





El tizón de la fusariosis o costra de los granos pequeños está causado por el hongo Fusarium graminearum (Schwabe), aunque los Países Bajos y otras zonas de Europa central informan de F. culmorum como la especie más prevalente (Snidjers, 1989). En Polonia, F. culmorum, F. graminearum y F. nivale han mostrado niveles similares de virulencia de moderada a grave, mientras que F. avenaceum ha demostrado ser de mediana a moderadamente virulenta. Sin embargo, en varios estudios destinados a identificar el organismo causal, hasta 18 Fusarium spp. fueron aislados e identificados (Mihuta-Grimm y Foster, 1989; Reis, 1985).
Artículo de L. Gilchrist, H.J. Dubin La sarna es frecuente en regiones cálidas y húmedas en las que la floración coincide con periodos lluviosos. La incidencia de esta enfermedad ha ido en aumento en los últimos diez años por diversas razones. Tal vez la razón más importante sea el aumento de la superficie en la que se rota el trigo con maíz u otros cereales. Otras razones son los cambios en el sistema de cultivo con fines de protección del suelo y los cambios en el cultivo de trigo de zonas tradicionales a zonas más húmedas y no tradicionales (Gilchrist et al., 1997).
La sarna del fusarium causa graves pérdidas de producción en todo el mundo, que pueden llegar al 50%. De forma similar, en Paraguay las condiciones climáticas de 1972 y 1975 favorecieron las epidemias de Fusarium y Septoria, que juntas causaron pérdidas de hasta el 70% (Viedma, 1989). Polonia, los Países Bajos, el Reino Unido, la antigua Checoslovaquia, la Federación Rusa, Francia y Austria son algunos de los países europeos que informan de la incidencia de la sarna. Zonas de Canadá, Estados Unidos, México, Guatemala, Brasil, Ecuador, Uruguay y Argentina en América están muy afectadas por Fusarium (Ireta y Gilchrist, 1994).
La sarna puede causar importantes daños en el rendimiento y la calidad, así como toxicosis en animales y humanos (Ireta y Gilchrist, 1994; Baht et al., 1989; Luo, 1988; Snidjers, 1989; Marasas et al., 1988). Los daños causados por la sarna en Estados Unidos se estimaron en más de $1.000 millones de USD en 1993 y en $500 millones de USD en 1994. En China, se calcula que la sarna puede afectar hasta a 7 millones de hectáreas, y se pueden perder 2,5 millones de toneladas de grano en años epidémicos. En China, India y Japón se han registrado enfermedades humanas relacionadas con la micotoxina de la fusariosis, mientras que en animales se han registrado enfermedades en numerosas partes del mundo (Dubin et al., 1997).
En la actualidad, existen normativas mundiales sobre micotoxinas (Van Egmond y Dekker, 1995). Sin embargo, en muchos países las normativas no se aplican, y muchas personas, sobre todo de zonas rurales, consumen los cereales sin ningún control, ya sea en grano o de forma indirecta como carne procedente de animales alimentados con grano contaminado. La tabla 16.1 muestra los principales efectos sobre los cerdos y las aves de corral inducidos por las toxinas más importantes producidas por algunos Fusarium especies.
Las condiciones climáticas húmedas y cálidas entre el espigado y la madurez aumentan la gravedad de la sarna. El punto de entrada de F. graminearum es la espiga, especialmente los órganos florales. Esto afecta al cuajado de las semillas y al llenado de los granos. Las espiguillas infectadas pierden rápidamente la clorofila y adquieren un color pálido. Más tarde se vuelven de color rosa o melocotón, especialmente en la base y los bordes de las glumas (Lámina 41). Si las condiciones ambientales siguen siendo favorables, la infección avanza a las espiguillas adyacentes y en algunos casos puede infectar toda la espiga, incluyendo el raquis y su pedúnculo. Cuando la infección es grave, los granos dañados se cubren de micelio y adquieren el aspecto de una masa algodonosa rosada. Si los niveles de la enfermedad son moderados, el grano puede estar arrugado, con poco peso y de color blanquecino (granos de lápida).
Principales efectos tóxicos en cerdos y aves de corral inducidos por las toxinas más importantes producidas por algunas especies de Fusarium
| Micotoxina | Signos clínicos |
| Zearalenona | Vulva roja e hinchada; prolapso vaginal en cerdos; los lechones pueden mostrar agrandamiento de las vulvas; problemas de fertilidad |
| Vomitoxina (desoxinivalenol, DON) | Disminución de la ingesta de alimento y aumento de peso en cerdos con DON a >2 mg/kg de alimento; vómitos; rechazo del alimento a concentraciones muy altas de DON (>20 mg/kg de alimento)a |
| Otros tricotecenos Toxina T-2 Toxina HT-2 Diacetoxiescirpenol> | Más tóxico que el DON; reducción de la ingesta de alimentos; emesis; irritación cutánea y gastrointestinal; neurotoxicidad; crías anormales; aumento de la sensibilidad a las enfermedades; hemorragias. |
| Ocharatoxina | Afecta principalmente a los túbulos proximales de los riñones en cerdos y aves de corral; los riñones están groseramente agrandados y pálidos; hígados grasos en aves de corral. |
amg/kg = partes por millón (ppm).
Fuente: Trenholm et al., 1984.
Fusarium graminearum (Lámina 42) puede atacar a las plantas de trigo en todas las etapas de crecimiento, causando enfermedades de pudrición de plántulas, tallos y raíces. Las infecciones primarias pueden surgir de ascosporas o macroconidias depositadas en glumas y anteras extruidas. Temperaturas de 10° a 30°C y humedad relativa superior al 95 por ciento durante 40 a 60 horas suelen ser suficientes para que los macroconidios infecten con éxito las espigas (Ireta, 1989).
Fusarium graminearum es uno de los pocos Fusarium especie que produce peritecios en condiciones de campo (Lámina 43, Lámina, 44). Los peritecios representan la fase sexual del hongo, Gibberella zeae, y se producen en las glumas del trigo. Los peritecios desempeñan un papel importante en la supervivencia del patógeno de un año a otro (Khonga y Sutton, 1988) y coexisten con micelios en residuos del cultivo anterior para constituir la fuente de inóculo inicial de la sarna. Las investigaciones llevadas a cabo en China demostraron que la temperatura más baja para la producción de peritecios era de 7° a 10°C, y la más adecuada de 15° a 20°C. El contenido de humedad del suelo más bajo para la producción de peritecios era del 50 al 60 por ciento, y el más adecuado del 70 al 80 por ciento (Wang, 1997).
Los factores que determinan el desarrollo de la enfermedad incluyen el clima, los niveles de inóculo y la fase de crecimiento del trigo. Las epidemias de sarna del trigo dependen principalmente de la cantidad de inóculo primario, más que del inóculo secundario. La invasión se produce sobre todo durante el periodo de floración. La infección secundaria con conidios (Lámina 45) se produce después de que las espigas enfermas aparezcan en el campo.
Los residuos de cultivos y las prácticas culturales desempeñan un papel importante en la conservación de F. graminearum y, en consecuencia, epidemias. Las infecciones en trigo sembrado en un campo de residuos de maíz pueden ser dos o tres veces más graves (Teich y Nelson, 1984). En general, el patógeno sólo era saprofito en rastrojos de arroz en las zonas de rotación arroz-trigo y en tallos de maíz en las zonas de siembra en seco de China (Wang, 1997). Si se ara el residuo, disminuye la supervivencia de los peritecios y se reduce la fuente primaria de inóculo (Reis, 1989).
Fusarium graminearum es un parásito facultativo y es patógeno en muchas otras gramíneas, incluidas malas hierbas comunes y cultivos de cereales (centeno, arroz, cebada y triticale). Si algunas prácticas, como la rotación con cultivos no hospedantes o la gestión de los residuos de cultivos, no son eficaces por sí solas, pueden, en combinación, reducir la fuente de inóculo primario. El control de la enfermedad se basa eficazmente en una gestión integrada, que incluye prácticas agronómicas adecuadas, la utilización de cultivares resistentes o tolerantes y aplicaciones químicas.
Biología del tizón de la cabeza por Fusarium
Patógenos ocasionales
El FHB está causado por especies fúngicas del género Fusarium. La especie más común causante de FHB es Fusarium graminearum (etapa sexual - Gibberella zeae). Este hongo es el mismo que se asocia con frecuencia a la podredumbre del tallo del maíz. Otro Fusarium especie causante de la FHB es Fusarium culmorum. Ambos F. graminearum y F. culmorum también puede causar la pudrición de las raíces de los granos pequeños. En la cebada, otros dos Fusarium especies, F. poae y F. avenaceumtambién puede causar el tizón de la semilla.
Supervivencia y propagación
El hongo persiste y se multiplica en los residuos de cultivos infectados de cereales pequeños y maíz. Cuando el tiempo es húmedo, las esporas del hongo son arrastradas por el viento o salpicadas sobre las cabezas de los cultivos de cereales. Las esporas pueden proceder del interior de un cultivo o ser arrastradas por el viento desde los cultivos circundantes, a veces a grandes distancias. Los cultivos de trigo y trigo duro son susceptibles de infección desde el periodo de floración (polinización) hasta la fase de desarrollo del grano. Las esporas del hongo causal pueden posarse en las anteras expuestas en el momento de la floración y luego crecer en los granos, glumas u otras partes de la cabeza. En el caso de la cebada de primavera, que florece cuando el cogollo está en la bota, la infección es más común después del periodo de floración, una vez que el cogollo atraviesa la vaina de la hoja. La infección en ambos cultivos puede continuar hasta cerca de la madurez del grano en condiciones ambientales favorables para el organismo u organismos.
Las condiciones más favorables para la infección son periodos prolongados (48 a 72 horas) de alta humedad y temperaturas cálidas (75 a 85 grados Fahrenheit (24°C a 30°C)). Sin embargo, la infección se produce a temperaturas más bajas cuando la humedad alta persiste durante más de 72 horas. Las infecciones tempranas pueden producir esporas transportadas por el aire, que son responsables de la propagación secundaria de la enfermedad, especialmente si el cultivo tiene una floración irregular debido a los macollos tardíos.
Dado que el desarrollo de la FHB depende de condiciones ambientales favorables desde la floración (aparición de la cabeza en la cebada) hasta el desarrollo del grano, la aparición y gravedad de la enfermedad varía de un año a otro. Una combinación de factores que puede conducir a las pérdidas más severas de rendimiento y calidad son: abundante inóculo, períodos prolongados o repetidos de humedad y alta humedad durante la floración (emergencia de la cabeza en la cebada) a través del desarrollo del grano, y el uso de un cultivar muy susceptible.
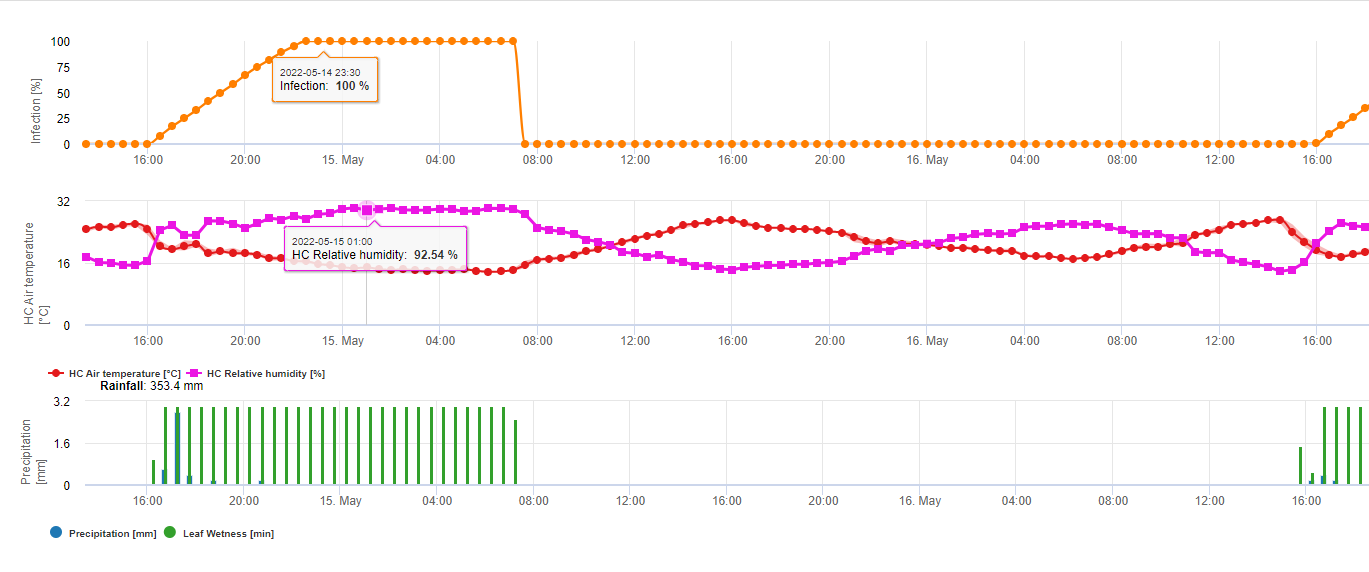
Modelo de infección por el tizón de la fusariosis
Se sabe que los agentes fúngicos pertenecientes al complejo Fusarium Head Blight en el trigo se ven favorecidos por temperaturas cálidas de 20 °C a 30 °C y largos periodos húmedos. Los periodos de varios días de humedad en las hojas provocan síntomas visibles tempranos. Pero los síntomas pueden observarse tras un periodo de latencia más largo si la infección va seguida de un periodo de humectación de la hoja de 18 horas o incluso menos. Los síntomas también pueden aparecer tras la inoculación artificial a temperaturas de 15°C.
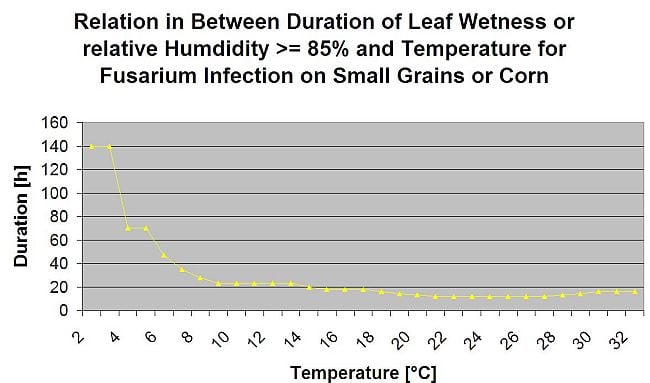

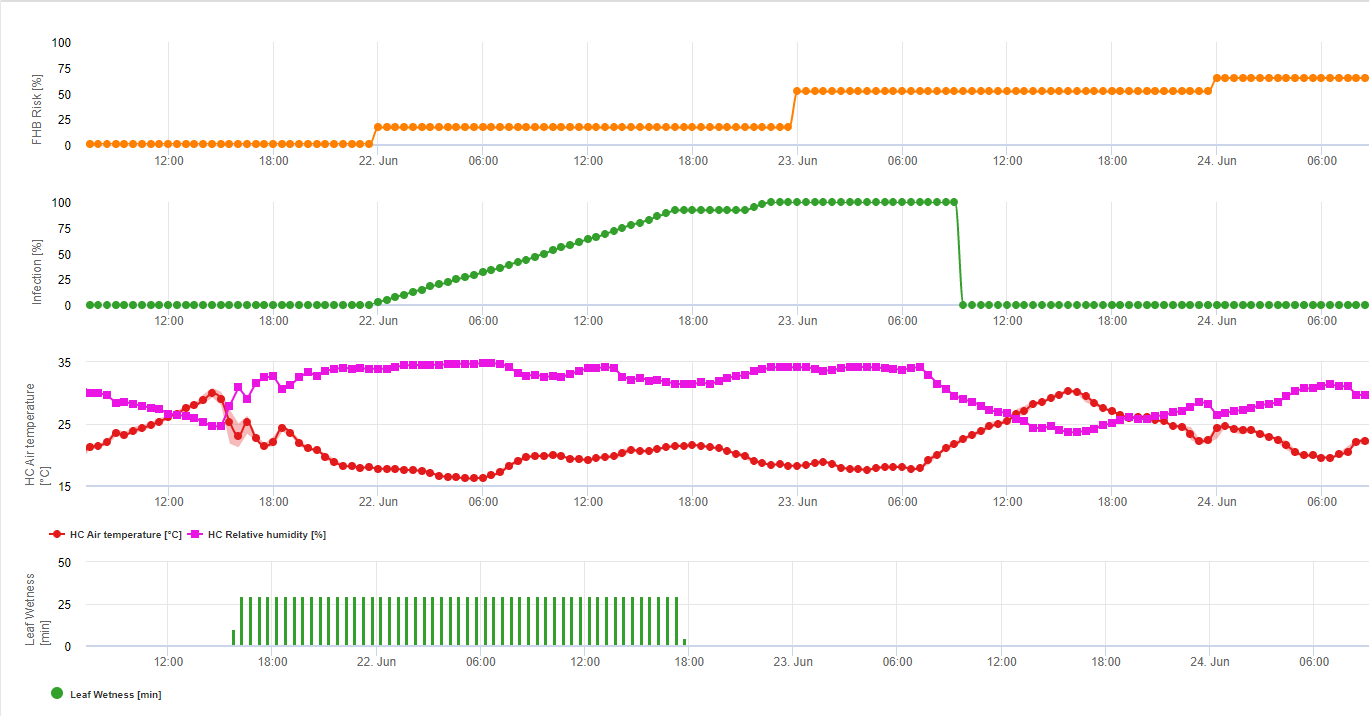
Resumiendo todas las diferentes combinaciones de temperatura y humedad que hemos encontrado en la literatura, hemos decidido señalar las infecciones por Fusarium Head bligh si la temperatura y el periodo de humedad de la hoja o periodos con más de 85% de humedad relativa superan los valores visualizados en el siguiente gráfico.
Las infecciones se inician con una lluvia de 2 mm. Se puede suponer que existe una infección por Fusarium Head Blight si el valor de progreso de la infección alcanza 100%. El cálculo de este valor de progreso sigue la relación entre la duración de las condiciones húmedas y la temperatura mostrada en el gráfico anterior.
Este modelo se utiliza para visualizar los días de infección y las condiciones climáticas durante el clima. El conocimiento por parte de los cultivadores del estado de desarrollo de las distintas variedades de trigo permite decidir la aplicación de una pulverización curativa inmediatamente después de la infección.
Alerta por micotoxinas de fusarium
Los ensayos de infección con periodos prolongados de humectación de las hojas de Fusarium head blight dan lugar a contenidos elevados de micotoxinas. A partir de esta información, se asume que un periodo de humectación de la hoja de 48 horas o más durante los estadios 61 y 69 supone un alto riesgo de micotoxinas. Las experiencias realizadas a lo largo de los análisis de DON en trigo cultivado comercialmente mostraron que los periodos de humedad de la hoja suficientemente largos para la infección después de una infección inicial en las fases 61 a 69 pueden aumentar también los valores de DON. En el caso de periodos de humedad foliar más largos, las micotoxinas pueden aumentar hasta el estadio 85. FieldClimate acumula una cifra de riesgo proporcional al progreso de la infección por cada periodo de infección exitoso durante el periodo que se ha seleccionado como adecuado para este cálculo. 6 periodos de infección acabados de completar supondrían un riesgo de 100%. Normalmente, el periodo de humedad de la hoja que conduce a una infección por fusarium es superior al mínimo necesario. Por lo tanto, la mayoría de las infecciones por fusarium provocarán un aumento del riesgo superior a 17%. El valor de riesgo que apunta a una situación problemática de micotoxinas depende de la historia del campo. El trigo cultivado después de maíz sin laboreo o el trigo sin laboreo después de maíz sin laboreo sólo puede conllevar un pequeño riesgo si no se pulveriza en la situación óptima. En el trigo no rociado tenemos que esperar un aumento de los valores de DON ya después de 35% de riesgo. El trigo después del trigo sin laboreo que sigue a cualquier otro cultivo que no sea maíz o trigo puede conllevar un riesgo mayor de 50%. Si tenemos trigo después de maíz o trigo con labranza el riesgo que se acsted puede extenderse a 70%. El trigo de primer año debe someterse a pruebas de DON si el riesgo supera los 100%.
1) Fusarium Head Blight Modelo de riesgo que calcula los periodos de riesgo de infección. Cuando se alcanza la infección 100% (línea verde), el riesgo (línea azul) es muy alto y las condiciones para el hongo han sido favorables para la infección. En función del método de aplicación (curativo, preventivo), la línea azul indica el periodo de riesgo.
2) Tizón de la cabeza por Fusarium: en este modelo la infección del FHB se calcula por la precipitación (se necesitan 2mm), la humedad relativa (por encima de 85%) o la humedad de la hoja, la temperatura durante el proceso. Si la infección alcanza los 100%, se habrán alcanzado las condiciones óptimas para el hongo patógeno. A continuación, el modelo calcula el riesgo de micotoxinas del FHB.
Existen dos enfermedades principales de Septoria en el trigo. Se trata de la mancha Septoria tritici, provocada por el hongo Septoria tritici (teleomorfo: Mycophaerella graminicola), y Septoriosis nodular causada por el hongo Septoria nodorum (teleomorfo: Leptosphaeria nodorum). Ambas enfermedades causan graves pérdidas de rendimiento que oscilan entre el 31 y el 53% (Eyal, 1981; Babadoost y Herbert, 1984; Polley y Thomas, 1991). En todo el mundo están afectadas más de 50 millones de hectáreas de trigo, que crecen principalmente en zonas de alta pluviosidad. Durante los últimos 25 años, estas enfermedades han ido en aumento y se han convertido en un importante factor limitante de la producción de trigo en determinadas zonas. Cuando las epidemias son graves, los granos de los cultivares de trigo susceptibles se marchitan y no son aptos para la molienda. Las epidemias de Septoria tritici blotch y Septoria nodorum blotch del trigo están asociadas a condiciones climáticas favorables (lluvias frecuentes y temperaturas moderadas), prácticas culturales específicas, disponibilidad de inóculo y presencia de cultivares de trigo susceptibles (Eyal et al., 1987).
Septoria spp. Biología
Después de Erick De Wolf, Septoria Tritici Blotch, Kansas State University, abril de 2008 Septoria tritici mancha conocida como mancha moteada de la hoja, está causada por el hongo Septoria tritici. Se distribuye por todas las zonas trigueras del mundo y constituye un grave problema en muchas regiones. La mancha de Septoria tritici es más dañina cuando la enfermedad ataca las hojas superiores y las cabezas de las variedades susceptibles a finales de la temporada.
Síntomas
Septoria tritici Los síntomas de la mancha aparecen por primera vez en otoño. Los síntomas iniciales son pequeñas manchas amarillas en las hojas. Estas lesiones suelen adquirir un color bronceado claro a medida que envejecen, y los cuerpos fructíferos del hongo pueden verse incrustados en las lesiones de las axilas. Las lesiones tienen forma irregular y varían de elípticas a largas y estrechas (Figura 1). Las lesiones contienen pequeñas motas redondas negras que son los cuerpos fructíferos del hongo. Los cuerpos fructíferos negros parecen granos de pimienta negra y normalmente pueden verse sin ayuda de una lupa. La enfermedad comienza en las hojas inferiores y avanza gradualmente hasta la hoja bandera. Las vainas de las hojas también son susceptibles de ser atacadas. En años húmedos, el hongo de la mancha moteada de la hoja puede desplazarse a los cogollos y causar lesiones marrones en las glumas y las axilas, conocidas como mancha de las glumas. Estas lesiones a menudo se vuelven de color bronceado claro a medida que envejecen y los cuerpos fructíferos del hongo se ven a menudo incrustados en las lesiones de las glumas.
La fase de la mancha de la gluma puede causar una pérdida significativa de rendimiento, pero no se conoce bien la relación entre la gravedad de la enfermedad y la pérdida de rendimiento. Septoria tritici puede confundirse con otras enfermedades de las hojas que presentan síntomas muy similares: mancha bronceada y Stagonspora nodorum por ejemplo. Es frecuente que las plantas estén infectadas por más de una de estas enfermedades foliares, y puede ser necesario realizar exámenes de laboratorio para diagnosticar con precisión qué enfermedades son las más prevalentes. El examen de laboratorio es casi siempre necesario para distinguir la causa de la mancha de la gluma. Conocer la especie no es importante para las decisiones de pulverización porque las tres enfermedades responden de forma similar a los fungicidas. Sin embargo, saber cuáles son las enfermedades más prevalentes es una parte importante de la selección de variedades, ya que diferentes genes controlan la resistencia a las enfermedades.
La forma más fiable de distinguir la mancha por Septoria tritici de las otras enfermedades es por la presencia de los cuerpos fructíferos negros del hongo. El hongo que causa la mancha bronceada no produce este tipo de estructura reproductiva. Sin embargo, en condiciones húmedas, el hongo que causa la mancha Stagonospora nodorum producirá cuerpos fructíferos de color marrón claro. Además de la diferencia de color, estas estructuras también son más pequeñas que las producidas por Septoria tritici.
Ciclo de vida
Septoria tritici sobrevive durante el verano en los residuos de un cultivo de trigo anterior e inicia las infecciones en otoño. Existen algunas pruebas de que el hongo es capaz de sobrevivir en asociación con otras gramíneas hospedadoras y semillas de trigo. Estas fuentes del hongo son probablemente más importantes cuando no hay residuos de trigo. Independientemente de las prácticas de rotación o de gestión de residuos, suele haber suficiente inóculo para iniciar las infecciones otoñales. Septoria tritici La mancha se ve favorecida por el clima fresco y húmedo. El intervalo óptimo de temperatura es de 16 a 21 °C; sin embargo, pueden producirse infecciones durante los meses de invierno a temperaturas tan bajas como 5 °C. La infección requiere al menos 6 horas de humedad en las hojas, y hasta 48 horas de humedad para una infección máxima. Una vez que se ha producido la infección, el hongo tarda entre 21 y 28 días en desarrollar los característicos cuerpos fructíferos negros y producir una nueva generación de esporas. Las esporas producidas en estos cuerpos fructíferos se exudan en masas pegajosas y requieren lluvia para salpicarlas sobre las hojas superiores y los cogollos.
Infección por Septoria tritici
Picnidiosporas de S. tritici germinan en agua libre de ambos extremos de la espora o de las células intercalares (Weber, 1922). La germinación de las esporas no comienza hasta unas 12 horas después del contacto con la hoja. Los tubos germinativos crecen al azar sobre la superficie de la hoja. Weber (1922) sólo observó la penetración directa entre las células epidérmicas, pero otros concluyeron que la penetración a través de los estomas abiertos y cerrados es el principal medio de penetración en el hospedador (Benedict, 1971; Cohen y Eyal, 1993; Hilu y Bever, 1957). Kema et al. (1996) sólo observaron penetración estomática. Las hifas que crecen a través de los estomas se contraen hasta alcanzar un diámetro de aproximadamente 1 μm, y luego se ensanchan tras alcanzar la cavidad subestomática.
Las hifas crecen paralelas a la superficie de la hoja por debajo de las células epidérmicas, después a través del mesófilo hasta las células de la parte inferior de la epidermis, pero no dentro de la epidermis. No se forman haustorios y el crecimiento hifal está limitado por las células del esclerénquima alrededor de los haces vasculares, excepto cuando las hifas son muy densas. Los haces vasculares no son invadidos. Las hifas crecen intercelularmente a lo largo de las paredes celulares a través del mesófilo, ramificándose en el tabique o en el centro de una célula. No aparecen síntomas macroscópicos durante unos 9 días, salvo alguna célula muerta ocasional, pero las células del mesófilo mueren rápidamente a los 11 días. Los picnidios se desarrollan en cámaras substomáticas. Las hifas rara vez crecen dentro de las células huésped (Hilu y Bever, 1957; Kema et al, 1996; Weber, 1922).
La infección exitosa sólo se produce después de al menos 20 horas de alta humedad. Sólo se desarrollan unas pocas motas marrones si las hojas permanecen húmedas durante 5-10 horas después de la deposición de las esporas (Holmes y Colhoun, 1974) o hasta 24 horas (Kema et al., 1996). Las relaciones huésped-parásito son las mismas en trigos resistentes o susceptibles. La germinación de las esporas en la superficie de la hoja es la misma independientemente de la susceptibilidad. El número de penetraciones exitosas es aproximadamente el mismo, pero el crecimiento hifal es más rápido en cultivares susceptibles, resultando en más lesiones. Las hifas se extienden más allá de la zona necrótica en todos los cultivares. Una toxina puede jugar un papel en la patogénesis (Cohen y Eyal, 1993; Hilu y Bever, 1957). Por el contrario, la colonización se redujo considerablemente en una línea resistente (Kema et al., 1996).
Stagonospora (Septoria) y Septoria patógenos de los cereales: El proceso de infección
B.M. Cunfer, Departamento de Patología Vegetal, Universidad de Georgia, Griffin, GA
El proceso de infección se ha estudiado más intensamente para Stagonospora (Septoria) nodorum y Septoria tritici. Un estudio en profundidad sobre Septoria passerinii está disponible. Casi toda la información comunicada se refiere a la infección por picnidiosporas. Sin embargo, el proceso de infección de otras formas de esporas es bastante similar. La información presentada se refiere principalmente a la infección de hojas en condiciones óptimas. Algunos estudios se realizaron con plantas de semillero intactas, mientras que otros se llevaron a cabo con hojas desprendidas. Infección del coleóptilo y la plántula de trigo por S. nodorum fue descrito en detalle por Baker (1971) y revisado por Cunfer (1983). Aunque no se han hecho comparaciones precisas, parece que el proceso de infección tiene muchas similitudes en cada sistema huésped-parásito y es típico de muchos patógenos necrótrofos. Se excluye la información sobre los factores que influyen en el desarrollo de los síntomas y la expresión de la enfermedad, pero han sido revisados por otros autores (Eyal et al., 1987; King et al., 1983; Shipton et al., 1971). Se incluye un resumen de los factores que afectan a la longevidad de las esporas en la superficie de las hojas.
Papel de los cirros y supervivencia de las esporas en la superficie de la hoja La información más detallada sobre la función del cirro que recubre las picnidiosporas exudadas del picnidio corresponde a S. nodorum. El cirro es un gel formado por compuestos proteínicos y sacáridos. Su composición y función son similares a las de otros hongos de la familia Sphaeropsidales (Fournet, 1969; Fournet et al., 1970; Griffiths y Peverett, 1980). Las funciones principales de los componentes de los cirros son la protección de las picnidiosporas frente a la desecación y la prevención de la germinación prematura.
El cirro protege las picnidiosporas de modo que algunas permanecen viables al menos 28 días (Fournet, 1969). Cuando el cirro se diluía con agua, si la concentración de la solución de cirro era >20%, germinaban menos de 10% de picnidiosporas. A menor concentración, los componentes proporcionan nutrientes que estimulan la germinación de las esporas y la elongación de los tubos germinativos. La longitud de los tubos germinativos aumentó hasta una concentración de cirros de 15%, y luego disminuyó moderadamente a concentraciones más altas (Harrower, 1976). Brennan et al. (1986) informaron de una mayor germinación en cirros diluidos. Los componentes de los cirros redujeron la germinación a una humedad relativa de 10-60%. Una vez que las esporas se dispersan, los efectos estimulantes del fluido de cirros son probablemente insignificantes (Griffiths y Peverett, 1980).
A una humedad relativa de 35-45%, las esporas de S. tritici en cirros permanecieron viables al menos 60 días (Gough y Lee, 1985). Los componentes de los cirros pueden actuar como inhibidores de la germinación de las esporas, o el alto potencial osmótico de los cirros puede impedir la germinación. Las picnidiosporas de S. nodorum no sobrevivieron durante 24 horas a una humedad relativa superior a 80% a 20 C. Las esporas sobrevivieron dos semanas o más a una humedad relativa <10% (Griffiths y Peverett, 1980). Cuando el fluido cirroso de S. nodorum se diluyó con agua, aproximadamente dos tercios de las picnidiosporas perdieron viabilidad en 8 horas, y después de 30 horas a la luz del día, sólo 5% germinaron. Cuando las esporas se almacenaron en la oscuridad, 40% siguieron siendo viables después de 30 horas (Brennan et al., 1986).
Conidios secos de S. nodorum, a la sombra y a la luz solar directa, sobrevivieron a la intemperie al menos 56 horas (Fernandes y Hendrix, 1986a). La germinación de las picnidiosporas de S. nodorum fue inhibida por la UV-B continua (280-320 nm), mientras que la germinación de las de S. tritici no lo fue. La extensión del tubo germinativo bajo UV-B continua fue inhibida para ambos hongos, en comparación con la oscuridad (Rasanayagam et al., 1995).
Infección por Septoria nodorum
El proceso de penetración en el huésped y el desarrollo de S. nodorum dentro de la hoja fue examinada en detalle por varios investigadores (Baker y Smith, 1978, Bird y Ride 1981, Karjalainen y Lounatmaa, 1986; Keon y Hargreaves, 1984; Straley, 1979; Weber, 1922). Las picnidiosporas tienden a alojarse en las depresiones entre dos células epidérmicas, y muchos intentos de penetración en las hojas comienzan allí. Las esporas germinan en la superficie de la hoja en respuesta a la humedad libre (Fernandes y Hendrix, 1986b). Comienzan a germinar 2-3 horas después de la deposición, y después de 8 horas la germinación puede alcanzar 90%. La penetración en la hoja comienza unas 10 horas después de la deposición de las esporas (Bird y Ride, 1981; Brönnimann et al., 1972; Holmes y Colhoun, 1974).
Al inicio de la germinación, el tubo germinativo está rodeado de un material amorfo que se adhiere a la hoja. Los tubos germinativos que crecen desde cualquiera de los extremos de una espora y desde las células intercalares tienden a crecer a lo largo de las depresiones entre las células y a menudo se orientan a lo largo del eje largo de la hoja (O'Reilly y Downes, 1986). Las hifas de las esporas que no están en las depresiones crecen al azar con ramificaciones ocasionales (Straley, 1979). Se forma un apresorio con una púa de infección que penetra la cutícula y las paredes periclinales de las células epidérmicas directamente en el lumen celular, lo que provoca la muerte rápida de las células.
Muchas penetraciones son primero subcuticulares o el crecimiento lateral de una hifa ocurre dentro de la pared celular antes del crecimiento hacia el citoplasma (Bird y Ride, 1981; O'Reilly y Downes, 1986). La penetración a través de estomas abiertos y cerrados también se produce y puede ser más rápida que la penetración directa (Harrower, 1976; Jenkins, 1978; O'Reilly y Downes, 1986; Straley, 1979). Los tubos germinativos se ramifican en los estomas y en las uniones de las células epidérmicas. La penetración de un tubo germinativo en un estoma puede producirse sin formación de un apresorio. En ocasiones, la penetración se produce a través de los tricomas (Straley, 1979). Aparentemente, la mayoría de los intentos de penetración fracasan, formándose densas papilas en las células en el lugar del intento de penetración (Karjalainen y Lounatmaa, 1986; Bird y Ride, 1981).
Tras la penetración, las células epidérmicas mueren rápidamente y se lignifican, y las hifas crecen hacia el mesófilo. Las células del mesófilo se deforman y el material lignificado se deposita en el exterior de algunas células, que se colapsan. La lignificación se produce antes de que las hifas alcancen la célula. El proceso es el mismo en todos los cultivares, pero se desarrolla más lentamente en los cultivares resistentes. Las hifas crecen intercelularmente entre las células epidérmicas y después en el mesófilo. Cuando penetran en el mesófilo, el deterioro del cloroplasto comienza en 6-9 días (Karjalainen y Lounatmaa, 1986).
Sin embargo, la tasa fotosintética empieza a disminuir un día después de la infección y antes de que los síntomas sean visibles (Krupinsky et al, 1973). El tejido esclerénquima alrededor de los haces vasculares impide la infección del tejido vascular. Los haces vasculares bloquean la propagación de las hifas a través del mesófilo, excepto cuando el tejido esclerénquima es joven y no está completamente formado (Baker y Smith, 1978).
Stagonospora nodorum libera una amplia gama de enzimas que degradan la pared celular, como amilasa, pectina metil esterasa, poligalacturonasas, xilanasas y celulasa in vitro y durante la infección de hojas de trigo (Baker, 1969; Lehtinen, 1993; Magro, 1984). La información relacionada con la degradación de la pared celular por las enzimas concuerda con las observaciones histológicas. Estas enzimas pueden actuar conjuntamente con las toxinas. La sensibilidad enzimática puede estar relacionada con la resistencia y la tasa de colonización fúngica (Magro, 1984). Como muchos necrótrofos, los patógenos Septoria y Stagonospora producen compuestos fitotóxicos in vitro. El deterioro celular y la muerte antes del crecimiento hifal en el tejido mesófilo (Bird y Ride, 1981) es consistente con la producción de toxinas. Sin embargo, no se ha establecido un papel definitivo para las toxinas en el proceso de infección y su relación con la resistencia del huésped (Bethenod et al, 1982; Bousquet et al, 1980; Essad y Bousquet, 1981; King et al, 1983). Las diferencias en el rango de hospedadores entre las cepas de trigo y cebada adaptadas de S. nodorum puede estar relacionada con la producción de toxinas (Bousquet y Kollmann, 1998). El inicio de la germinación de las esporas y el porcentaje de esporas germinadas no están influidos por la susceptibilidad del hospedador (Bird y Ride, 1981; Morgan 1974; Straley, 1979; Straley y Scharen, 1979; Baker y Smith, 1978).
Bird y Ride (1981) informaron que la extensión de los tubos germinativos en la superficie de la hoja era más lenta en los cultivares resistentes que en los susceptibles. Este mecanismo, expresado al menos 48 horas después de la deposición de las esporas, indica una resistencia a la elongación de los tubos germinativos previa a la penetración. Hubo menos penetraciones exitosas en cultivares resistentes, y la penetración procedió más lentamente en cultivares resistentes (Baker y Smith, 1978; Bird y Ride, 1981). Se propuso que la lignificación limitaba la infección tanto en cultivares resistentes como susceptibles, pero otros factores ralentizaban el desarrollo del hongo en las líneas resistentes. En las líneas susceptibles, las hifas de crecimiento más rápido pueden escapar a la lignificación de las células huésped.Cuatro días después de la inoculación de cebada con un aislado de biotipo de trigo de S. nodorum, las hifas crecían a través de la cutícula y a veces en las capas externas de celulosa de las paredes celulares epidérmicas. Debajo de las hifas de penetración se depositaban papilas gruesas que no penetraban en las células (Keon y Hargreaves, 1984).
Infección por Septoria passerinii
Green y Dickson (1957) presentan una descripción detallada del proceso de infección de S. passerinii en la cebada. El proceso de infección es similar al S. tritici. Al igual que S. tritici, el tiempo necesario para la penetración en las hojas es considerablemente mayor que el de S. tritici. S. nodorum. Los tubos germinativos se ramifican y crecen sobre la superficie de la hoja al azar, pero a veces a lo largo de depresiones entre células epidérmicas. La penetración en la hoja se produce casi exclusivamente a través de los estomas. Las hifas germinativas se hinchan y, si la penetración no tiene éxito, las hifas siguen alargándose. No se produce penetración 48 horas después de la deposición de las esporas. Después de 72 horas, los tubos germinativos se engrosan sobre los estomas, crecen entre las células protectoras y en las caras urfales de las células accesorias y en las cavidades substomáticas. La penetración directa entre las células epidérmicas sólo se observa en raras ocasiones.
La germinación de las esporas y la penetración en el huésped son las mismas en los cultivares resistentes y susceptibles. La extensión de las hifas dentro de las hojas es mucho menor en los cultivares resistentes y se observan papilas en muchas paredes celulares, pero no en todas. Las hifas crecen bajo la epidermis de un estoma a otro, pero no penetran entre las células epidérmicas. El mesófilo está colonizado, pero no se forman haustorios. Cuando las células del mesófilo se necrosan, las células epidérmicas se colapsan. El desarrollo micelial en la hoja es escaso y suele estar bloqueado por haces vasculares. En las hojas más jóvenes, si la vaina vascular está menos desarrollada, las hifas pasan entre el haz y la epidermis. Los picnidios se forman en cavidades substomáticas, principalmente en la superficie superior de la hoja (Green y Dickson, 1957).
Factores que afectan a la longevidad de las esporas en la superficie de la hoja Entre los patógenos Stagonospora y Septoria de los cereales, sólo se dispone de información definitiva sobre el proceso de infección en el caso de S. nodorum, S. triticiy S. passerinii. Al igual que muchos otros patógenos necrótrofos, ninguno de los dos grupos de patógenos provoca la reacción de hipersensibilidad. Una diferencia significativa en el proceso de infección entre los patógenos Septoria y Stagonospora es que la germinación y penetración de las esporas es mucho más rápida en el caso de Septoria y Stagonospora. S. nodorum que para S. tritici y S. passerinii. Esto influye considerablemente en la epidemiología de las enfermedades.
Los patógenos de Septoria penetran en la planta principalmente a través de los estomas, mientras que S. nodorum penetra tanto directamente como a través de los estomas. S. nodorum penetra y mata rápidamente las células epidérmicas, pero S. tritici y S. passerinii no matan las células epidérmicas hasta que las hifas se han ramificado a través del mesófilo de la hoja y comienza una rápida necrosis. Los estudios histológicos del crecimiento del hongo tras la penetración en el hospedador coinciden con los datos generados por los estudios epidemiológicos de la resistencia del hospedador. La resistencia ralentiza la tasa de colonización del hospedador, pero no tiene un efecto apreciable en el proceso de desarrollo de la lesión.
Los mecanismos que controlan la respuesta del hospedador, ya estén relacionados con enzimas y toxinas u otros metabolitos liberados por los patógenos durante la infección, aún no están claros. Existe poca información sobre la infección por ascosporas. El proceso de infección es probablemente muy similar al de las picnidiosporas. Las ascosporas de Phaeosphaeria nodorum germinan en un amplio rango de temperaturas y sus tubos germinativos penetran directamente en la hoja. Sin embargo, según Rapilly et al. (1973), las ascosporas, a diferencia de las picnidiosporas, no germinan en agua libre.
Modelo de infección por Septoria spp.
Las infecciones por Septoria son posibles a bajas temperaturas, mientras que las temperaturas inferiores a 7°C pueden no provocar una infección en un plazo de 2 días. La temperatura óptima de la enfermedad se alcanza en la zona de 16 a 21°C. Las infecciones son posibles en un periodo de alta humedad relativa o de humedad foliar de 14 horas o más. Para cumplir las condiciones hemos decidido separar en modelos las infecciones débiles, moderadas y graves. Las infecciones débiles pueden darse si es posible que el patógeno infecte el tejido del huésped. Esto significa que las infecciones débiles pueden tener lugar si las temperaturas son mínimas y los periodos de humedad de las hojas tienen una duración crítica. Una infección moderada tendrá lugar en condiciones en las que la mayoría de los ensayos de infecitón conducen a resultados razonables y las infecciones graves tienen lugar en condiciones en las que el patógeno dispone de condiciones óptimas para la infección.
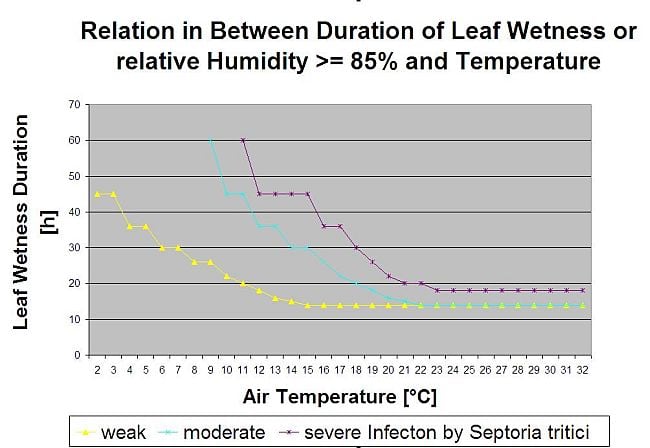

Comienza la infección tras una lluvia de 0,5 mm. Decidimos no utilizar un modelo para la formación de picnidios. Se supone que la condición necesaria para la formación de picnidios es un periodo con una humedad relativa superior a 85%. Se espera que el tiempo de vida de los picnidios sea de 24 horas. En todos los climas en los que Septoria tritici tiene posibilidades de infectar, encontraremos 2 horas cumpliendo estas condiciones casi todos los días alrededor de la salida del sol.
Evaluación de la gravedad de la infección
Para poder evaluar la presión de la infección por Septoria tritici entre el estadio 10 (primera hoja a través del coleóptilo) y el estadio 32 (segundo nudo al menos 2 cm por encima del primer nudo) y entre el 32 y el 51 (inicio del espigado), debemos evaluar la gravedad de las infecciones en función de las condiciones climáticas. Esta evaluación se realiza en una escala de 1 a 5. Se da una gravedad de 1 si la infección es grave. Se da una severidad de 1 si se cumplen las condiciones para una infección débil y ha llovido menos de 5 mm, de lo contrario el valor de severidad correspondiente será 2. Se da una severidad de 3 si se cumplen las condiciones para una infección débil y ha llovido menos de 5 mm, de lo contrario el valor de severidad correspondiente será 2. La gravedad será 3 si se cumple la condición de infección moderada y ha llovido menos de 5 mm. Si ha llovido más de 5 mm durante una infección moderada o menos de 5 mm durante una infección grave, la gravedad será 4.
Una infección grave con más de 5 mm de lluvia corresponde a un valor de gravedad de 5.
Septoria tritici evaluación de la presión de la enfermedad
El clima es sólo uno de los factores que determinan la presión de la enfermedad en el campo. Los otros dos factores son el historial del campo y la susceptibilidad de la variedad cultivada. Si podemos acumular los valores de severidad de la enfermedad desde el estadio 10 al 32 hasta un valor de 4, podemos esperar una presión débil de la enfermedad por el clima. Si este valor alcanza 6 podemos esperar una presión moderada de la enfermedad y si alcanza 10 podemos esperar una alta presión de la enfermedad por el clima. Conocer la susceptibilidad de la variedad y el historial del campo nos llevará a pulverizar o no sobre una presión de enfermedad débil o moderada en esta situación. Tener un valor acumulado de 10 puede conducir a una pulverización en la fase 32 de todos modos. La decisión de pulverizar en una fase posterior depende más del clima primaveral. Si somos capaces de acumular los valores de severidad desde el estadio 10 hasta un valor de 6, podemos esperar una presión débil de la enfermedad. Si este valor alcanza 10 podemos esperar una presión moderada de la enfermedad y si este valor alcanza 15 podemos esperar una alta presión de la enfermedad debido a la situación climática.
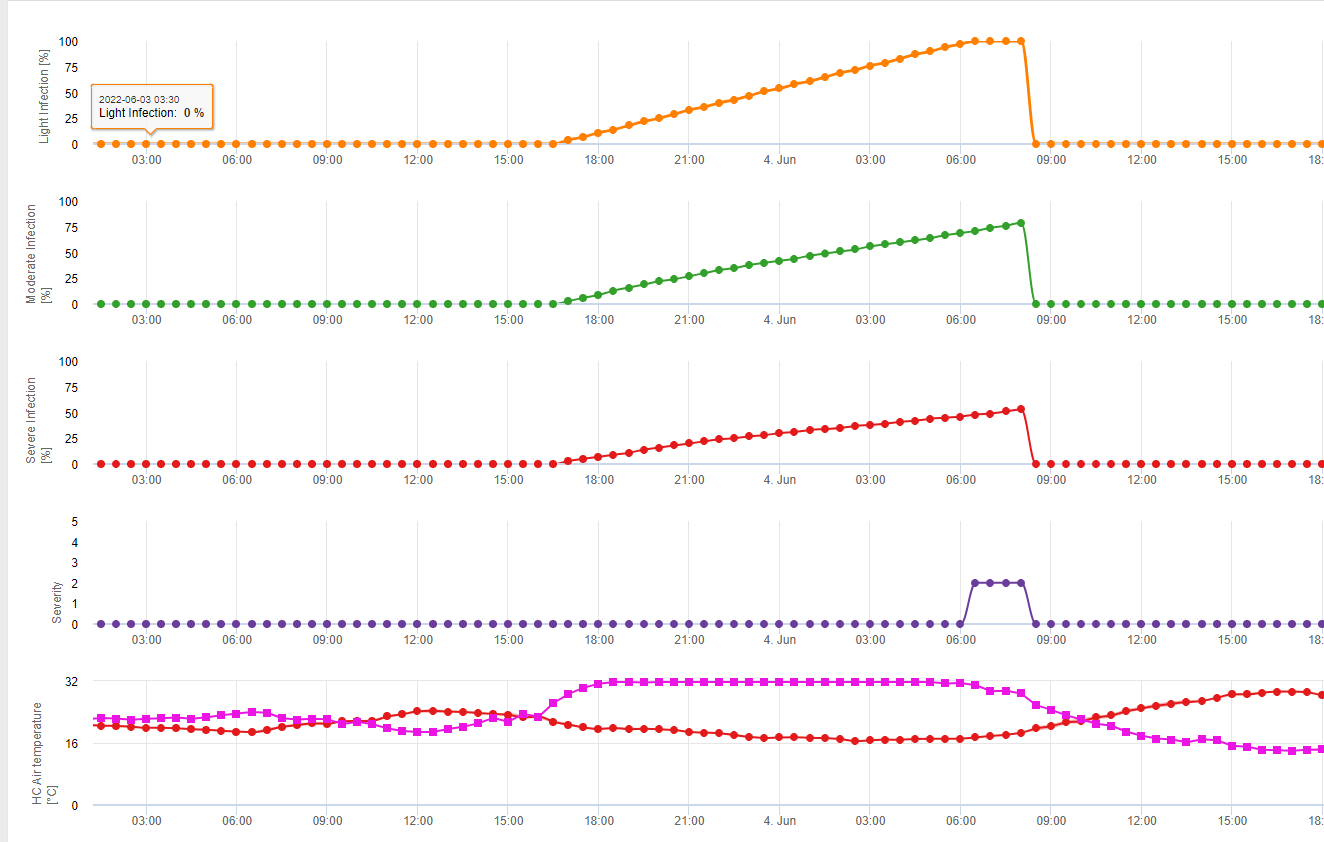
En FieldClimate mostramos el Septoria tritici Severidad junto con las tres diferentes severidades de infección en un gráfico (véase arriba). Debido a las precipitaciones y a los largos periodos de humedad de las hojas, se dan las condiciones para una infección grave por S. tritici han sido campo lleno los días 14 y 16 de mayo. Los niveles de gravedad alcanzan el valor más alto de 5 el 14 de mayo, lo que significa que ahora hay un alto riesgo de infección.
La biología de la infección por Stagonospora nodorum difiere en cierta medida de la de S. tritici, pero esta diferencia no es lo suficientemente grande como para crear un modelo independiente. Por lo tanto, sugerimos utilizar este modelo para todo el complejo de enfermedades causadas por Stagnospora y Septoria en cereales, incluyendo S. passerinii. S. tritici y S. passerinii tienden a necesitar periodos de humectación de la hoja más largos que S. nodorum. En zonas con una alta presión de S. nodorum Las infecciones clasificadas como débiles, con un valor de gravedad de 2, deben tratarse con más seriedad que en otros ámbitos.
Para Septoriosis nodular En FieldClimate se muestra un modelo de riesgo (véase más arriba). Se determinó un riesgo elevado el 17 de junio y el 7 de julio (100%). Dependiendo de la fase de la planta susceptible a la infección, deben tenerse en cuenta las medidas fitosanitarias si el riesgo alcanza los 80% (véase también la previsión meteorológica, protección fitosanitaria). Si el riesgo es de 100% y ya se ha detectado una infección, deben tomarse medidas fitosanitarias sistémicas (aplicación curativa) para proteger la planta.