der Blattnässe (nasse Blätter oder rel. Luftfeuchtigkeit >80% (hält das Infektionsniveau bis unter 80% r.h.)) und der Lufttemperatur (über 2°C) ab. Höhere Temperaturen führen unter feuchten Bedingungen zu einer schnelleren Durchdringung, während bei niedrigeren Temperaturen längere Perioden der Blattnässe erforderlich sind. Diese Beziehung wurde erstmals von MILLS und LAPLANTE (1945) veröffentlicht. Unsere Berechnung erfolgt auf der Grundlage der Veröffentlichungen von SCHWABE (1980). SCHWABE beschrieb die Schwere der Schorfinfektionen in Abhängigkeit von der Temperatur. Diese Beobachtungen sind deckungsgleich mit unseren eigenen Beobachtungen.
Das Ascosporen-Infektionsmodell berechnet den Infektionsfortschritt in Abhängigkeit von Blattnässe und Temperatur für schwache, mittlere und schwere Infektionen. Die Einteilung der Vorhersage in drei "Schwereklassen" ist bei Apfelschorfmodellen sehr üblich. Der Infektionsfortschritt wird als Grafik für Stundenwerte dargestellt. Eine Infektion hat bereits stattgefunden, sobald die Verlaufskurve für eine schwache Infektion 100% erreicht. Eine mittelschwere oder schwere Infektion ist abgeschlossen, wenn die Verlaufskurven für diese Schweregrade (100%) erreichen. In der Praxis oder in der Realität kommt es häufig vor, dass eine Infektion am frühen Morgen 70% oder 80% erreicht, und in Abhängigkeit von der aktuellen Wetterlage können Sie erkennen, ob diese Infektion beendet ist oder nicht. Die Anzeige der täglichen Maximalwerte der Infektionsverlaufskurve gibt einen Überblick über die zu erwartenden Infektionstermine in der Saison.
Primäre Verwendung des Modells
Für den Markt ist es absolut wichtig, krankheitsfreie Apfelfrüchte zu haben, um einen guten Preis zu erzielen. Daher neigen die Erzeuger dazu, in ihren Obstgärten überhaupt keine Schorftoleranz zu haben. Das Modell sollte in ein kuratives Anwendungsprogramm integriert werden (Anwendung von Spritzmitteln mit Cyprodinil- oder Pyrimethanilverbindungen in der Vorblütezeit), wenn das Modell auf eine Infektion hinweist (unabhängig davon, ob sie schwach oder stark ist). In der Blütezeit und in der frühen Nachblütezeit sind schwere Infektionen am häufigsten, und alle feuchten Perioden wurden durch Schutzspritzungen abgedeckt. Außerdem wächst das Pflanzengewebe sehr schnell, so dass ein Schutz nur 4 Tage lang wirken kann. In diesem Fall kann auch eine kurative Spritzung nach einer unvollständigen Infektionsperiode erforderlich sein. In diesem Fall kommt es jedoch darauf an, ob es sich um eine schwache oder starke Infektion gehandelt hat und ob die Zahl der infektionsbereiten Ascosporen niedrig oder hoch war.
Literatur
- Schwabe WFS, 1980. Feuchtigkeits- und Temperaturanforderungen für die Infektion von Apfelblättern durch Venturia inaequalis in Südafrika. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Wetterbedingungen, die die Infektion mit Apfelschorf in Südafrika begünstigen. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Biologie, Epidemiologie und Management des Apfelschorfs. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Sekundärinfektionen durch Konidien
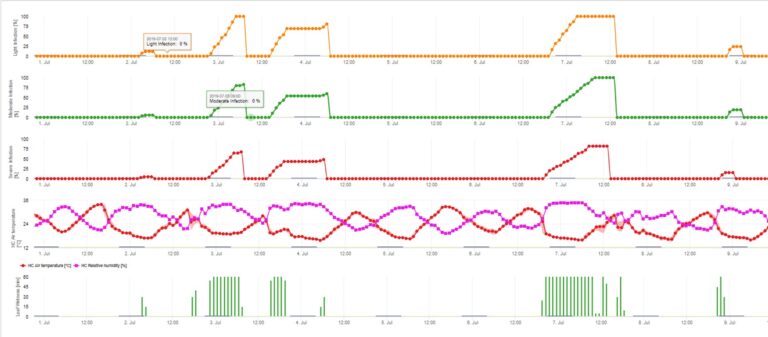
Die Infektion des Apfelschorfs durch Konidien (ungeschlechtlich) hängt von denselben Faktoren ab wie die durch Ascosporen verursachte Infektion. Der Unterschied besteht darin, dass Konidien unter hellen und dunklen Bedingungen ausgeschieden werden können und Konidieninfektionen hauptsächlich im Sommer stattfinden.
Je nach Lufttemperaturregime benötigen die Konidien von Venturia inaequalis längere oder kürzere Blattnässeperioden für die Keimung und das Eindringen in Blätter oder Früchte des Apfelbaums. Diese Beziehung wurde erstmals von MILLS und LAPLANTE (1945) veröffentlicht. Unsere Berechnungen basieren auf den Veröffentlichungen von SCHWABE (1980). Während Mills und LAPLANTE (1945) nur davon ausgingen, dass die Konidien eine bestimmte Zeitspanne der Blattnässe benötigen, bezog SCHWABE (1980) auch die Bedeutung der Temperatur in den Infektionsprozess ein.
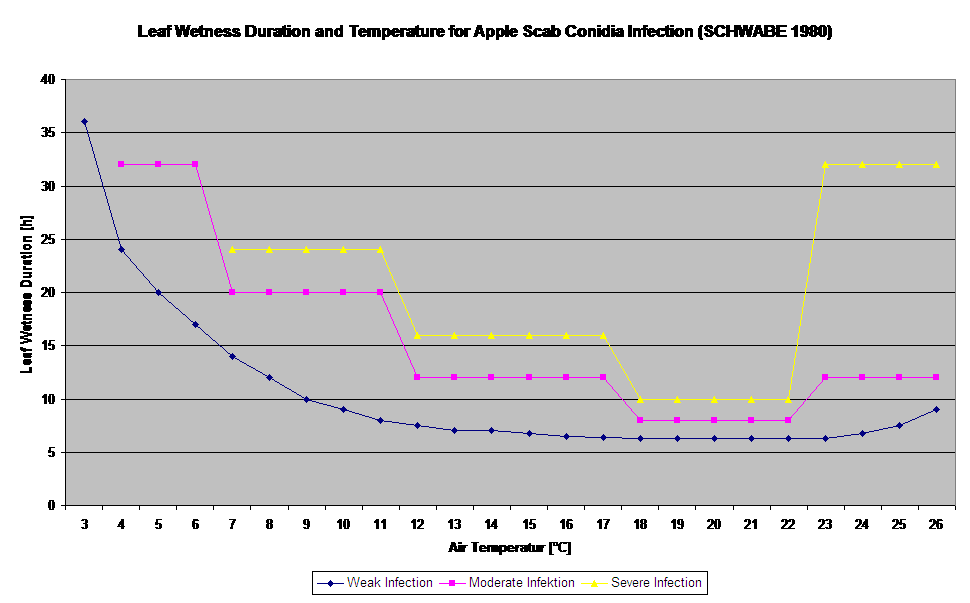
Das Konidien-Infektionsmodell berechnet den Infektionsfortschritt in Abhängigkeit von Blattnässe und Temperatur für schwache, mittlere und schwere Infektionen. Die Einteilung der Vorhersage in "drei Schweregrade" ist für Apfelschorfmodelle sehr üblich. Der Infektionsfortschritt wird als Grafik für Stundenwerte dargestellt. Eine Infektion hat bereits stattgefunden, sobald die Verlaufskurve für eine schwache Infektion 100% erreicht. Eine mittelschwere oder schwere Infektion ist abgeschlossen, wenn die Verlaufskurven für diese Schweregrade den Wert 100% erreichen. In der Praxis oder in der Realität kommt es häufig vor, dass eine Infektion am frühen Morgen 70% oder 80% erreicht, und in Kombination mit dem aktuellen Wetter können Sie erkennen, ob diese Infektion beendet wird oder nicht. Die Anzeige der täglichen Maximalwerte der Infektionsverlaufskurve gibt einen Überblick über die zu erwartenden Infektionstermine in der Saison.
Literatur:
- Infektion mit Venturia inaequalis unter Angabe der Daten der Ascosporen- und Konidieninfektion (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Biologie, Epidemiologie und Management des Apfelschorfs. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Grafische Darstellung des Modells
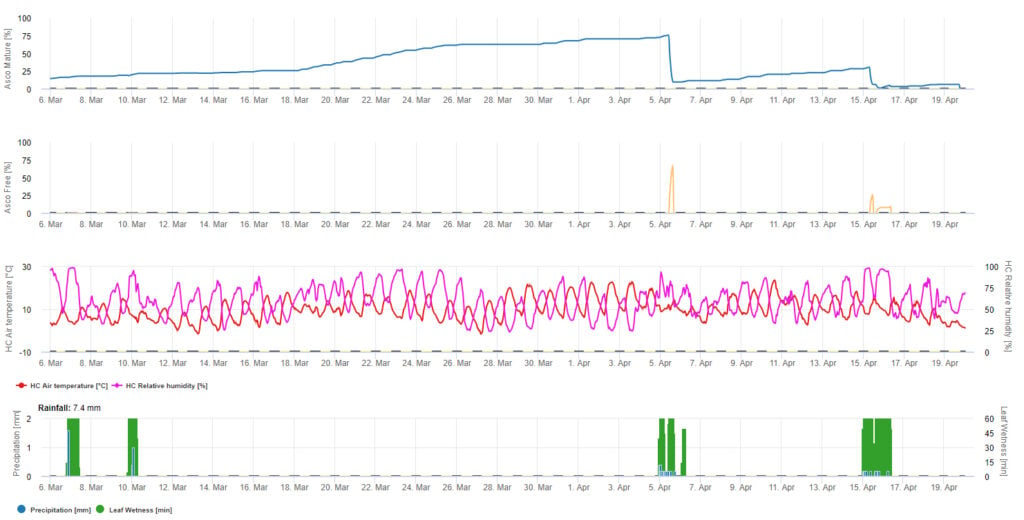
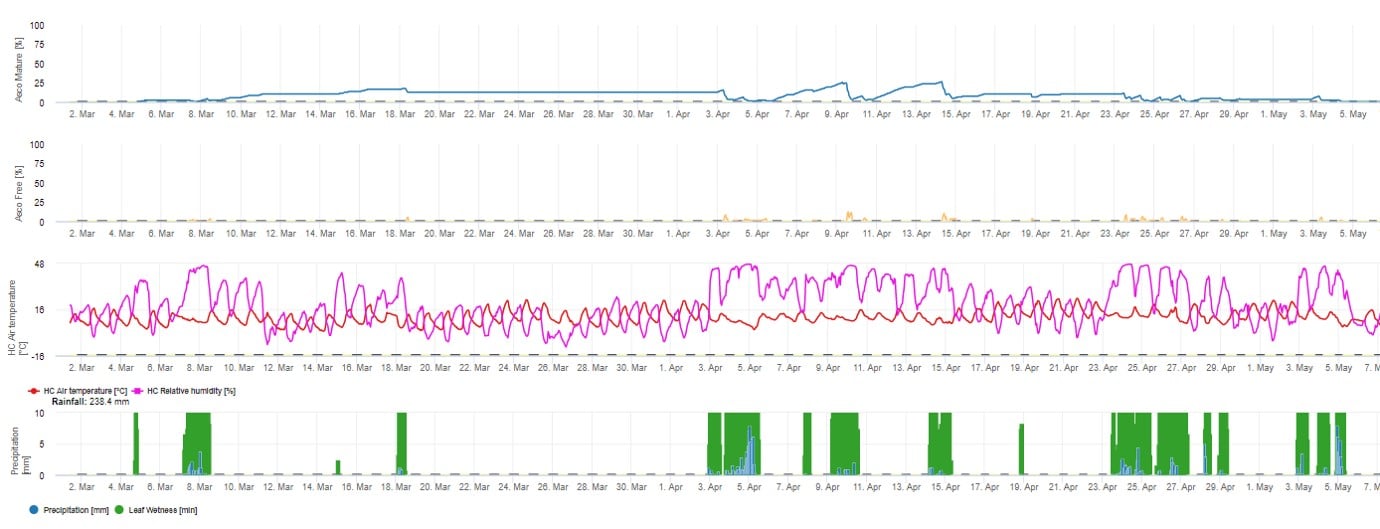
Für die grafische Darstellung des Modells der Ascosporenreifung, -bildung und -abgabe werden Tageswerte verwendet. Es zeigt, dass Blattnässeperioden zu Ascosporenfreisetzungen und Kurven für reife und freigesetzte Ascosporen führen. Der Schweregrad der Freisetzung und die Anzahl der reifen Ascosporen kann nicht in absoluten Werten bewertet werden. Werte zwischen 0 und 100% geben die relative Menge der reifen und freigesetzten Ascosporen in Abhängigkeit von den klimatischen Bedingungen an.
1. Ascosporenreifung, -bildung und -ausstoß:
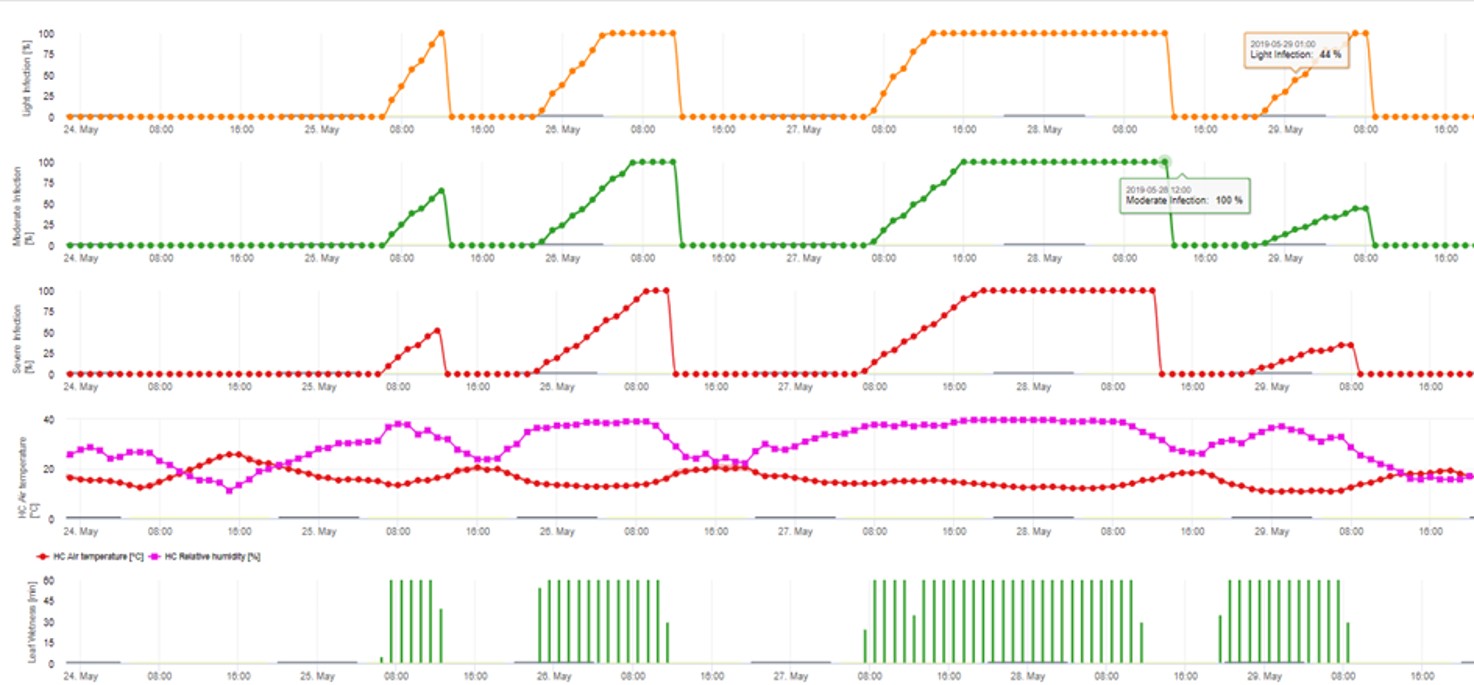
2. Die Ascosporeninfektion wird in drei verschiedenen Schweregraden dargestellt (schwach: orange, mäßig: grün, stark: rot). Wann immer eine 100%-Infektion erreicht wurde, wurden die optimalen Bedingungen für die Infektion des Pflanzengewebes durch die Ascospore ermittelt. Daher müssen Pflanzenschutzmaßnahmen (vor der Infektion prophylaktisch, nach der 100%-Infektion kurativ) berücksichtigt werden:
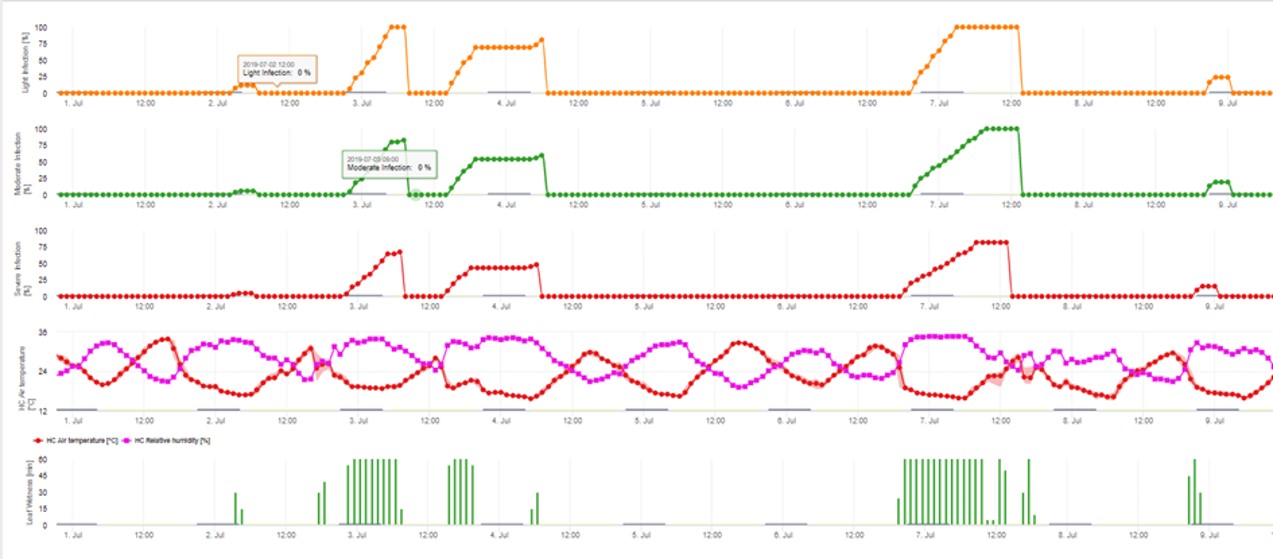
3. Die Konidieninfektion wird in drei verschiedenen Schweregraden dargestellt (schwach: orange, mäßig: grün, stark: rot). Wann immer eine 100%-Infektion erreicht wurde, wurden die optimalen Bedingungen für die Infektion des Pflanzengewebes durch die Ascospore ermittelt. Daher müssen Pflanzenschutzmaßnahmen (vor der Infektion prophylaktisch, nach der 100%-Infektion kurativ) berücksichtigt werden:
Kommerziell angebaute Äpfel müssen frei von Schorf sein. Schorfige Äpfel werden nur noch zur Verarbeitung verkauft. Daher ist es das Ziel aller Pflanzenschutzmaßnahmen sowohl im konventionellen als auch im ökologischen Anbau, schorffreie Früchte zu erhalten. Modelle, die den Ascosporenauswurf und die Ascosporen-/Konidieninfektion bei Äpfeln zeigen, sind sehr wichtige Instrumente, um dieses Ziel zu erreichen.
In konventionellen Anbausystemen werden grundsätzlich zwei Arten von Fungiziden gegen Apfelschorf eingesetzt: a) präventive Produkte wie Captan, Mancozeb, Dithianon und Strobilurine oder b) kurative Produkte wie Cyprodinil (Chorus) oder Pyrimethanil (Scala) oder für die Anwendung in den späteren wärmeren Perioden der Saison die DMI-Fungizide. Tatsächlich verfolgen die meisten Landwirte eine präventive Strategie. Eine praktische vorbeugende Strategie ist jedoch nicht in der Lage, Apfelbäume vollständig zu schützen, da der Apfelbaum wächst und Blüten, Früchte und Blätter entwickelt. Daher schützt die vorbeugende Spritzung nur für einen Zeitraum von 4 bis 7 Tagen in Abhängigkeit vom tatsächlichen Wachstum des Baumes. Solch enge Spritzintervalle sind nicht handhabbar; daher werden die Erzeuger ihre Erfahrungen mit dem lokalen Klima, der Wettervorhersage und den Apfelschorfmodellen in ihr Spritzmanagement einbeziehen. Sie werden die vorbeugenden Spritzungen auf der Grundlage ihrer Erfahrungen und der Wettervorhersage planen. Die Apfelschorf-Infektionsmodelle zeigen ihnen den genauen Zeitpunkt der Infektion (schwach, mäßig und stark) sowie das Ascosporen-/Konidienausscheidungsmodell an, und mit ihrer Erfahrung können sie die Bedeutung einer Infektion einschätzen. Dies gibt ihnen die Möglichkeit, mit einem kurativen Produkt zu handeln, wenn eine Apfelschorfinfektion zu lange nach der letzten vorbeugenden Spritzung aufgetreten ist.
Im ökologischen Apfelanbau hat sich Kalkschwefel als das wirksamste Bekämpfungsmittel gegen Schorf erwiesen. Die optimale Bekämpfung kann erreicht werden, wenn kurz vor der Infektion oder zu Beginn der Infektion gespritzt wird. Dies muss auf der Grundlage der Wettervorhersage geplant werden. Manchmal verpassen wir diesen optimalen Zeitraum und müssen in die feuchten Blätter einer fast vollständigen Schorfinfektion sprühen. Dadurch wird immer noch eine gute Wirksamkeit erzielt. Die Apfelschorfmodelle helfen bei der Entscheidung, ob eine Auflaufspritzung in eine fast vollständige Infektion erforderlich ist.
Symptome des Feuerbrandes
Überwinternde Krebsgeschwüre, die den Erreger des Feuerbrandes beherbergen, sind an Stämmen und großen Ästen oft deutlich als leicht bis tief eingedrückte Stellen mit verfärbter Rinde zu erkennen, die manchmal an den Rändern Risse aufweisen. Die meisten Krebsgeschwüre sind jedoch viel kleiner und nicht so leicht zu erkennen. Sie treten an kleinen Ästen auf, an denen im Vorjahr Blüten- oder Triebinfektionen aufgetreten sind, und oft auch an Schnitten, die zur Entfernung befallener Äste vorgenommen wurden. Da viele dieser Krebsgeschwüre erst später in der Saison auftreten, sind sie oft nicht stark eingedrückt und zeigen selten Rindenrisse an ihren Rändern. Außerdem sind sie oft recht klein, weniger als 2,5 cm lang, mit rötlicher bis violetter Rinde, die mit winzigen schwarzen Pilzfruchtkörpern bedeckt sein kann (vor allem Botryosphaeria obtusa, dem Erreger der Schwarzfäule des Apfels).
Die Symptome der Blütenfäule treten meist innerhalb von ein bis zwei Wochen nach der Blüte auf und betreffen in der Regel die gesamte Blütentraube, die verwelkt und abstirbt und bei Äpfeln braun und bei Birnen ganz schwarz wird. Wenn das Wetter für die Entwicklung des Erregers günstig ist, sind auf den Blüten Kügelchen mit Bakterienschleim zu sehen. Der Sporn, der den Blütenstand trägt, stirbt ebenfalls ab, und die Infektion kann sich auf Teile des tragenden Gliedes ausbreiten und diese abtöten. Die Spitzen junger infizierter Triebe verwelken und bilden ein sehr typisches "Hirtenstab"-Symptom. Ältere Triebe, die nach der Entwicklung von etwa 20 Blättern infiziert werden, zeigen dieses Kräuselsymptom an der Spitze möglicherweise nicht. Wenn sich die Infektion entlang der Triebachse ausbreitet, zeigen die Blätter zunächst dunkle Streifen in den mittleren Adern, welken dann und werden braun, wobei sie während der gesamten Saison fest am Trieb haften bleiben. Wie bei der Blüteninfektion befällt der Erreger oft einen Teil des Astes, der den infizierten Trieb trägt, und tötet ihn ab. Das erste Symptom an Wassertrieben und Trieben, die systemisch von aktiven Krebsen in der Nähe befallen werden, ist die Entwicklung einer gelben bis orangefarbenen Verfärbung der Triebspitze, bevor sie welkt. Außerdem werden die Blattstiele und die mittleren Adern der Grundblätter an solchen Trieben in der Regel vor denen an der Triebspitze nekrotisch.
Je nach Sorte und Entwicklungsstadium zum Zeitpunkt der Infektion kann eine einzige Blüte oder ein einziger Trieb zum Absterben eines ganzen Zweiges führen, und wenn der Haupttrieb oder der Stamm des Baumes befallen ist, kann ein großer Teil des Baumes in nur einer Saison absterben. Im Allgemeinen führen Infektionen jeglicher Art, die zwischen dem Blattfall und dem terminalen Knospenansatz auftreten, zu den größten Verlusten an Ästen und Bäumen. Außerdem neigen stark strukturierte Bäume dazu, weniger schwere Astverluste zu erleiden als solche, die auf schwächere Systeme für hohe Produktivität getrimmt sind. Wenn hochgradig anfällige Apfelunterlagen (M.26, M.9) infiziert werden, bleibt ein Großteil des Edelreises und der Hauptäste oberhalb der Veredelungsstelle in der Regel symptomlos, während sich um die Unterlage herum ein deutlicher dunkelbrauner Krebs entwickelt. Wenn dieser Wurzelstockkrebs den Baum umgürtet, zeigt der obere Teil des Baumes Mitte bis Ende der Saison Symptome eines allgemeinen Rückgangs (schlechte Laubfarbe, schwaches Wachstum). In manchen Fällen verfärbt sich das Laub von Bäumen, die von der Wurzelstockfäule betroffen sind, Ende August bis Anfang September rot, ähnlich wie bei der durch einen bodenbürtigen Pilz verursachten Kragenfäule. Bei einigen Bäumen mit Wurzelstockfäule zeigen sich die Symptome erst im darauffolgenden Frühjahr, wenn sich die Krebsgeschwüre nach oben in den unteren Stamm erstrecken.
Krankheitszyklus
Der bakterielle Erreger des Feuerbrands überwintert fast ausschließlich in den Rissen der im Vorjahr infizierten Äste. Die meisten Krebsgeschwüre und damit auch die wichtigsten Inokulumspender treten an Ästen mit einem Durchmesser von weniger als 38 mm auf, insbesondere an den Schnittflächen, die im Vorjahr zur Entfernung der befallenen Äste angelegt wurden. Im zeitigen Frühjahr beginnen sich die Bakterien an den Krebsrändern als Reaktion auf die wärmeren Temperaturen und die rasche Knospenentwicklung rasch zu vermehren und produzieren einen dicken gelblichen bis weißen Schleim, der sich bis zu mehreren Wochen vor der Blütezeit auf der Rindenoberfläche ausbreitet. Viele Insektenarten (vor allem Fliegen) werden von dem Schleim angezogen und verbreiten die Bakterien in der Folge im gesamten Obstgarten. Sobald die ersten geöffneten Blüten von den Bakterien besiedelt sind, übertragen bestäubende Insekten den Erreger rasch auf andere Blüten und lösen damit eine weitere Blütenfäule aus. Diese kolonisierten Blüten werden innerhalb von Minuten nach einem durch Regen oder starken Tau verursachten Befeuchtungsereignis infiziert, wenn die durchschnittlichen Tagestemperaturen 16 °C oder mehr betragen und die Blütenblätter noch intakt sind (Blütenbehälter und junge Früchte sind nach dem Abfallen der Blütenblätter resistent). Sobald Blüteninfektionen auftreten, kann mit ersten Symptomen gerechnet werden, wenn mindestens 57 Gradtage (DD) über 13 °C erreicht werden, was je nach Tagestemperaturen 5 bis 30 Kalendertage dauern kann.
Mit dem Auftreten von Symptomen der Blütenfäule nehmen Anzahl und Verteilung der Inokulumquellen in der Obstanlage stark zu. Das Inokulum aus diesen Quellen wird durch Wind, Regen und viele zufällige Insektenbesucher an den jungen Triebspitzen weiter verbreitet, was die Wahrscheinlichkeit eines Ausbruchs der Krautfäule erhöht. Jüngste Untersuchungen in Pennsylvania haben ergeben, dass der Fraß von Blattläusen nicht zur Krautfäule beiträgt. Weitere Untersuchungen sind erforderlich, um festzustellen, ob Blattläuse eine Rolle beim Auftreten der Kraut- und Knollenfäule spielen oder nicht. Die meisten Infektionen der Triebspitzen treten zwischen dem Zeitpunkt auf, an dem die Triebe etwa neun bis zehn Blätter haben, und dem Austrieb der Endknospen, wenn Quellen für Inokulum und Insektenvektoren verfügbar sind und die Tagestemperaturen im Durchschnitt 16 °C oder mehr betragen.
In Jahren, in denen keine Blüteninfektionen auftreten, sind die primären Inokulumquellen für die Triebfäulephase die überwinternden Kanker und insbesondere junge Wassertriebe in der Nähe dieser Kanker, die infiziert werden, wenn die Bakterien systemisch von den Kankerrändern in sie eindringen. Solche systemischen Triebinfektionen, die als "canker blight" bezeichnet werden, werden offenbar etwa 111 DD größer als 13 °C nach der grünen Spitze eingeleitet, obwohl sichtbare Symptome möglicherweise erst bei einer Akkumulation von mindestens 157 DD größer als 13 °C nach der grünen Spitze sichtbar werden. In Abwesenheit von Blüteninfektionen ist die Entwicklung von Triebfäuleinfektionen oft auf Bereiche mit überwinternden Krebsen beschränkt.
Modellblütenkrankheit
Das Modell setzt voraus, dass der Anwender spezifische und sich ständig ändernde lokale Ereignisse und Aspekte seiner Obstanlage erkennt, die das Feuerbrandrisiko im Vergleich zu anderen Obstanlagen in der Region erhöhen oder verringern können. Das Modell setzt voraus, dass der Anwender davon ausgeht, dass immer dann ein Feuerbrandrisiko besteht, wenn Blüten an den Bäumen vorhanden sind, insbesondere während des Blütenfalls und der "Nachblüte", wenn an vielen Apfel- und Birnensorten vereinzelte Blüten zurückbleiben können. Der Modellanwender wird gebeten, die Situation an seinem Standort sorgfältig zu bewerten und Bekämpfungsmaßnahmen einzuleiten, wenn Blüten vorhanden sind, die Risikostufen "Hoch" oder "Extrem" sind und eine Benetzung der Blüten in den nächsten 24 Stunden wahrscheinlich ist.
Struktur des Modells: Temperaturen und Nässe: Der wichtigste Feuerbrandprozess, der modelliert werden muss, ist das potenzielle Bakterienwachstum auf den Narben der Apfel- und Birnenblüten. Dieses Wachstum ist temperaturabhängig, so dass eine zuverlässige Vorhersage des Infektionsrisikos eine Messmethode erfordert, die das Wachstum der Bakterien möglichst genau widerspiegelt. Erwinia amylovora Kolonien. Die größte Uneinigkeit unter den Modellierern besteht darin, wie dies geschehen soll.
Das Cougar-Brand-Modell schätzt die bakterielle Wachstumsrate mit Gradstunden auf der Grundlage einer spezifischen Wachstumsratenkurve. Diese Wachstumskurve basiert auf der Wachstumsrate von E. amylovora Bakterien in Labortests. Die Gradstundenwerte werden für jede Stunde des Tages kumuliert, in der die Temperatur über 15 °C liegt. Die Stundenwerte steigen mit dem Temperaturanstieg von 15 °C auf 29 °C, sinken bei höheren Temperaturen und erreichen für jede Stunde mit Temperaturen über 40 °C den Wert Null.
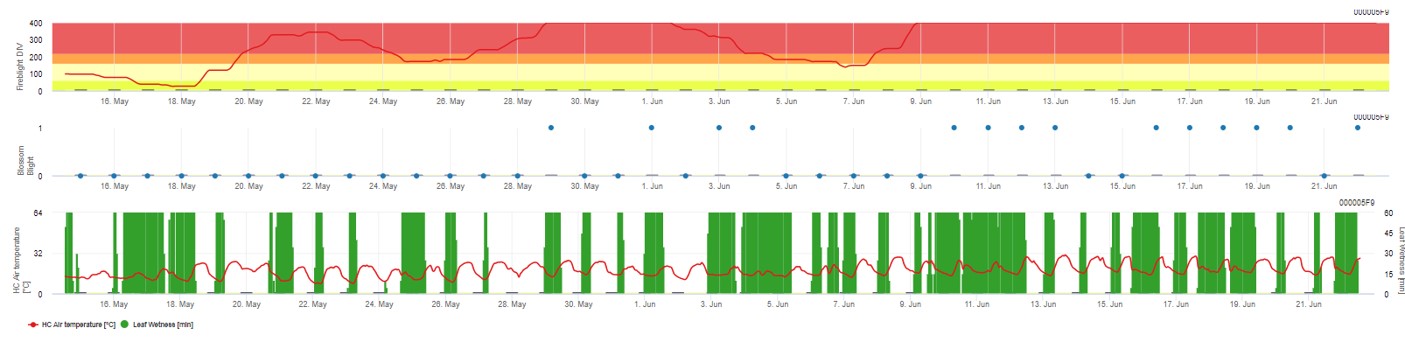
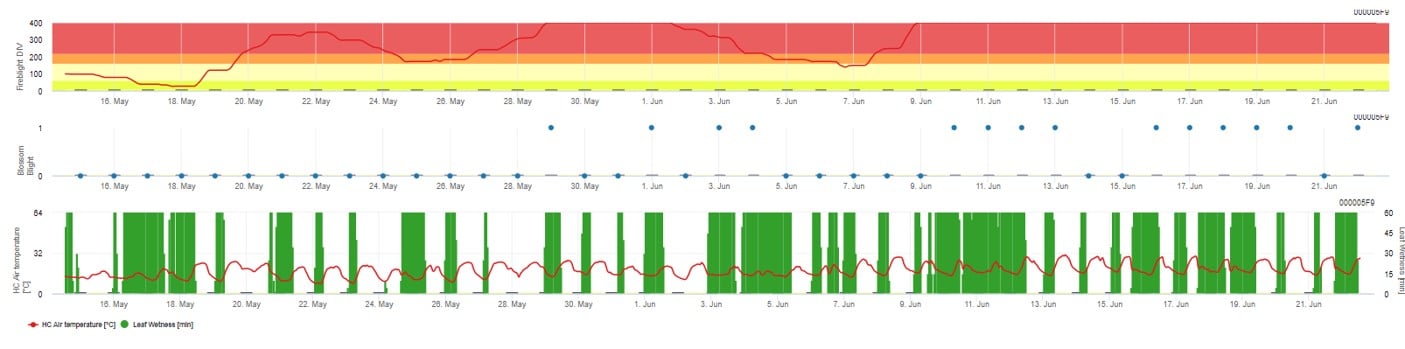
In FieldClimate.com werden beide Modelle in der gleichen Abbildung dargestellt. Das Modell Fireblight DIV basiert auf Courgarblight und das Modell Blossomblight basiert auf Maryblight.
Modell Puma-Fäule
Das Auftreten von Blütenbrand ist einer der sporadischsten Aspekte von Feuerbrand-Epidemien. Das Modell basiert auf der Annahme, dass es eine Fülle von Inokulum gibt und dass vier strenge Bedingungen nacheinander erfüllt sein müssen, damit eine Blüteninfektion auftritt. Diese Bedingungen sind:
1) Die Blüte muss mit intakten Narben und Blütenblättern geöffnet sein, die Narben müssen für die Besiedlung freigelegt sein, Blüten im Blütenblattfall sind resistent;
2) Akkumulation von mindestens 110 °C-Stunden > 18,3 °C innerhalb der letzten 44 °C-Tage > 4,4 °C definiert das epiphytische Infektionspotenzial für die älteste offene und damit am stärksten kolonisierte Blüte im Obstgarten
3) ein Benetzungsereignis in Form von Tau oder 0,2 mm Regen oder 2,5 mm Regen am Vortag ermöglicht die Bewegung von Bakterien von den besiedelten Narben zu den Nektarthoden
4) eine durchschnittliche Tagestemperatur von >= 15,6°C: Dies kann sich auf die Geschwindigkeit auswirken, mit der die Bakterien in die Nektarthoden einwandern, sowie auf die Vermehrung der Bakterien, die für eine Infektion erforderlich ist.
Grafische Darstellung der Modelle
Beide Modelle werden auf der gleichen Seite angezeigt. Die grafische Darstellung der Feuerbrandmodelle ist am besten geeignet, wenn die Werte auf Tagesbasis aggregiert werden. Die Grafik zeigt die Klimawerte im Zusammenhang mit dieser Krankheit. Die Cougar Blight-Ergebnisse heißen Fire Blight DIV und die Blossom Blight-Ergebnisse. Zur Interpretation der Cougar Blight Ergebnisse ist die Grafik mit 5 verschiedenen Farben unterlegt. Die Verteilung dieser Farben erfolgt auf der Grundlage der Einstellungen in der ersten Krautfäule-Historie der Obstanlage (siehe Schaltfläche rechts oben). Die 5 Farben geben die Risikoklasse für die DIV-Werte an.
Interpretation der Werte (in Abhängigkeit vom Feuerbrandvorkommen der letzten Jahre, Grundeinstellung):
Aktive Krebsgeschwüre in der Nähe vorhanden
Legen Sie die Einstellungen für die Feuerbrandhistorie mit dem blauen Boden "Einstellungen" auf der rechten oberen Seite fest:
Anzeige der Ergebnisse des Feuerbrandmodells durch 1) Modell Cougar Blight (aufgrund der Historie und der Hintergrundfarben als Fireblight DIV bezeichnet) und 2) Modell Blossom Blight
| Potenzial für das Vorhandensein von Krankheitserregern | Niedrig | Mäßig | Hoch | Extrem |
| Potenzial für das Vorhandensein von Krankheitserregern | Niedrig | Mäßig | Hoch | Extrem |
| Kein Feuerbrand in den letzten zwei Jahren | 0-350 | 350-500 | 500-800 | 800+ |
| Feuerbrand in der Region zwei Saisons | 0-300 | 300-500 | 500-750 | 750+ |
| Feuerbrand in der Region zwei Saisons | 0-250 | 250-450 | 450-700 | 700+ |
| Feuerbrand im Obstgarten im letzten Jahr | 0-200 | 200-350 | 350-500 | 500+ |
Aktionsschwelle:
Eine Bekämpfung wird empfohlen, wenn ein hohes oder extremes Infektionsrisiko festgestellt wird. Faktoren, die den Schaden bestimmen, sind: Anzahl der Blüten, Alter des Baumes, Wuchsstärke und Sorte,... zu.
Die Wahrscheinlichkeit einer Blossom Blight-Infektion wird durch einen Balken von 0 bis 1 angezeigt (d. h. alle Bedingungen sind erfüllt oder nicht).
Ziel der Feuerbrandmodelle ist es, die Wahrscheinlichkeit einer Infektion durch Erwinia amyloflora im Obstgarten.
Das Mary Blight-Modell, das für die Blütenfäule ausgewertet wird, zeigt sehr gut Infektionssituationen mit hohen wirtschaftlichen Auswirkungen an. Aus diesem Grund wird es häufig verwendet, um den Einsatz von Antibiotika gegen diesen Krankheitserreger anzuzeigen. Cougar Blight gibt Auskunft über das Risiko einer Feuerbrandinfektion aufgrund der allgemeinen Ausbreitungsmöglichkeiten des Erregers. Seine Gewichtung durch die Historie einer Obstanlage ist sehr hilfreich, um uns anzuzeigen, wie sorgfältig wir die Obstanlagen auf Feuerbrandsymptome kontrollieren müssen, selbst in Situationen, in denen Mary Blight keine Infektion anzeigt.
Biologie des Apfelwicklers
Die C. pomonella überwintert als reife Larve eingesponnen in einem Kokon, der sich an geschützten Stellen des Baumes (Rindenspalten oder -risse), die bevorzugt werden, oder am Fuß des Baumes in der Bodenstreu befindet. In modernen, dicht bepflanzten Apfelplantagen gibt es wegen der glatten Rinde nur sehr wenige Kokonstellen an den Bäumen. Die Puppen bilden sich in dem von der reifen Larve gesponnenen Kokon.
Das adulte Stadium schlüpft aus der Puppenhülle und fliegt nach einer kurzen Zeit der Aushärtung des Exoskeletts in den Baum. Aus der Literatur geht hervor, dass die Mottenaktivität im oberen Drittel der Baumkronen stattfindet. Die erwachsenen Tiere sind in der Dämmerung für einige Stunden aktiv und haben eine durchschnittliche Aktivitätsschwelle von 13 bis 15 °C. Die Paarung findet in der Regel auf dem Wirt statt, und die Eiablage erfolgt unmittelbar danach. An warmen Abenden ist es möglich, dass ein weiblicher Falter innerhalb von zwei Tagen schlüpft, sich paart und Eier legt. A C. pomonella Die Motte kann unter idealen Bedingungen über einen Zeitraum von 20 Tagen überleben, aber auf dem Feld ist es wahrscheinlich, dass sie viel kürzer überlebt, besonders im heißen Sommer. Unter Laborbedingungen legt ein Mottenweibchen bis zu 100 Eier, aber die tatsächliche Fruchtbarkeit im Feld ist wahrscheinlich geringer und die meisten werden in den ersten Tagen nach dem Auftauchen und der Paarung gelegt.
Die Eier werden einzeln auf der Blattoberseite, auf Zweigen oder auf Früchten abgelegt. In der ersten Generation werden die Eier nur auf Blättern abgelegt, bis etwa zwei Wochen nach dem Fruchtansatz die Unschärfe der Früchte nachlässt. In der zweiten Generation werden etwa 65% der Eier auf Blättern abgelegt, die sich jedoch in der Regel in unmittelbarer Nähe einer Frucht befinden. Die Eier sind bei der ersten Ablage cremeweiß, haben in der Mitte der Entwicklung einen roten Ring und kurz vor dem Schlupf ist die schwarze Kopfkapsel zu sehen.
Eine frisch geschlüpfte Larve wandert umher, bis sie eine Frucht gefunden hat. Sie beißt sich durch die Schale und ernährt sich einige Tage lang vom Apfelfleisch unter der Schale. Beim Durchbeißen der Schale und beim Fressen unter der Schale stößt die Larve den Kot aus dem Eintrittsbereich heraus. Um die Eintrittsstelle herum bildet sich oft ein roter Ring, vor allem bei jungen Früchten. Nachdem sich die Larve in das nächste Stadium gehäutet hat, bohrt sie sich in den Kern, wo sie sich bis zur Reife von den Samen ernährt. Wenn die Larve fast reif ist, gräbt sie einen Tunnel vom Kerngehäuse zur Außenseite des Apfels und verstopft das Loch mit Kot und Seide. Wenn die Larve bereit ist, einen Kokon zu spinnen, verlässt sie die Frucht und sucht sich einen geschützten Platz, indem sie über den Baum wandert oder sich an einem Seidenfaden auf den Boden fallen lässt.
Modell für Apfelwickler
Unser Modell basiert auf der Akkumulation der effektiven Temperaturen zwischen >= 10°C und <=32°C. In der nördlichen Hemisphäre beginnt diese Akkumulation mit dem ersten Januar. Auf der Südhalbkugel beginnt sie mit dem ersten Juli. Wir akkumulieren jede Messung in ihrem Äquivalent für den Tag (siehe Grafik unten). Die folgenden Schwellenwerte werden für die Entwicklung der Insekten verwendet:
*Vollständige Erzeugung= 650
| *Eierlegen bis zum Schlüpfen der Eier | = 88 |
| *Erstes Larvenstadium | = 60 |
| *Zweites Larvenstadium | = 65 |
| *Drittes Larvenstadium | = 55 |
| *Vorläufiges Larvenstadium | = 45 |
| *Fünftes Larvenstadium plus Puppen | = 335 |
Das vom Modell erstellte Diagramm zeigt das erste Auftreten und die Dauer der Larvenstadien 1 bis 5 und die Anwesenheit von Erwachsenen in 5 Stufen.
| *Stufe 1: | Flugaktivität, Nicht-Eierlegen |
| *Stufe 2: | Flugaktivität mit möglicher Eiablage. |
| *Stufe 3: | Flugaktivität mit gelegentlicher Eiablage. |
| *Stufe 4: | Flugaktivität mit mäßiger bis starker Eiablage. |
| *Stufe 5: | Flugaktivität mit starker Eiablage. |
Darstellung der adulten Stadien des Apfelwicklers in 5 verschiedenen Stufen (erstes Diagramm) sowie die akkumulierten Gradtage (rote Linie, zweites Diagramm) für die adulte Entwicklung und die blau dargestellten Larvenstadien (drittes Diagramm).
In Apfelplantagen kommen mehrere Blattlausarten vor. Die Grüne Apfelblattlaus, die Haferapfelblattlaus oder Apfelgrasblattlaus und die Rosige Apfelblattlaus. In den privaten Gärten hat vor allem die Rosige Apfelblattlaus eine gewisse Bedeutung. Diese Blattlaus führt bei Befall zu starken hormonellen Veränderungen an den Triebspitzen. Die Blätter rollen sich ein und der Trieb wird verkürzt. Später werden die Blätter gelb und sterben ab. Diese Blattlaus befällt vor allem junge Triebe. Die Rosige Apfelblattlaus kann schon bei geringem Befall schwere Schäden verursachen. Dagegen können die Grüne Apfelblattlaus und die Apfelgraslaus in größeren Mengen auftreten, ohne zu wirtschaftlichen Schäden zu führen. Die Apfelgraslaus ist überhaupt als nicht wirtschaftlich bedeutend einzustufen.
Rosarote Apfelblattlaus
Es handelt sich um eine alte europäische Art, die um 1870 in die Vereinigten Staaten eingeführt wurde. Gegen Ende des 19. Jahrhunderts wurde sie zu einem bedeutenden Schädling an Äpfeln. Damit diese Art gedeihen kann, muss ihre Sommerwirtspflanze, der Schmalblättrige Wegerich, im Überfluss vorhanden sein. Zwischen der Einschleppung und Ausbreitung der Rosa Apfelblattlaus und dem Schmalblättrigen Wegerich besteht eine bemerkenswerte Parallelität. Diese Blattlaus kommt weltweit in allen Obstanbaugebieten vor.
Lebensphasen
Ei: Das Ei ist oval und an der Seite neben der Rinde leicht abgeflacht. Die Länge schwankt zwischen 0,49 und 0,56 mm. Bei der ersten Eiablage ist es leuchtend gelb und mit einer klebrigen Substanz bedeckt, die mit der Zeit aushärtet. Die Farbe ändert sich allmählich in grünlich-gelb und schließlich in ein glänzendes Tiefschwarz. Die Zeit, die für diesen Farbwechsel benötigt wird, schwankt unter normalen Außenbedingungen zwischen etwa neun Tagen und mehr als zwei Wochen.
Nymphe: Die aus den Eiern schlüpfenden Tiere sind allesamt lebendgebärende, flügellose Weibchen. Es gibt fünf Stadien. Das letzte ist die reife Stammmutter, die kurz nach dem vierten Instadium beginnt, lebende Jungtiere parthenogenetisch zu produzieren. Sie bringt durchschnittlich fünf bis sechs Junge pro Tag zur Welt. Die Nymphen der zweiten Generation, bei denen es sich ausschließlich um Weibchen handelt, erreichen nach zwei bis drei Wochen die Geschlechtsreife. Die meisten Nymphen beginnen mit der Fortpflanzung auf dem Apfel, aber einige wenige entwickeln Flügel und wandern zum Wegerich.
Die dritte Generation wird im Juni und Anfang Juli produziert. Die meisten dieser Generation entwickeln Flügel und wandern zum Schmalblättrigen Wegerich. In manchen Jahreszeiten erzeugen die flügellosen Weibchen der dritten Generation eine vierte Generation auf Apfel. In den letzten Jahren wurde in einigen Gebieten beobachtet, dass schädliche Populationen der Rosa Apfelblattlaus in Obstanlagen bis zum Hochsommer oder noch später fortbestehen. Es ist nicht bekannt, ob diese Veränderung auf den Selektionsprozess, die Veränderung der Lebensweise oder den fehlenden Bedarf an einem alternativen Wirt zurückzuführen ist.
Erwachsene: Die erwachsenen Tiere unterscheiden sich erheblich in der Farbgebung. Die allgemeine Farbe ist rosabraun, mit einem rosafarbenen Schimmer aufgrund eines pudrigen Belags. Einige der älteren Exemplare sind violett, während die jüngeren Exemplare deutlich rötlich-rosa sind.
Host-Bereich
Der Apfel ist der bevorzugte Wirt, aber die Blattlaus ernährt sich auch von Birne und Weißdorn. Cortland, Golden Delicious, Rhode Island Greening und Ida Red sind besonders anfällige Apfelsorten. Die Blattlaus ist in allen Obstanbaugebieten der Vereinigten Staaten und Kanadas zu finden.
Verletzung oder Beschädigung: Der Fraß der Rosinenblattlaus führt häufig dazu, dass sich die Apfelblätter einrollen, beginnend mit dem Fall der Blütenblätter. Diese Blätter können sich später leuchtend rot verfärben. Der Fraß an den Blättern um die Fruchtstände herum führt oft zu einer Bündelung, Verkümmerung und Missbildung der sich entwickelnden Früchte. Diese Anomalien verschlimmern sich im Laufe der Fruchtentwicklung und können die Früchte schließlich unverkäuflich machen.
Große Blattlauspopulationen können große Mengen an Honigtau als Abfallprodukt des Saftes, von dem sie sich ernähren, produzieren. Der auf den Früchten ausgeschiedene Honigtau dient als Nährboden für Rußtaupilze, die die Oberfläche des Apfels angreifen. Die Giftstoffe im Speichel der Blattläuse dienen auch als "Stopptropfen" und verhindern, dass die Früchte bei der normalen Ernte abfallen (natürliche Trennung vom Baum).
Lebensgeschichte: Die Blattlaus überwintert im Ei-Stadium. Der Schlupf erfolgt zu Beginn des Frühjahrs, etwa eine Woche bis 10 Tage später als bei der Apfelkornblattlaus und etwa zur gleichen Zeit wie bei der Apfelblattlaus. Die Eier schlüpfen, wenn sich die Knospen im Frühjahr öffnen, und zwar über einen Zeitraum von zwei Wochen. Sobald sie schlüpfen, suchen die Jungtiere die sich öffnenden Knospen des Apfels auf; sie scheinen die Fruchtknospen zu bevorzugen. Sie ernähren sich von der Außenseite der Blattknospen und Fruchtknospen, bis sich die Blätter zu entfalten beginnen. Dann arbeiten sie sich ins Innere der Knospen vor und beginnen, den Saft aus den Stängeln und neu gebildeten Früchten zu saugen.
Durch ihren Fraß rollen sich die Blätter ein, was die Blattläuse vor Spritzmitteln und einigen Feinden schützt. Die starke Kräuselung der Blätter, die diese Art verursacht, ist wahrscheinlich das charakteristischste Merkmal ihrer Arbeit. Eine einzige Stängelmutter, die sich auf der Unterseite eines Blattes in der Nähe der Mittelrippe befindet, bewirkt, dass sich das Blatt stark einrollt. Es braucht nur wenige Stängelmütter, um alle Blätter rund um eine sich öffnende Blütenknospe stark zu kräuseln, was einen idealen Schutz für die sich schnell entwickelnden Blattläuse darstellt. Die Stängelmütter erreichen ihre Reife, wenn die Apfelbäume zu blühen beginnen.
Die reifen Stammmütter sind sehr inaktiv. Sie lassen sich nieder, ernähren sich und bringen schnell Junge zur Welt. Wenn sie sich verteilen, entfernen sie ihre Schnäbel schnell aus dem Pflanzengewebe und suchen sich eine andere Stelle, an der sie weitermachen können. Die Stängelmütter sind etwa zwei Wochen nach dem Schlüpfen reif. Wie lange das dauert, hängt weitgehend von den Witterungsverhältnissen ab. Die Produktion von Jungtieren beginnt in der Regel zwei oder drei Tage nach der letzten Häutung und dauert ohne Unterbrechung mehr als einen Monat lang an.
Die Gesamtproduktion eines einzelnen Weibchens liegt bei durchschnittlich 185. Normalerweise erstreckt sich die Fortpflanzungszeit von etwa Anfang Mai bis Juni. Das Maximum der Vermehrungsaktivität liegt in der Regel in der letzten Maiwoche und der ersten Juniwoche, wenn die jungen Früchte beginnen, sich zu setzen und aktiv zu wachsen. Die Rosige Apfelblattlaus befällt selten die jungen und schnell wachsenden Triebe. Sie beschränkt sich auf das Laub, die Blütenstängel und die jungen Früchte.
Eines der charakteristischen Merkmale dieser Art ist die Ansammlung der Jungtiere um die Mutter. Jedes einzelne Muttertier oder jede Gruppe von Muttertieren hat Hunderte von Jungtieren um sich geschart. Die befallenen Blätter können bald von mehr als einer Schicht von Blattläusen bedeckt sein. Diese Angewohnheit, sich zu versammeln, tötet die befallenen Blätter bald ab und bewirkt die erzwungene Abwanderung der Blattläuse. Die Jungtiere bewegen sich aktiv und eilig, scheinbar in dem Bestreben, einen geeigneten Futterplatz zu finden. In dieser Zeit findet man sie häufig auf den sich bildenden Früchten oder an den neuen, saftigen, sich entfaltenden Blättern.
Die zweite Generation benötigt vier bis 40 Tage, um die Geschlechtsreife zu erreichen und Junge zu produzieren. Die Mehrheit der zweiten Generation sind flügellose Weibchen. Die durchschnittliche Gesamtproduktion eines jeden Individuums in der zweiten Generation beträgt etwa 119. Die Gewohnheiten und Aktivitäten der dritten Generation unterscheiden sich nicht von denen der zweiten Generation. Die Blattläuse sammeln sich in großer Zahl auf der Unterseite des Laubes und verursachen starke Kräuselungen. Sie befallen auch die ansetzenden und sich entwickelnden Früchte, was zu charakteristischen Verletzungen führt.
Der Großteil dieser dritten Generation bekommt Flügel und wandert zum Schmalblättrigen Wegerich, der Sommerwirtspflanze. Es kann eine vierte Generation gebildet werden. Diese bekommen alle Flügel und wandern zum Schmalblättrigen Wegerich ab. Nach der letzten Häutung sind die geflügelten erwachsenen Tiere sehr zart und inaktiv. Sie bleiben zwei oder drei Tage lang in den eingerollten Blättern, bevor sie sich auf ihren Flug begeben. Unmittelbar vor dem Flug werden sie sehr aktiv und nervös, rennen umher oder bewegen ihre Flügel auf und ab in Erwartung ihres Fluges.
Grüne Apfelblattlaus
Die Grüne Apfelblattlaus, Aphis pomi (DeGeer), ist in allen Apfelanbaugebieten weit verbreitet. In der nördlichen Hemisphäre tritt sie in Apfelplantagen erstmals Ende Mai bis Anfang Juni auf. Das Insekt saugt den Saft aus den Blättern von Wassertrieben und sukkulenten Endblättern. Die Grüne Apfelblattlaus findet man in der Regel in der Nähe der Hauptadern auf der Unterseite des Blattes. Die Grüne Apfelblattlaus befällt auch Birne, Weißdorn, Quitte, Zierapfel und Spiraea.
Beschreibung der Grünen Apfelblattlaus
Die Eier der Grünen Apfelblattlaus sind oval und glänzend schwarz. Die Nymphen und erwachsenen Tiere sind dunkel- oder hellgrün. Die Grüne Apfelblattlaus hat einen grünen Kopf, einen gleichmäßig grünen oder gelblich-grünen Hinterleib, schwarze Beine und lange schwarze Rispen - die "Schwanzröhren" (am hinteren Ende des Insekts). Die erwachsenen Weibchen sind dunkelgrün bis schwarz und geflügelt. Während des Sommers bringen diese Weibchen (Alate) parthenogenetisch lebende Jungtiere zur Welt.
Biologie der Grünen Apfelblattlaus
Die Grüne Apfelblattlaus überwintert als Ei an Ablegern, an der Basis von Knospen und an Blattnarben von Endtrieben. Die Eier schlüpfen normalerweise, wenn die Knospen aufplatzen und sich die ersten Blätter entfalten. Die Nymphen beginnen sofort, sich von den sich entwickelnden Blättern zu ernähren. Sie halten sich zunächst am Endtrieb auf und wandern später zu den älteren Blättern der Traube. Ausgewachsene Blattlausweibchen produzieren innerhalb von zwei Wochen flügellose Weibchen, wobei jedes Weibchen 50-100 lebende Nachkommen hervorbringen kann. Die jungen Blattläuse entwickeln sich in sieben bis zehn Tagen. Die Populationen der Grünen Apfelblattlaus bauen sich auf Äpfeln in der Regel im zeitigen Frühjahr (Blüte, Blüte, Herbst) langsam auf und nehmen mit steigenden Tagesdurchschnittstemperaturen zu. Geflügelte Blattläuse und flügellose Blattläuse werden im Sommer produziert. Grüne Apfelblattläuse sind im Juli und Anfang August am zahlreichsten. In der Anfangsphase eines Befalls gibt es mehr Nymphen als erwachsene Alate. Mit zunehmender Population steigt der Anteil der Alate. Die Kolonien der Grünen Apfelblattlaus können sich unter günstigen Bedingungen innerhalb einer Woche verdoppeln. Wenn die Blattläuse kein saftiges neues Wachstum haben und sich von älteren Blättern ernähren müssen, kann die Zahl der produzierten Jungtiere um bis zu 50% sinken. Bei Tagestemperaturen von 30-32 °C und mehr vermehren sich die Weibchen nicht gut. Wenn die Temperaturen mehrere Tage lang hoch sind, sterben die Blattläuse ab. Im Laufe des Sommers entwickeln sich mehrere Generationen der Grünen Apfelblattlaus, und die geflügelten Formen verbreiten sich in der gesamten Obstanlage. Manchmal bilden die Grüne Apfelblattlaus und die Rosige Apfelblattlaus (Dysaphis plantaginea Pars.) Kolonien auf ein und demselben Blatt. Die rosafarbene Apfelblattlaus ist rosa oder violett. Im Herbst entwickeln sich die flügellosen Männchen und Weibchen und paaren sich, und die Weibchen legen Eier für den Winter ab. Die größte Anzahl von Eiern befindet sich 15-20 cm von den Zweigspitzen entfernt. Die Eier befinden sich nur selten an den großen Gerüstästen oder an den Stämmen der Apfelbäume.
Schaden
Die Grüne Apfelblattlaus saugt den Saft aus dem Blatt. Ein starker Befall verringert die Vitalität und das Wachstum der Triebe. Dies ist vor allem in Baumschulen und jungen, nicht tragenden Obstgärten von Bedeutung. Das Insekt kann die Größe der Knospen und die Länge der Internodien verringern und eine Blattrollung verursachen. Der Blattlausfraß kann auch das Wachstum von Seitenästen anregen, was die Baumform beeinträchtigen kann. Gekräuselte Blätter und geschwächte Enden sind anfällig für Winterschäden. Der von den Blattläusen produzierte Honigtau kann auf die Früchte tropfen, wodurch Rußpilze wachsen können. Der Pilz verunstaltet die Früchte und mindert den Marktwert. Bei starkem Befall kann sich die Grüne Apfelblattlaus von unreifen Äpfeln ernähren und Berostung verursachen.
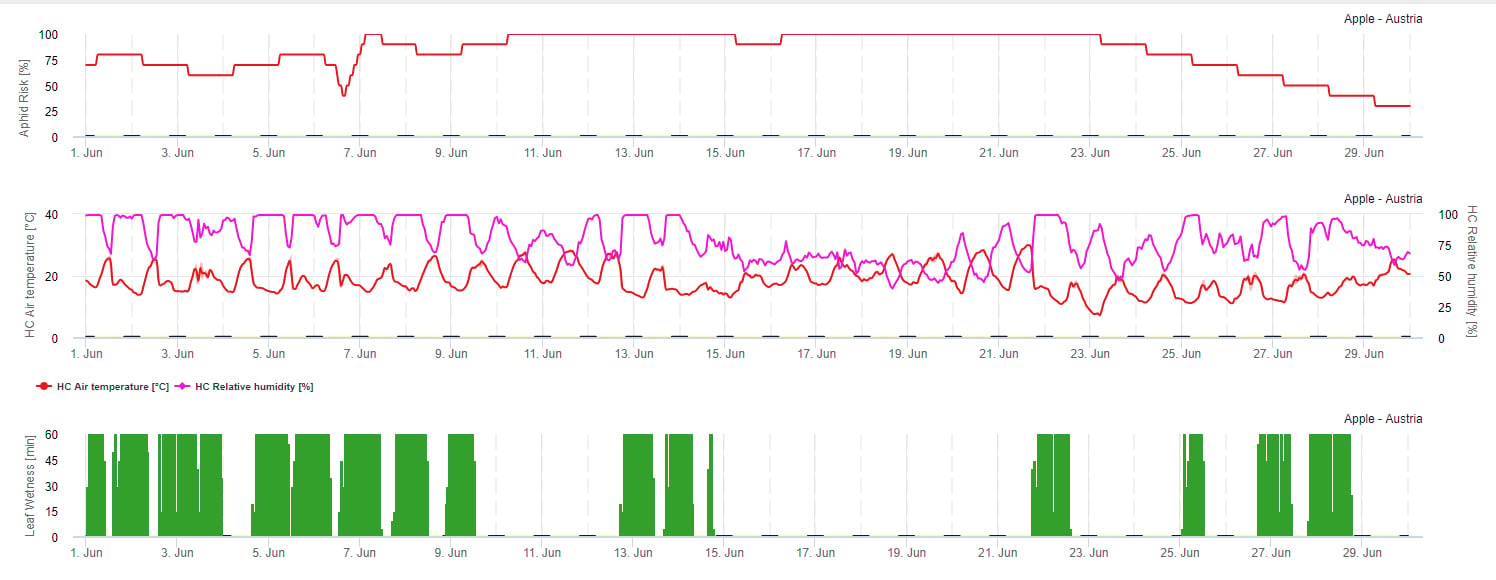
Modell des Ausbreitungsrisikos
Die Vermehrung von Blattläusen wird hauptsächlich durch die Temperatur beeinflusst. Je mehr Stunden in der Nähe des Temperaturoptimums (20-32 °C) liegen und je mehr Nahrungsquellen zur Verfügung stehen, desto höher ist die Vermehrungsrate. Die Eier und die jungen Stadien sind dagegen empfindlich gegenüber trockener Luft. Auch eine niedrige relative Luftfeuchtigkeit kann den Blattlauspopulationen schaden (unter 30% r.h.). Neben Insektiziden werden Blattläuse auch durch Raubtiere und parasitische Pilze bekämpft. Die parasitären Pilze sind von der Feuchtigkeit abhängig. Die meisten dieser Arten mögen Beauvaria bassiana werden durch freie Feuchtigkeit begünstigt.
Wir erhöhen das Risiko, wenn die optimale Temperatur zwischen 20 und 32°C und die relative Luftfeuchtigkeit zwischen 30 und 95% r. h. liegt, und verringern das Risiko, wenn es zu nass ist (Blattnässe, Regen oder relative Luftfeuchtigkeit über 95%), zu kalt (Temperaturen unter 20°C) oder zu heiß (Temperaturen über 32°C), nasse Nächte (Blattnässe in den Nachtstunden).