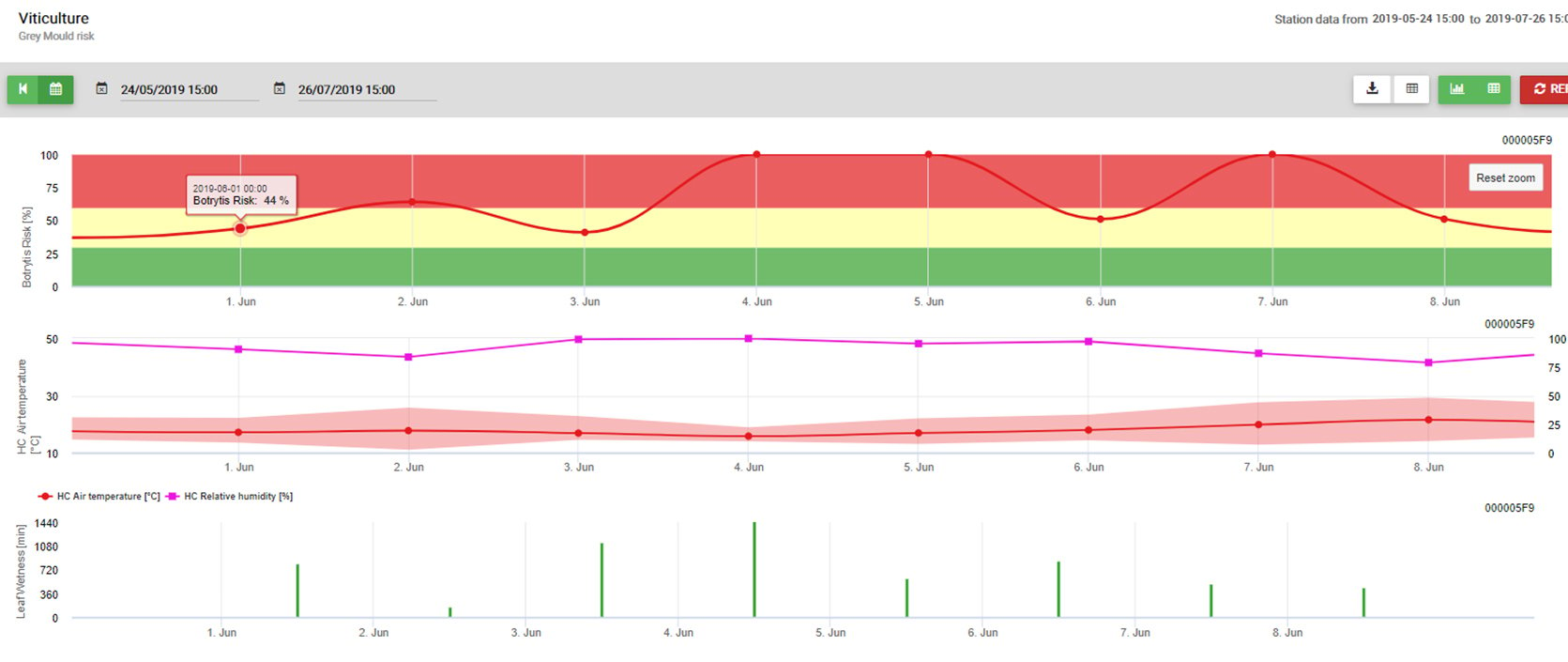
Cykl życia
Patogen zimuje na zainfekowanych resztkach pożniwnych i nasionach. Zainfekowane nasiona mogą odgrywać ważną rolę zarówno we wprowadzaniu zarazy Ascochyta na nowe tereny, jak i we wczesnym rozwoju choroby, ponieważ patogen jest łatwo przenoszony z nasion na siewki. Zarówno bezpłciowe zarodniki (konidia, rozprzestrzeniane przez deszcz) jak i zarodniki płciowe (askospory, rozprzestrzeniane przez wiatr) mogą być produkowane na resztkach pożniwnych. Późną jesienią i wczesną wiosną w wyniku rozmnażania płciowego powstają pseudotecje, w których znajdują się askospory. Rozwój pseudotecjów trwa pięć do siedmiu tygodni w warunkach odpowiedniej wilgotności i umiarkowanej temperatury (około 10°C).
Wiosną i wczesnym latem dojrzałe pseudotecje uwalniają askospory, które mogą przemieszczać się na odległość kilku kilometrów, aby zainfekować docelowe uprawy. Uważa się, że unoszące się w powietrzu askospory są początkowym źródłem infekcji wiosną, chociaż mogą w tym uczestniczyć również konidia rozrzucone przez deszcz. Po zakażeniu zarodnikami, objawy zaczynają się rozwijać w ciągu czterech do sześciu dni. Wczesne zmiany są brązowe do ciemnobrązowych, z ciemnobrązowym marginesem. Trzy do sześciu dni po powstaniu zmian rozwijają się ciemnobrązowe piknidia.
Piknidia są często ułożone w koncentryczne pierścienie i nie ulegają ścieraniu, natomiast szczątki przypominające tkanki ulegają ścieraniu. Konidia wydostają się z piknidiów w postaci lepkiej masy zarodników i są rozprowadzane przez deszcz na zdrowe części roślin, powodując nowe infekcje.
Większość zmian chorobowych w sezonie wegetacyjnym wynika z szybkiego rozwoju piknidiów i konidiów w wilgotnych warunkach. Do rozprzestrzeniania konidiów na nowe tkanki roślinne wystarczą nawet niewielkie opady deszczu. Jest więc nazywana chorobą policykliczną, co oznacza, że w sezonie wegetacyjnym przy odpowiedniej wilgotności i temperaturze (20-25°C) może wystąpić wiele cykli infekcji.
Model choroby
Modelujemy rozwój piknidiów i konidiów podczas sezonu wzrostu.
Rozwój choroby Ascochyta jest optymalny w temperaturze od 20 do 21°C (zakres temperatur wynosi od 4°C do 34°C) oraz w warunkach wilgotnych (wysoka wilgotność względna i zwilżenie liści).
W modelu uwzględniono dwa etapy cyklu chorobowego: 1. sporulację (rozwój piknidiów, uwalnianie konidiów), czyli wytwarzanie nowych jednostek zakaźnych oraz 2. optymalne warunki dla dalszej infekcji tych konidiów. Tak więc dla zakażenia nowej tkanki roślinnej najpierw zostały zapewnione optymalne warunki dla sporulacji (100%), a następnie krzywa infekcji osiągnęła 100%.
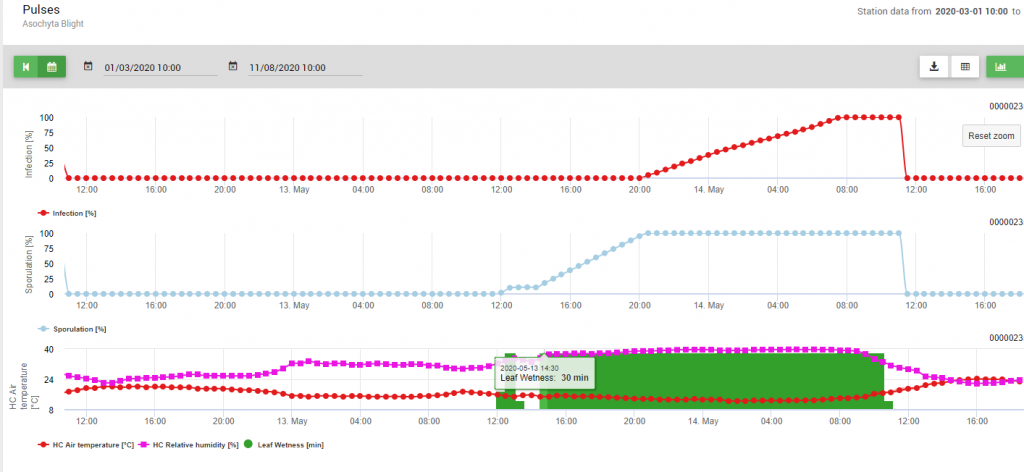
Ze względu na optymalne warunki (wilgotność liści, wysoka wilgotność względna i temperatura około 15°C przez długi czas) sporulacja piknidiów została określona 13 maja o godzinie 20:00 (niebieska linia, 100%). Warunki były nadal optymalne do rozpoczęcia postępu infekcji (czerwona linia), a optymalne warunki do infekcji zostały określone 14 maja o godzinie 8: 00 rano (osiągnięto 100%). Tak więc w zależności od strategii ochrony roślin należało uwzględnić pomiary profilaktyczne już przed wystąpieniem infekcji lub przy stosowaniu zabiegów leczniczych krótko po stwierdzeniu infekcji (100%).
Literatura:
Botrytis cinerea jest grzybem nekrotroficznym, który poraża wiele gatunków roślin, choć jego najbardziej godnymi uwagi gospodarzami mogą być winogrona.
W uprawie winorośli powszechnie znana jest jako zgnilizna pęczkowa botrytis; w ogrodnictwie nazywana jest zwykle szarą pleśnią lub szarą pleśnią.
Grzyb ten powoduje dwa różne rodzaje infekcji na winogronach. Pierwszy z nich, szara zgnilizna, jest wynikiem stale wilgotnych lub mokrych warunków i zazwyczaj prowadzi do utraty dotkniętych nią kiści. Drugi rodzaj, szlachetna zgnilizna, pojawia się, gdy po wilgotnych warunkach następują bardziej suche, a jego efektem mogą być charakterystyczne słodkie wina deserowe, takie jak Sauternes czy Aszú z Tokaju. Nazwa gatunkowa Botrytis cinerea pochodzi od łacińskiego "winogrona jak popiół"; choć poetyckie, "winogrona" odnosi się do zgrupowania zarodników grzyba na ich konidioforach, a "popiół" po prostu odnosi się do szarawego koloru mas zarodników. Grzyb określany jest zwykle nazwą anamorf (forma bezpłciowa), ponieważ faza płciowa jest rzadko obserwowana. Teleomorfem (formą płciową) jest ascomycete, Botryotinia cinerea.
Biologia B. cinerea
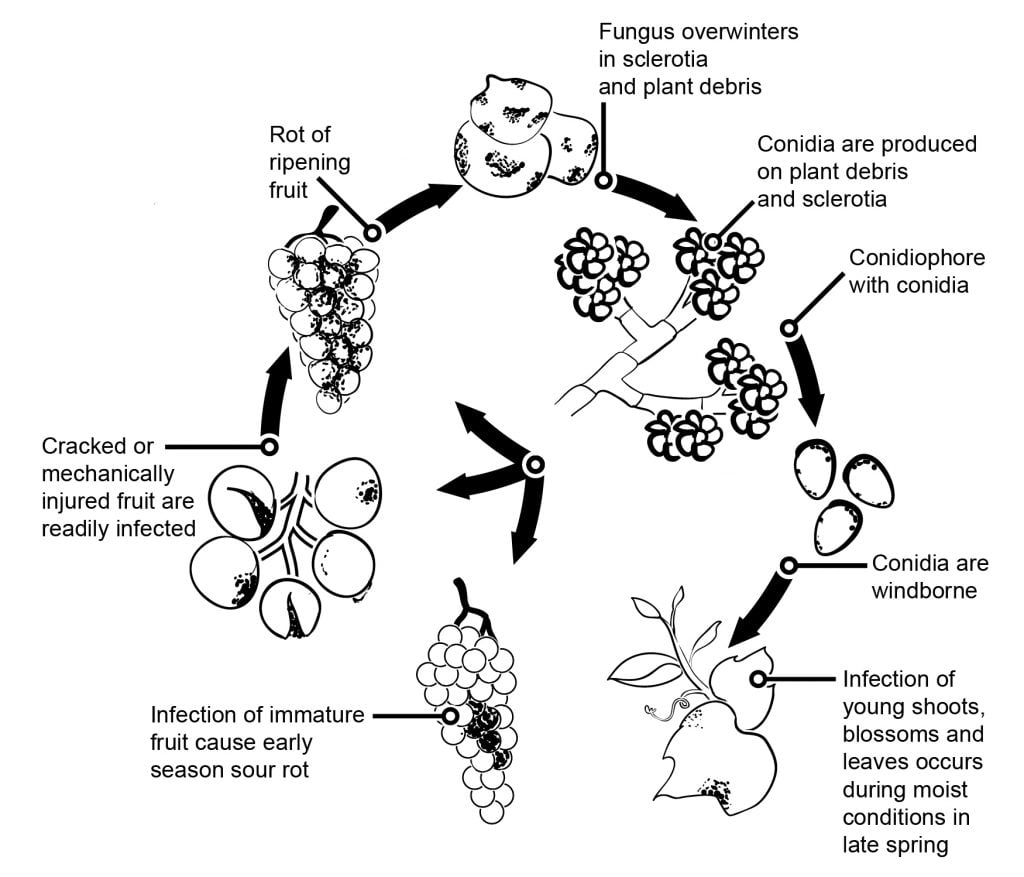
W przypadku owoców i winogron infekcje kwiatowe, po których następuje okres utajenia, mają duży wpływ na epidemiologię szarej pleśni. Można wyróżnić kilka ścieżek infekcji od kwiatów do owoców. W winogronach, kiwi, malinach postuluje się infekcje przez trzonek do zalążni. W zalążni patogen pozostaje w stanie latencji, co wydaje się być wynikiem wstępnie ukształtowanej strategii obronnej gospodarza (podobnie jak zawartość resweratrolu w młodych jagodach winogron). W winogronach, kiwi i truskawkach istotne są infekcje przez pręcik, płatki lub działkę. W winogronach badania wykazały, że B. cinerea może zainfekować pręciki i rośnie bazipetalnie, aby zainfekować zawiązek, a następnie rozrasta się systemowo do szypułki i tkanek naczyniowych w jagodach.

Sześcioletni projekt badawczy w rejonie Cape wykazał, że jagody winogron mogą być zakażone przez stomię i przetchlinki szypułki oraz w dolnej części rachityki. Infekcje szypułki są możliwe także w okresie kwitnienia. Później ta tkanka zwiększa odporność na B. cinerea zakażenia.
Inne drogi infekcji postulują saprofityczny rozwój patogenu na resztkach kwiatowych i późniejsze zakażenie jagód, gdy podatność wzrasta wraz z dojrzewaniem lub w wyniku uszkodzenia jagód przez owady lub grad. Inną tezą jest założenie o gromadzeniu się konidiów wewnątrz owoców w okresie letnim i zakażaniu podatnych jagód w późniejszym okresie sezonu. Zakażenie konidialne dojrzewających owoców jest możliwe z każdego źródła inokulum. Najprawdopodobniej powstaje niewielka liczba utajonych zakażonych jagód, które wykazują rozległą sporulację, gdy podatność jagód wzrasta wraz z dojrzałością. Można przyjąć, że jagody stają się podatne od zawartości cukru 6%.
W owocach kiwi mamy duży wpływ warunków zbioru na występowanie B. cinerea. Owoce zbierane z mokrą powierzchnią mogą zostać zainfekowane przez B. cinerea na mikro zmiany ustawione przez palce zbieraczy.
W praktycznej kontroli B. cinerea musimy wydzielić dwa ważne okresy infekcji: Kwitnienie i starość. 1) Podczas kwitnienia mamy podatne młode owoce, gdzie po infekcji następuje okres latencji. 2) Podczas gdy infekcja na dojrzałych (starzejących się) owocach prowadzi do objawów bez okresu latencji. Znaczenie infekcji podczas kwitnienia w winogronach może się zmieniać w zależności od sezonu i regionu. W przypadku owoców, na których musimy spodziewać się pewnej trwałości (winogrona stołowe, kiwi czy truskawki) objawy widoczne są podczas przechowywania w warunkach chłodniczych w sklepach lub magazynach. Chemiczne zwalczanie winogron wykazujących dobrą odporność na B. cinerea w czasie kwitnienia nie przyniesie żadnych efektów ekonomicznych. Dlatego przy podejmowaniu decyzji o zastosowaniu przeciwko należy wziąć pod uwagę wszystkie warunki ryzyka i prawdopodobieństwa infekcji, podatność owoców oraz okres trwałości, warunki przechowywania. Botrytis cinerea podczas kwitnienia.
W owocach pestkowych zakażenie przez B. cinerea występuje najczęściej w okresie kwitnienia. W tym czasie zabiegi przeciwko Monilina spp. są brane pod uwagę, które również infekują Botrytis cinerea infekcja.
Model B. cinerea i praktyczne zastosowanie
Potrzebne czujniki: Wilgotność liści, temperatura, wilgotność względna
Botrytis cinerea jest fakultatywnym pasożytem. Rośnie również na martwym materiale roślinnym. Z tego powodu jest zawsze obecny w winnicach i sadach. Botrytis cinerea jest związana z wilgotnym klimatem. Do infekcji potrzebuje bardzo wysokiej wilgotności względnej lub obecności wolnej wody (czujnik: wilgotność liści, wilgotność względna). Grzyb nie jest w stanie zainfekować zdrowego, dorosłego materiału roślinnego przez zarodniki. Infekcja następuje na młodych pędach winorośli podczas dłuższych okresów wilgotnych lub na pędach uszkodzonych przez burze gradowe.
Do obliczenia ryzyka infekcji model wykorzystuje następującą korelację pomiędzy czasem trwania zwilżenia liści a temperaturą.
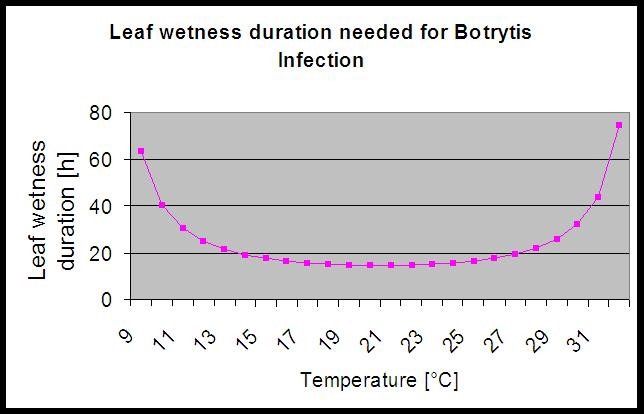
Na stronie Botrytis Model ryzyka daje w wyniku wartość ryzyka od 0 do 100%. Wartość ta wskazuje na ciśnienie B. cinerea w danym czasie. Jeśli mamy wartość 100% oznacza to, że kilkakrotnie wystąpił okres mokry na tyle długi, aby zainfekować podatną tkankę (obliczamy tzw. "punkty mokre" (macierz pomiędzy mokrością liścia, temperaturą z maksimum początkowo 38400 punktów (początek sezonu, który wykazuje ryzyko 30%). Po tym okresie każdy okres mokry z około 4000 punktów mokrych (macierz) zwiększa ryzyko o 10% lub z drugiej strony każdy okres suchy zmniejsza ryzyko o 1/5 poprzedniej wartości.
Wniosek przeciwko B. cinerea zależy od owoców i celu produkcji.