Робота базується на двох наступних статтях:
- Хвороба чорної плямистості листя; Шкідники та хвороби Американського Самоа Номер 10, Громадський коледж Американського Самоа, Кооператив з досліджень та поширення знань про громаду та природні ресурси. 2004
- Чорна Сігатока - зростаюча нитка до вирощування бананів; Дуглас Х. Марін, Del Monte Fresh Produce; Рональд А. Ромеро, Chiquita Brands; Маурісіо Гусман, Національна бананова корпорація Коста-Ріки; Тернер Б. Саттон, Державний університет Північної Кароліни, Ролі
Чорна плямистість листя (ЧПЛ), або "чорна сігатока", є найбільш шкідливою хворобою листя бананів у всьому світі. Більшість районів тропіків і субтропіків уражені цією хворобою, втрати становлять 30-50%. Перше повідомлення про BLS в Тихоокеанському регіоні з'явилося 40 років тому в долині Сігатока, Фіджі. Більшість фунгіцидів, що використовуються у виробництві бананів, призначені для боротьби з ВБС. Банан сорту Кавендіш "Вільямс", який вирощують більшість комерційних виробників, дуже сприйнятливий до грибкового захворювання. Заражені рослини мають менше листя, що призводить до меншої кількості плодів, затримки збору врожаю, передчасного дозрівання і зниження якості плодів.
Потенційний вплив
Чорна сігатока викликає руйнування тканин бананового листя, що впливає на фотосинтетичні можливості рослини і може знизити врожайність до 50%. Чорна сігатока є одним з основних факторів, що спричиняють занепад експортної галузі бананів у країнах південної частини Тихого океану. Комерційні плантації, що вирощують банани на експорт, змушені підтримувати дороговартісну програму обприскування фунгіцидами для боротьби з чорною сигАТОКОЮ, яка піддається критиці з міркувань захисту довкілля та здоров'я людей. Однак, якщо не контролювати хворобу, фрукти, вирощені на хворих рослинах, можуть передчасно дозріти під час транспортування і спричинити подальші збитки. У наступні роки хворобу контролювали за допомогою хімічних обприскувань (15-17 обробок фунгіцидами), але це значно збільшило собівартість продукції.
Постраждалі рослини
Певні симптоми чорної сігатоки були зафіксовані лише на Муса! (Банани), Муса райська (Подорожники), Муса гостра (Musa acuminata) (Дикий банан) та Муса гостра (Musa acuminata) (підп. bantesii і підполковник. zebrina). Культурні сорти відрізняються за своєю реакцією на патоген. Імунітет невідомий, і можливо, що інші дикі види та підвиди Муса! інфіковані, але хвороба не набуває значного розвитку. Mycosphaerella fijiensis також може атакувати саджанці диких бананів на стадії проростків.
Сезон появи
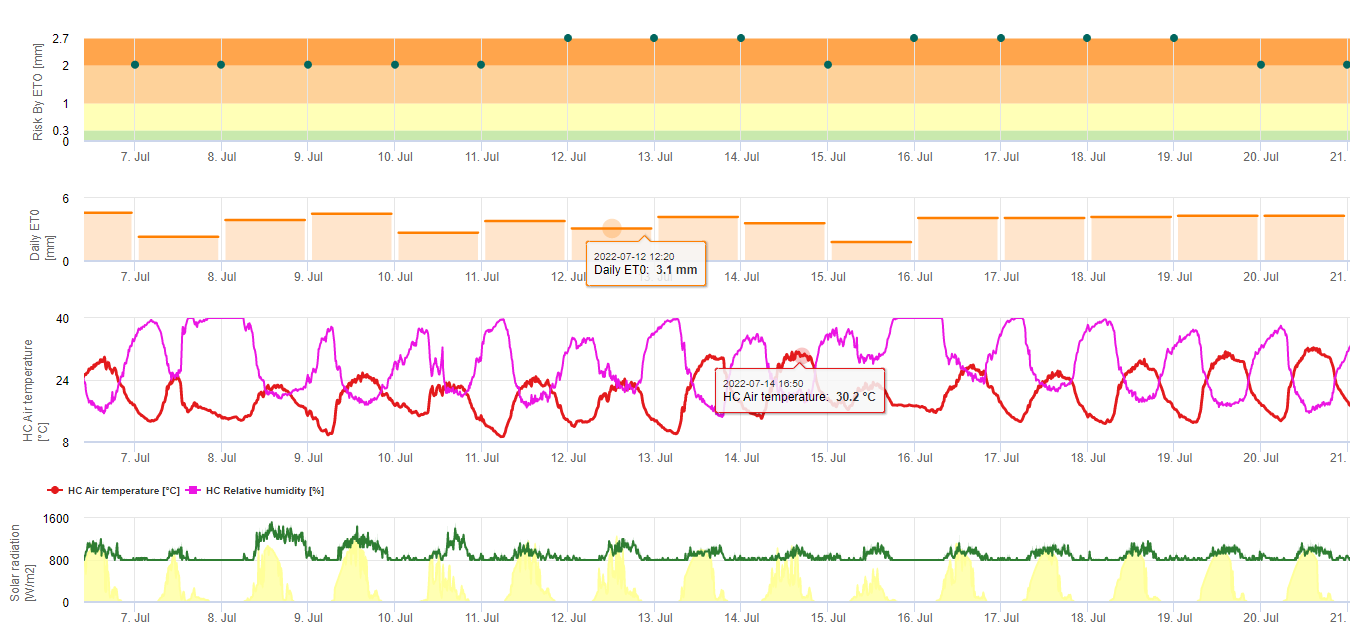
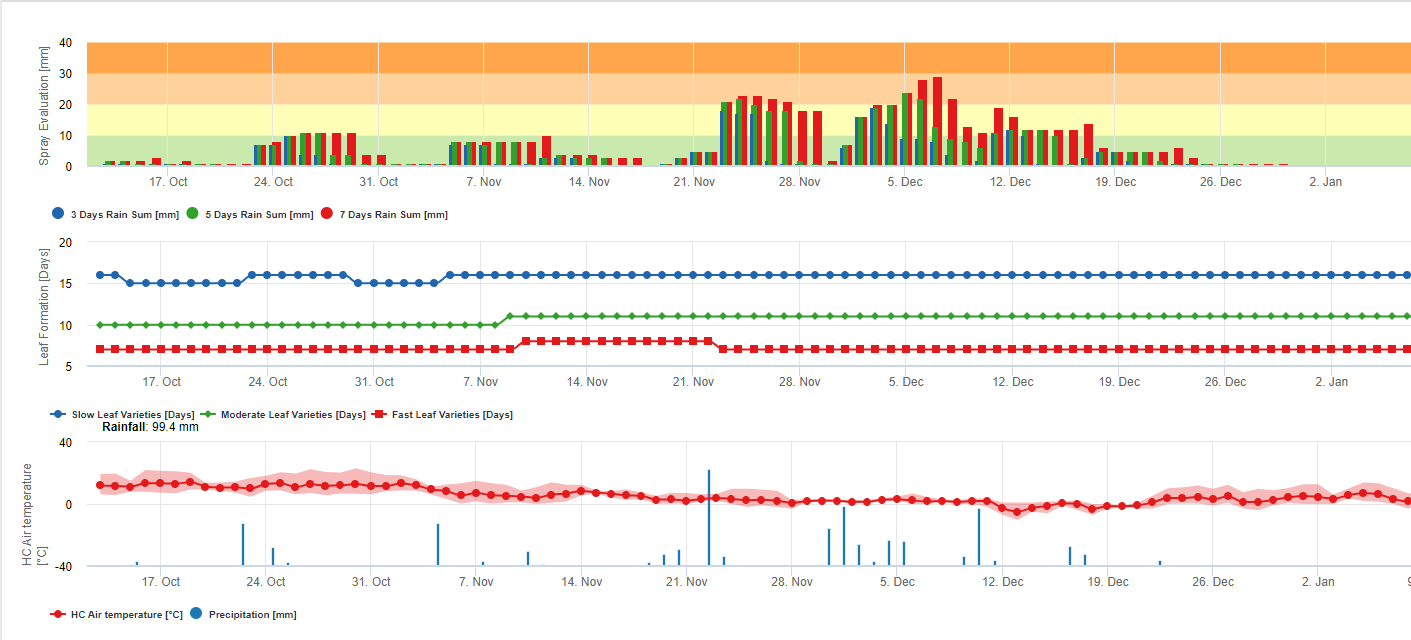
Спори чорної сігатоки легко утворюються протягом тропічного та субтропічного літа або в умовах висока вологість і опадиособливо якщо на листках є плівка вільної води. Основним способом поширення є дощовий змив або розбризкування спор, але на більш пізніх стадіях розвитку хвороби спори також примусово викидаються в повітряні потоки. Зараження відбувається на наймолодших листках рослини під час і відразу після розгортання, але старіші листки є більш сприятливими для розвитку хвороби. Очевидні візуальні симптоми з'являються приблизно через 15-20 днів після першого зараження.
Симптоми
Першими видимими симптомами зараження листя є короткі червонувато-коричневі лінії довжиною близько 2 мм. Лінії перетворюються на коричневі смуги довжиною 20-30 мм, які розширюються в еліптичні плями, коричневі на нижній стороні листка, чорні на верхній поверхні. Плями злегка заглиблені і оточені жовтуватим ореолом. Потім центр великої плями стає сірим, облямованим чорною лінією і жовтим ореолом. Нарешті, плями зливаються, вбиваючи листок.
Епідеміологія
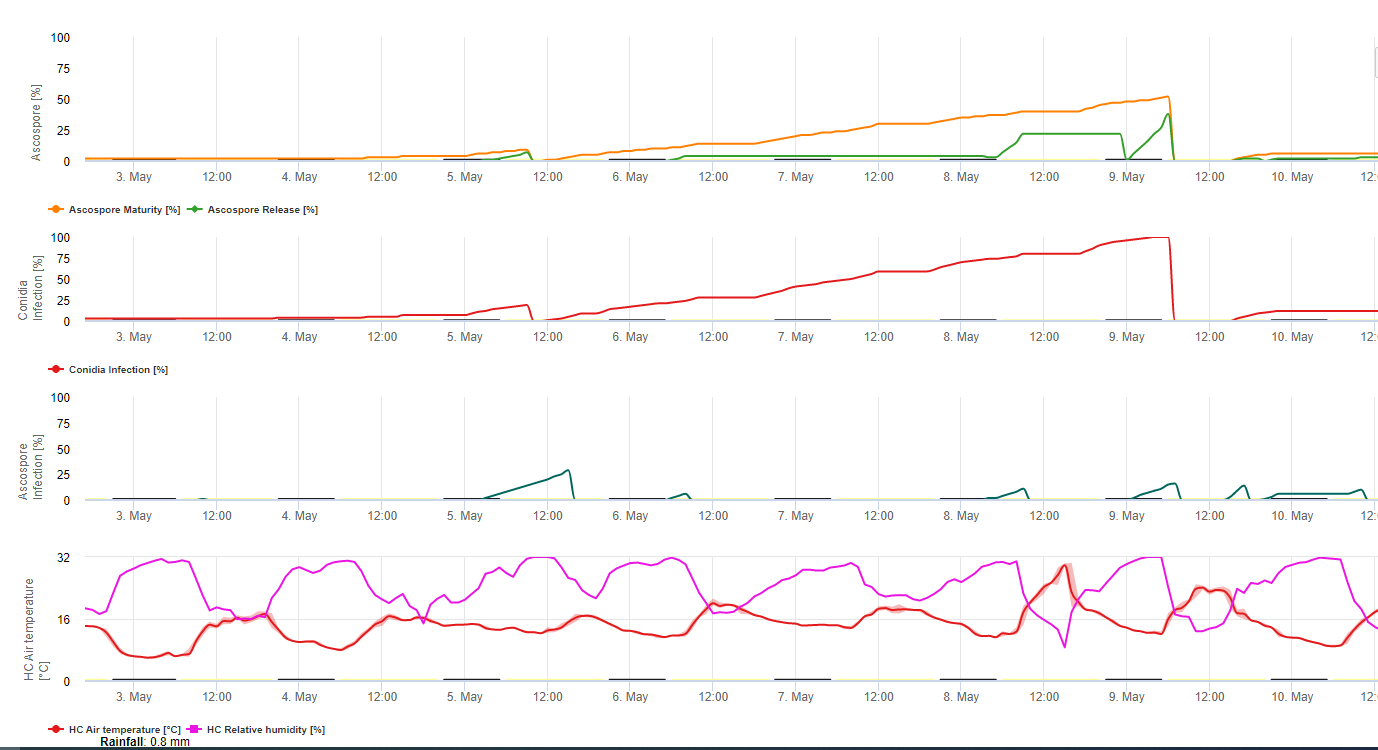
Конідії та аскоспори відіграють певну роль у поширенні хвороби. Конідії утворюються за умов підвищеної вологостіособливо якщо на листках є плівка вільної води. Вони утворюються під час розвитку перших стадій хвороби (особливо під час стадії 2, стадії 3 і стадії 4 плямистості). Основними шляхами поширення є змив і розбризкування дощу; конідії не розносяться вітром. Конідії асоціюються здебільшого з місцевим поширенням хвороби і відіграють важливу роль у періоди високої вологості, частих рясних рос і періодичних злив.
Тому що M. fijiensis виробляє відносно мало конідій, аскоспори вважаються більш важливими у поширенні чорної сігатоки (4,20). Однак Jacome та ін. (10) стверджують, що конідії здатні викликати значні обсяги захворювання, а симптоми хвороби ідентичні тим, що викликаються аскоспоровою інфекцією. Він зазначив, що конідії набувають більшого значення в посушливі періоди, коли розвиток хвороби затримується через наявність менш сприятливих кліматичних умов (10). Аскоспори є основним засобом розосередження на більші відстані в межах плантацій і на нові території, і є звичайним засобом поширення під час тривалих періодів вологої погоди (3,4,6,10, 11,15).
Аскоспори утворюються в псевдотеціях у зрілих уражених ділянках, які часто зустрічаються на старому листі рослини або на мертвому листі, що лежить на землі. Мередіт і Лоуренс (14) повідомляють, що псевдотеції утворюються з обох боків листкової поверхні, але більша їх кількість присутня на адаксіальній поверхні. І навпаки, Gauhl та ін. (6) виявили, що більше псевдотеціїв та аскоспор утворюється на абаксіальній поверхні листка. Хоча Burt et al. (2) виявили, що на один псевдотецій вивільняється приблизно 4,5 аскоспор, це число здається дуже низьким, враховуючи високу концентрацію інокуляту, яка присутня в повітрі після дощу (4).
Вивільнення аскоспор
Потрібна наявність плівка води від дощу або роси який поглинає псевдотеції і призводить до примусового викиду аскоспор через прикордонний шар листка, де вони поширюються повітряними потоками (15,20). Дозрівання псевдотеції вимагає насичення з мертвих тканин листя протягом приблизно 48 годин (3,4,7,10,11,15). В умовах Гавайських островів концентрація аскоспор зростала вночі, була найвищою близько 06:00, а вдень значно знижувалася. У дощові дні пікові концентрації спостерігалися незабаром після початку дощу. Сезонне збільшення середньодобових концентрацій аскоспор пов'язане зі збільшенням кількості опадів і відносної вологості (15). Gauhl (4), працюючи з бананами, повідомив про подібні результати в умовах Коста-Ріки. У той час як Мередіт та ін. (15) не знайшли доказів того, що на виробництво або вивільнення аскоспор впливають мінімальні температури, Гауль (4) визначив, що виробництво посівного матеріалу зменшується в більш сухі (або менш дощові) місяці року в Карибській зоні Коста-Ріки, що також є сезоном з найнижчими температурами. Аскоспори розносяться вітром, проте їхнє поширення на великі відстані обмежене кількома сотнями кілометрів через їхню чутливість до ультрафіолетового випромінювання (17). Послідовного зв'язку між викидом аскоспор і розвитком хвороби не виявлено, що обмежує корисність відлову спор для прогнозування розвитку хвороби (5,11). Аскоспори осідають переважно на нижній поверхні листка під час розгортання нового листка, створюючи смуговий характер інфекції на стороні, яка відкривається першою, що є відображенням підвищеного осідання спор на циліндричному листі кандели під час його розгортання, на відміну від всієї відкритої пластинки листка. Отже, більшість інфекцій відбувається на абаксіальній поверхні листків (3,7).
Інфекція
Зародкові трубки проникають у продихи приблизно за 48-72 години (1,20), хоча гриб може рости епіфітно на поверхні листка до 6 днів, перш ніж фактично проникнути в листя (1). Успішне зараження просувається за рахунок розширеного періоди підвищеної вологості та наявності вільної води на листках (3,7,13). Максимальне проростання відбувається за наявності вільної води. Конідії проростають у ширшому діапазоні відносної вологості (від 92 до 100%), ніж аскоспори (від 98 до 100%). Вплив температура на проростання можна охарактеризувати квадратичною функцією відгуку з оціночним оптимумом 26,5°C (10). Стовер (21) спостерігав максимальний ріст зародкових трубок аскоспор при від 26 до 28°C після 24-годинної інкубації. Jacome і Schuh (8,9) повідомляють, що старіші листки більш сприйнятливі, проте E. Bureau (особисте повідомлення) і Romero (18) відзначають, що молоді листки більш сприйнятливі (від першого до третього), ніж старі.
Інкубаційний період
Час між зараженням і появою симптомів варіює залежно від погодних умов і сприйнятливості рослин (68). У бананів час зараження листя збігається з появою нового листка з верхівки псевдостебла (20). За дуже сприятливих умов у Коста-Ріці та за наявності сприйнятливого хазяїна інкубаційний період може становити від 13 до 14 днів, тоді як у періоди несприятливої погоди тривалість інкубаційного періоду може збільшуватися до 35 днів. Подібні повідомлення існують з Нігерії щодо подорожників (16). У сезон дощів інкубаційний період становив 14 днів, а в сухий сезон - 24 дні. Тривалість інкубаційного періоду також залежить від сприйнятливості сорту до M. fijiensis. Інкубаційний період становив 26,2 і 25,1 днів, відповідно, у сортів FHIA 1 і FHIA 2, двох тетраплоїдних синтетичних гібридів, розроблених FHIA, порівняно з 22,4 днями у сприйнятливого сорту Grande Naine (19).
Після проникнення в листок гіфи <M. fijiensis колонізують сусідні клітини протягом приблизно 7 днів без будь-яких ознак руйнування клітин. Вегетативні гіфи можуть виходити з продихів і рости на поверхні листка, проникати в сусідні продихи або утворювати конідієносці та конідії. Цей епіфітний ріст дозволяє грибу колонізувати прилеглі тканини листка, що призводить до швидкого розвитку симптомів (4,20). M. fijiensis має більшу здатність проникати в декілька продихів, ніж М. Музиколащо призводить до більшої плямистості, ніж у випадку з попереднім патогеном (20).
Латентний період
Хоча конідії можуть утворюватися в ураженнях з ранніми симптомами M. fijiensis латентний період визначається часом, коли гриб починає продукувати ураження зрілими псевдотеціями та аскоспорами, які є основним джерелом інокуляту. Як і інкубаційний період, латентний період також залежить від погодних умов, сприйнятливості хазяїна та інтенсивності інфекції. У Гуапілес, Коста-Ріка, латентний період становив від 25 днів у сезон дощів (з червня по грудень) до 70 днів у сухий сезон. Коли погода дуже сприятлива для виділення аскоспор та інфікування, багато інфекцій з'являється на листках. Коли інфекції є щільними, вони швидко об'єднуються на дуже ранній стадії розвитку, прискорюючи появу зрілих плям, які характеризуються наявністю псевдотеціїв та аскоспор (22). За таких умов листя швидко і сильно пошкоджується. Латентний період також варіює залежно від рівня стійкості. Наприклад, час від появи листя до першого симптому зрілої плямистості в однакових природних умовах для сорту Curraré, кулінарного банана, що належить до підгрупи подорожникових, становив 44 дні порівняно з 34 днями для сорту Valery, банана, що належить до підгрупи кавендіш (4). Термін "час розвитку хвороби" широко використовується в літературі про банани в тропіках для позначення латентного періоду і визначається як час між зараженням і формуванням зрілих плям. Іншим поширеним терміном, який використовується в літературі про банани, є "час розвитку симптомів" або "перехідний період", тобто час від появи перших симптомів до появи зрілих плям (6). Час розвитку симптомів дає хороше уявлення про те, як швидко хвороба прогресує на листках.
Література:
- Beveraggi, A. 1992. Etude des interactions hôte-parasite chez des bananiers sensibles et résistants inoculés par Cercospora fijiensis responsible de la maladie des raies noires. Дисертація 3-го циклу. Університет Монпельє, Монпельє, Франція.
- Burt, J. P. A., Rosenberg, L. J., Rutter, J., Ramírez, F., and González, O. H. 1999. Прогнозування повітряно-крапельного поширення Mycosphaerella fijiensis, збудника чорної хвороби Сігатока на бананах: Оцінка кількості перитеціїв та аскоспор. Ann. Appl. Biol. 135:369-377.
- Фуллертон, Р. А. 1994. Хвороби листя сігатоки. Сторінки 12-14 в: Компендіум хвороб тропічних фруктів. Р. К. Плоц, Г. А. Зентмайєр, В. Т. Нішиджинія, К. Г. Рорбах та Х. Д. Ор, редактори. Американське фітопатологічне товариство, Сент-Пол, штат Міннесота.
- Gauhl, F. 1994. Епідеміологія та екологія чорної сигатоки (Mycosphaerella fijiensis Morelet) на подорожнику та бананах (Musa spp.) в Коста-Ріці, Центральна Америка. INIBAP, Монпельє, Франція.
- Gauhl, F. 2002. Чи є спорові пастки корисним інструментом для прогнозування розвитку хвороби Сігатока на комерційних бананових плантаціях. Сторінка 22 в: Int. Семінар з мікофорельозної плямистості листя бананів, 2-й - Програма і тези. Банани, 2-й - Програма та тези доповідей. (Abstr.)
- Gauhl, F., Pasberg-Gauhl, C., and Jones, D. R. 2000. Цикл хвороби та епідеміологія. Сторінки 56-62: Хвороби банана, абаки та енсету. Д. Р. Джонс, ред. CAB International, Уоллінгфорд, Великобританія.
- Гонсалес, М. 1987. Enfermedades del cultivo del banano. Oficina de Publicaciones de la Universidad de Costa Rica, San José.
- Jacome, L. H., and Schuh, W. 1992. Вплив тривалості та температури зволоження листя на розвиток чорної хвороби Сігатока на бананах, інфікованих Mycosphaerella fijiensis var. difformis. Фітопатологія 82:515-520.
- Jacome, L. H., and Schuh, W. 1993. Вплив температури на ріст і конідієносність in vitro, а також порівняння інфекції та агресивності in vivo серед ізолятів Mycosphaerella fijiensis var. difformis. Trop. Agric. 70:51-59.
- Jacome, L. H., Schuh, W., and Stevenson, R. E. 1991. Вплив температури та відносної вологості на проростання та розвиток зародкової трубки Mycosphaerella fijiensis var. difformis. Фітопатологія 81:1480-1485.
- Jeger, M. J., Eden-Green, S., Thresh, J. M., Johanson, J. M., Waller, J. M., and Brown, A. E. 1995. Бананові хвороби. Сторінки 317-381: Банани та подорожники. С. Гоуен, ред. Chapman & Hall, Лондон.
- Марін, Д. Х. та Ромеро, Р. А. 1992. El combate de la Sigatoka negra. Бюлетень № 4, Департамент розслідувань, Національна банківська корпорація, Коста-Ріка.
- Mayorga, M. H. 1990. La Raya negra (Mycosphaerella fijiensis Morelet) del plátano y del banano. I. Ciclo de vida del patógeno bajo las condiciones del Urabá. Revista ICA 25:69-77.
- Мередіт, Д. С., і Лоуренс, Д. С. 1969. Хвороба чорної плямистості листя бананів (Mycosphaerella fijiensis): Симптоми хвороби на Гаваях та примітки щодо конідіального стану гриба-збудника. Trans. Br. Mycol. Soc. 52:459-476.
- Мередіт, Д. С., Лоуренс, Д. С. та Фірман, І. D. 1973. Вивільнення та розсіювання аскоспор при хворобі чорної смугастості листя бананів (Mycosphaerella fijiensis). Trans. Br. Mycol. Soc. 60:547-554.
- Mobambo, K. N., Gauhl, F., Pasberg-Gauhl, C., and Zuofa, K. 1996. Оцінка впливу сезону та віку рослин на реакцію подорожника на хворобу чорної сигатоки. Crop Prot. 15:609- 614.
- Парнелл, М., Берт, П. Д. А. і Вілсон, К. 1998. Вплив впливу ультрафіолетового випромінювання в імітованому сонячному світлі на аскоспори, що викликають чорну хворобу Сігатока бананів і подорожника. Int. J. Biometeorol. 42:22-27.
- Ромеро, Р. А. 1995. Динаміка стійких до фунгіцидів популяцій Mycosphaerella fijiensis та епідеміологія чорної сігатоки бананів. Дисертація на здобуття наукового ступеня доктора філософії. Університет Північної Кароліни, Ролі.
- Ромеро, Р. А. та Саттон, Т. Б. 1997. Реакція чотирьох генотипів Musa при трьох температурах на ізоляти Mycosphaerella fijiensis з різних регіонів. Plant Dis. 81:1139-1142.
- Стовер, Р. Х. 1980. Сигатозна плямистість листя бананів і подорожників. Plant Dis. 64:750-756.
- Стовер, Р. Х. 1983. Вплив температури на ріст зародкових трубок аскоспор Mycosphaerella musicola та Mycosphaerela fijiensis var. difformis. Fruits 38:625-628.