Biología de Plasmopara viticola (Mildiú velloso)
P. viticola es un parásito obligado. Lo que significa que necesita órganos verdes y frescos de la vid para crecer. Durante el periodo libre de vegetación persiste formando cuerpos fructíferos, las llamadas oosporas. Las oosporas de los oomicetos pueden sobrevivir períodos muy largos en el suelo. Por ello, podemos encontrar mildiu velloso en lugares donde no es posible que se produzcan infecciones todos los años. En primavera, cuando la capa superior del suelo está lo suficientemente húmeda y caliente, las oosporas forman los llamados macrosporangios, que pueden liberar hasta 200 zoosporas en el agua libre. Las zoosporas se desplazan hasta las hojas y los racimos por el viento en gotas de agua. Tienen dos flagelos y se mueven en una película de agua en el envés de las hojas o los racimos y bayas jóvenes para encontrar un estoma y entrar en el tejido vegetal. Entran y germinan en el estoma, en el que transfieren todo su plasma en menos de una hora. En estudios microscópicos, el hallazgo del estoma, el enquistamiento y la germinación en el estoma finalizaron en 90 minutos.
P. viticola crece en el espacio intercelular y se alimenta de haustorias que penetran en las células epidérmicas y parenquimatosas. En función de la temperatura y la humedad relativa desarrolla suficiente crecimiento intercelular con suficientes haustorias para formar un cuerpo subestomático que llena toda la zona subestomática y que levanta el tejido epidérmico del tejido parenquimatoso. Esto da lugar al síntoma visible de la mancha de aceite.
Los oomicetos son esporulando en ausencia de luz cuando la humedad relativa es muy alto. En P. viticola no hay esporulación si las temperaturas son inferiores a 12°C y la humedad relativa es inferior a 95%. Los esporangióforos están formados por las vesículas substomatélicas y saldrán del estoma. Los esporangios recién formados son pegajosos y sólo pueden desprenderse de los esporangióforos con agua. Cuando disminuye la humedad relativa, los esporangios se vuelven pegajosos y también pueden ser eliminados por el viento.
Los esporangios liberan hasta 20 zoosporas en el agua libre. Estas zoosporas también tienen que ser distribuidas por el viento en gotitas de agua para llegar a las hojas frescas, o los esporangios pueden ser distribuidos por la lluvia o el propio viento. El proceso de infección primaria y secundaria es el mismo.
Dada la gran importancia de la fase sexual para la hibernación del patógeno, podemos suponer que los tipos de apareamiento encajan en todos los viñedos en los que se da el mildiu de la vid. La formación de zoosporas tiene lugar en las hojas más viejas durante el verano y principios del otoño.
Infección primaria en detalle
Las infecciones procedentes de las oosporas se denominan infecciones primarias. Este término es engañoso, ya que pueden producirse varias infecciones primarias a principios del verano. En los eventos epidemiológicos, la infección primaria no desempeña un papel importante si hay suficientes manchas de aceite en los viñedos y el potencial de infección de las esporas de verano (esporangios) supera al de las oosporas.
Las oosporas invernantes se convierten en los llamados macrosporangios cuando hay suficiente humedad relativa durante unas 24 horas. Los macrosporangios liberan sus zoosporas en el agua libre y la lluvia intensa (como en una tormenta eléctrica). Las esporas son transportadas a las hojas de la vid y/o a los brotes verdes. Infecciones primarias, por tanto, necesitan más tiempo lluvioso periodos o varias tormentas sucesivas. La primera lluvia hace que las hojas caídas se saturen de agua y las fuertes lluvias de los días siguientes hacen que los macrosporangios liberen sus zoosporas, que alcanzan las hojas o los sarmientos de la vid. Para que la infección tenga éxito, es necesario un periodo de humectación de las hojas suficientemente largo para que las zoosporas alcancen el estoma de las hojas o los sarmientos y los infecten (la ilustración anterior muestra el ciclo de desarrollo del mildiú velloso de la vid (Plasmopara viticola).
El Modelo para la infección primaria de Mildiu velloso comprueba primero, si el clima es adecuado para el desarrollo de macrosporangios. Este es el caso siempre y cuando las hojas estén húmedas, o la humedad relativa después de la lluvia no caiga por debajo de 70%. Dependiendo de la temperatura, los macrosporangios pueden desarrollarse en un plazo de 16 a 24 horas. Si hay macrosporangios maduros, se mostrará gráficamente en la pantalla de la infección primaria de Mildiú velloso. Si hay macrosporangios, una lluvia fuerte puede propagar sus zoosporas. Una lluvia continua de 5 mm se interpreta como una lluvia fuerte y las zoosporas se esparcen, una infección primaria es entonces posible si la humedad de la hoja ha durado lo suficiente.
 Infecciones secundarias
Infecciones secundarias
 Infecciones secundarias
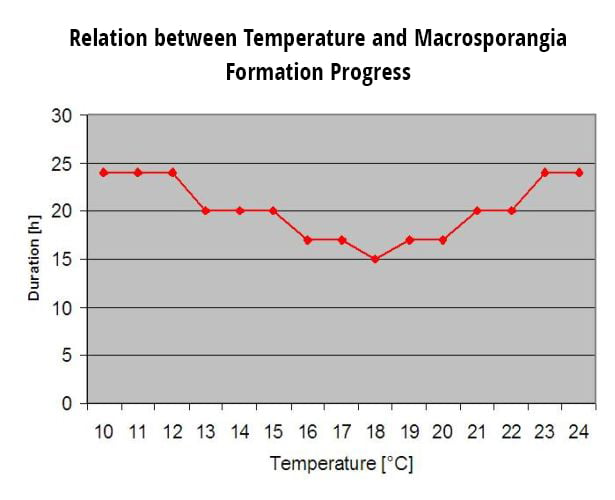
Infecciones secundariasInfecciones secundarias de Plasmopara viticola sólo son posibles si ya existen manchas de aceite maduras en su viñedo o cerca de él. Las manchas de aceite son maduras cuando son capaces de esporular (producir esporangios). Los esporangios sólo se producen noche. La luz solar inhibe la esporulación. Los esporangios se producen si más de 12°C y el humedad relativa es muy elevada. La tasa de producción de esporangios aumenta con la temperatura hasta 24°C. La temperatura óptima para la esporulación en las variedades de uva europeas (Vitis vinifera) es de unos 24°C. Si las temperaturas superan los 29°C, no puede producirse esporulación. En nuestro modelo comprobamos si humedad superior a 95% se produce durante la noche. Si esta condición se prolonga durante una temperatura horaria acumulada de más de 50°C, la esporulación finaliza y aparecen nuevos esporangios de Plasmopara existen en el viñedo. Por ejemplo, 50°C horas corresponden a 4 horas con 13°C o 3 horas con 17°C.
Esporangios de Plasmopara viticola tienen una vida útil limitada. Cuanto más calor hace y más seco está el aire, más rápido mueren (en ng.fieldclimate.com se pone a 0 cuando la h.r. es inferior a 50%). Mueren definitivamente en el siguiente periodo de rocío o de humedad de las hojas, que es demasiado corto para la infección. Por tanto, la aptitud de los esporangios más viejos es bastante limitada. Nuestro modelo supone que la vida de los esporangios se limita a un día.
Para causar nuevas infecciones, los esporangios deben distribuirse. Hay dos formas de distribuir las esporas: Si llueve inmediatamente después de la formación de los esporangios, éstos se propagan con las salpicaduras de lluvia. Si las hojas de la vid permanecen el tiempo suficiente mojadas, se produce una Plasmopara viticola se produce la infección. Si la mañana siguiente comienza sin lluvia y con humedad decreciente, los esporangios desecados se desprenden de sus ramas. Incluso un ligero movimiento de aire los llevará a las hojas sanas. A menos que llueva pronto, morirán.
Mildiú velloso Infección primaria
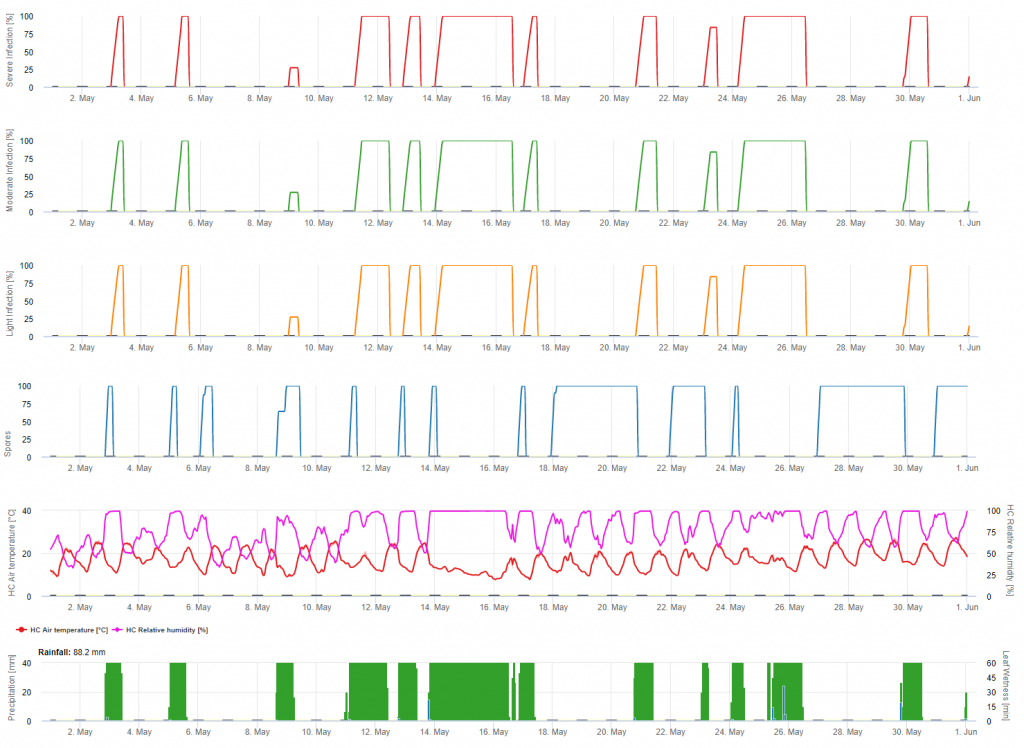
Las infecciones se muestran tras el desarrollo de las macrosporas (naranja, quinto gráfico) en tres clases de gravedad diferentes (débil: naranja, moderada: verde, grave: rojo) en función de la cantidad de lluvia y, por tanto, de la distribución de las esporas en el viñedo. Si se determina la infección por 100% se han dado las condiciones óptimas para entrar en el tejido vegetal (curativo fitosanitario) y la curva de incubación (primer gráfico) empieza a aumentar. Si la incubación termina (100%) y no se ha tenido en cuenta la protección fitosanitaria se deberían haber comprobado los síntomas (manchas de aceite) en el campo.
1. La curva de progreso de la infección y, tras la infección completa, el consiguiente periodo de incubación (tiempo transcurrido entre la infección y los síntomas visibles en el campo).
2. Se determina una curva de progreso de la infección cuando hay macrosporangios o esporangios y las hojas están húmedas. Aumenta con la humedad de las hojas y la temperatura. Si la curva de progreso de la infección alcanza 100%, se puede suponer que la infección se ha completado. La curva de progreso de la incubación que pertenece a la infección específica se empieza a calcular con el inicio de una infección. Si una infección no puede completarse, la curva de incubación también se detendrá.
3. Las condiciones meteorológicas se muestran en la misma página que los resultados del modelo de la enfermedad. Por lo tanto, puede comprobar de un vistazo la temperatura, la humedad relativa, la lluvia y la humedad de las hojas durante la infección.
4. El modelo señala las fechas de infección para las infecciones primarias y secundarias de mildiu de la vid. Esta es la información más crítica. Las infecciones, que aún no han sido cubiertas por fungicidas preventivos o curativos, pueden ser desastrosas para el rendimiento/la calidad. Además, la información sobre la gravedad de la infección (débil/moderada/grave) podría ser útil para decidir la estrategia fitosanitaria en función del historial del viñedo (cantidad de inóculo), la variedad y la experiencia de los años anteriores.
Estrategias fitosanitarias:
- Los fungicidas profilácticos y sistémicos o curativos se utilizan ampliamente para el control del mildiú velloso. Los productos químicos profilácticos se aplican antes, pero lo más cerca posible de un evento de infección en el período de mayor susceptibilidad del huésped, entre la longitud de los brotes de 10 cm y las bayas del tamaño de un guisante.
- También se pueden utilizar productos químicos profilácticos pulverizando lo más cerca posible antes de una previsión meteorológica de lluvia de más de 2 mm (para el círculo primario).
- El fungicida curativo debe aplicarse lo antes posible después de un evento de infección (infección 100%) y antes de que aparezcan las manchas de aceite. Tenga en cuenta la prevención del desarrollo de resistencia a los fungicidas.
- El seguimiento del viñedo para detectar la presencia de mildiu (manchas de aceite) debe hacerse con regularidad y teniendo en cuenta al menos 200 vides. Se considera que existe riesgo si se observan más de dos manchas por cada 50 cepas.
El modelo se basa en las publicaciones de MUELLER-THURGAU, ARENS, MUELLER y SLEUMER, BLAESER, HERZOG, GEHMANN y muchos otros trabajadores relacionados con la red de investigación sobre la epidemiología del mildiu y el oídio de la vid desde principios del siglo XIX.
Biología de Uncinula necator
El oídio de la vid está causado por el hongo Uncinula necator. Es un parásito obligado, lo que significa que necesita tejido verde del huésped para alimentarse. U. necator entró en Europa en el siglo XIX y se extendió por todos los países vitivinícolas hasta finales de este siglo. Tiene efectos muy fuertes sobre el rendimiento y puede provocar la defoliación de las vides si no hay un control químico. Todas las variedades de vid europeas son sensibles a este patógeno. La mejora de la resistencia a este patógeno es bastante reciente y los resultados requerirán más años de trabajo.
U. necator tiene que superar el periodo libre de vegetación en la mayoría de las zonas vitícolas. Para ello el patógeno utiliza dos mecanismos: a) sobrevivir en sus cuerpos fructíferos, las llamadas cleistotecias o b) sobrevivir también al invierno encapsulado en yemas latentes donde encuentra tejido vivo durante todo el invierno. La presencia de uno o de ambos modos de hibernación en un viñedo se basa en la historia de la enfermedad de este viñedo. Las yemas sólo pueden infectarse mientras no estén cerradas y rodeadas por un anillo de tejido senescente. Este es el caso sólo durante varias semanas después de la formación. En situación de viticultura ordinaria, las vides se podan en ramas con menos de 14 yemas o en espolones. Estos brotes restantes sólo pueden infectarse hasta que florecen. Por otro lado podemos encontrar brotes bandera del estadio BBCH 15 al 19. Los brotes bandera son brotes parcial o totalmente cubiertos de mildiu que se forman a partir de yemas infectadas latentes. Estos brotes bandera son una fuente de inóculo temprana y muy potente. En la mayoría de los casos no se distribuyen por el viñedo. Esto da lugar a manchas con una incidencia temprana y alta de la enfermedad en el interior de los viñedos y a manchas con una incidencia baja o moderada de la enfermedad al principio de la temporada. Los brotes bandera sólo representan un tipo de apareamiento debido a su historia vegetativa. Esto puede dar lugar a viñedos en los que encontremos pocas o ninguna cleistotecia debido a la dominancia de un único tipo de apareamiento.
Donde la cleistothecia es el único inóculo encontraremos una historia continua en la hibernación de cleistothecia que sólo puede cambiar a brotes bandera en años donde la enfermedad se acumula muy temprano. La formación de cleistothecia es bien visible debido a la coloración amarilla del tejido combinado sexual. Se puede encontrar en viñedos no fumigados a partir de mediados de verano. En los viñedos fumigados se necesita más tiempo para que la incidencia de la enfermedad aumente, lo que permite que los tipos de apareamiento se encuentren. La propagación de Uncinula necator durante el verano está influida principalmente por la temperatura.
Las temperaturas óptimas para el Mildiú Polvoroso se sitúan entre 21°C y 32°C. Cuanto mayor sea el número de horas diarias dentro de este intervalo de temperaturas, mayor será el riesgo de que aparezca el mildiú polvoroso. La formación de conidios, la infección y la velocidad de incubación necesitan rangos de temperatura similares. Debido al hecho de que U. necator no necesita agua libre para la infección ni una humedad relativa alta para la formación de conidios, no hay ningún impacto útil del parámetro de humedad directo sobre el patógeno.
La presencia de humedad libre interactúa con U. necator a través de su hiperparásito común Ampelomyces quisqualis. Este hiperparásito fúngico crece en el interior de las hifas del hongo oídio. Transforma los conidios y cleistotecios en picnidios propios. Necesita humedad libre o humedad relativa muy alta para infectar y crecer con hifas finas de crecimiento rápido en las hojas para encontrar lesiones de oídio. Muy infectado U. necator Las lesiones van adquiriendo un color marrón muy oscuro. Durante el periodo de lluvias, están cubiertas por un exudado de esporas blancas procedentes de los picnidios del hiperparásito. Ampelomyces quisqualis también está disponible como producto comercial para el control del mildiú polvoroso.
Trabajo práctico
Hemos aplicado diferentes modelos relativos al proceso de infección en FieldClimate.
a) el hongo pasa el invierno principalmente en forma de micelio. En primavera, el micelio empieza a crecer y forma conidióforos con conidios (por lo que no hay infección primaria).
b) si el hongo se desarrolla en otoño sobre las bayas, la forma sexual podría estar presente (formación de cleistotecios) y así, en primavera, la liberación de ascosporas podría conducir a la infección primaria (utilizar: modelo POLVOS/INFECCIÓN POR ASCOSPORAS).
Modelo de riesgo de California (por Thomas Gubler)
El Modelo de Evaluación del Riesgo de Mildiú Polvoroso de UC Davis (Modelo Gubler-Thomas) se basa en estudios epidemiológicos de laboratorio y de campo sobre el mildiú polvoroso de la vid en California.
Índice de evaluación de riesgos:
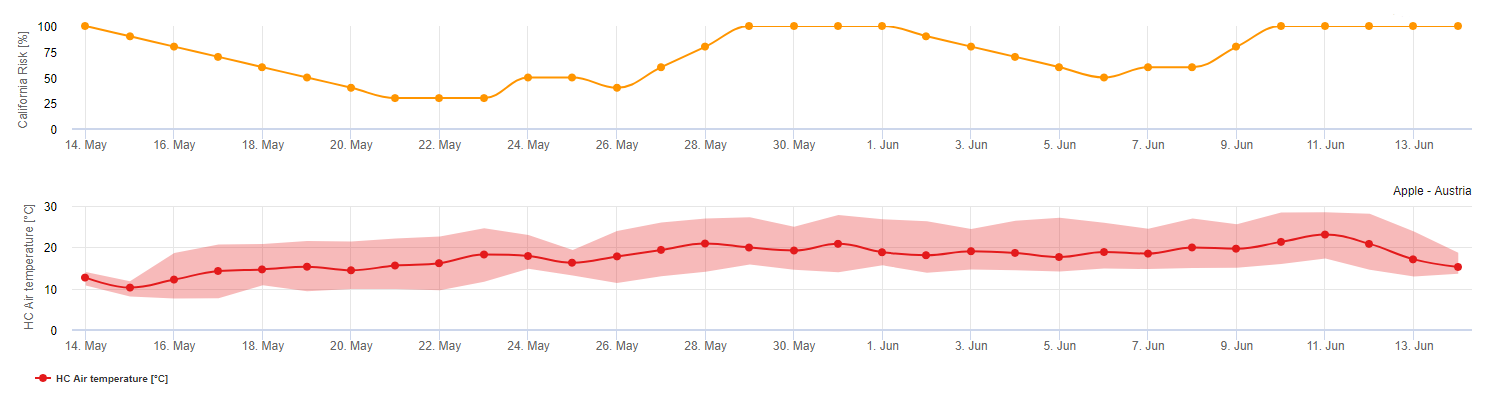
Tras la liberación y germinación de la ascospora (utilizar para ese modelo la INFECCIÓN DE ASCOSPORAS) , el posterior desarrollo y reproducción del oídio se ve influido principalmente por las temperaturas ambiente. El modelo de UC Davis evalúa las temperaturas ambientales y valora el riesgo de desarrollo del oídio mediante un índice de 0 a 100 puntos.
Para que se inicie el índice de evaluación del riesgo, después de la brotación y tras la aparición de la enfermedad, debe haber tres días consecutivos con un mínimo de seis horas consecutivas de temperaturas entre 21 y 30 °C para que se inicie la epidemia de oídio. A lo largo del cálculo del índice, por cada día con 6 o más horas consecutivas entre 21 y 32 °C, el índice aumenta 20 puntos. A continuación, el modelo fluctúa entre 0 y 100 puntos de índice en función de la duración de las temperaturas diarias. El índice gana 20 puntos por cada día que cumpla el requisito de seis horas consecutivas de temperaturas entre 21 y 32 °C, y pierde 10 puntos por cada día que no cumpla el requisito de las seis horas, o si en cualquier momento del día la temperatura accede por encima de 32 °C o está por debajo de 21 °C.
Interpretación del índice:
Los valores bajos del índice de 0-30 indican que el patógeno no se está reproduciendo. Los cultivadores pueden considerar posponer las aplicaciones de fungicidas durante periodos prolongados con valores bajos del índice. Un índice de 40-50 se considera moderado e implicaría una tasa de reproducción del oídio de aproximadamente 15 días. Los valores de índice de 60-100 indican que el patógeno se está reproduciendo rápidamente (cada 5 días) y que el riesgo de una epidemia de la enfermedad es grande.
Modelo de riesgo Pessl Instruments
La dispersión de Uncinula necator durante el verano está influenciada principalmente por temperatura. El antagonista de U. necator llamado Ampelomyces quisqualis se ve favorecido principalmente por la humedad libre. Las temperaturas óptimas para el mildiú polvoroso se sitúan entre 21°C y 32°C. Cuanto mayor sea el número de horas diarias en que la temperatura se encuentre dentro de este intervalo, mayor será el riesgo de que aparezca el mildiú polvoroso (véase el modelo Gubler). Los días con más de 6 horas de temperatura en este intervalo aumentan el riesgo unos 20 puntos. Los días en los que la temperatura no se encuentra dentro de este intervalo óptimo reducen el riesgo unos 10 puntos. Se trata de días con temperaturas inferiores a 21 °C durante todo el día o días en los que la temperatura supera los 32 °C durante 6 horas. Sin embargo, los días con humedad de hoja larga periodos están conduciendo a un establecimiento del hongo antagonista A. quisqualis. Esto conducirá a una disminución de U. necator.
Para este modelo recomendamos seguir estas reglas: Si el riesgo de mildiú polvoroso es inferior a 20 puntos, puede ampliarse el intervalo de pulverización. De 20 a 60 puntos, el intervalo de pulverización normal es válido. Si el riesgo es superior a 60 puntos, debe acortar el intervalo de pulverización.
Comienza el riesgo: con 3 días con igual o más de 6 horas de: 21°C <= Temperatura 60 Puntos
Aumenta el riesgo: Con cada día con igual o más de 6 horas de: 21°C <= Temperatura +20 Puntos
El riesgo disminuye: Con cada día en que la temperatura no alcance los 21°C o cada día con 6 horas de: 32°C -10 Puntos
Tercer modelo de riesgo: Inóculo primario de ascosporas
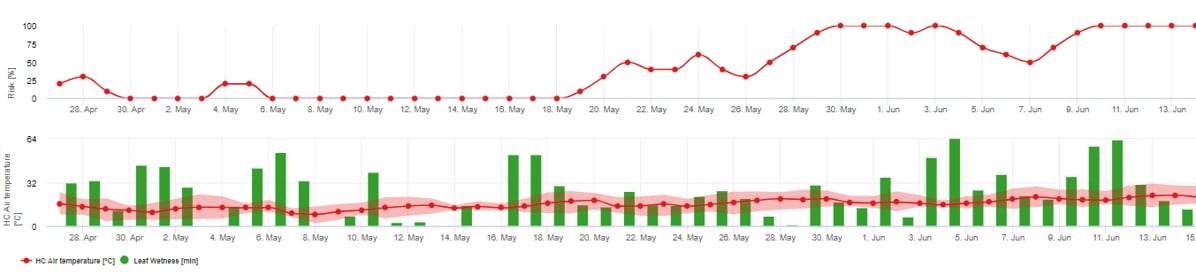
Recomendamos utilizar este modelo cuando las ascosporas se forman durante la época fría del invierno (por lo que no hay infección por micelio ("brotes bandera").
La infección por ascosporas predice la liberación de ascosporas y las infecciones iniciales, que se basan en las temperaturas medias durante los periodos prolongados de humedad de las hojas.
Para el cálculo en fieldclimate.com aproximadamente 2,5 mm de precipitaciones son necesarios para liberar las ascosporas, seguidos de al al menos 8-12 horas de humectación de las hojas tras la liberación de ascosporas a temperaturas entre 10 -15°C para la infección. Una vez que se ha producido la infección, el modelo pasa a una fase de evaluación del riesgo de enfermedad (utiliza el modelo Gubler/Risk) y se basa totalmente en los efectos de la temperatura sobre la tasa de reproducción del patógeno.
Uso práctico
En este modelo mostramos la infección por Uncinula necator en una línea comprendida entre 0 y 100%.
Mostramos dos modelos diferentes en FieldClimate:
1. Las posibles infecciones por ascosporas según los modelos propuestos por David Gadoury y Doug Gubler se muestran en el gráfico para la infección por ascosporas. Si la curva de progreso para el modelo de infección alcanza 100% existe la posibilidad de una infección por ascosporas. De 3 a 5 días después de este evento, los primeros síntomas de oídio pueden ser visibles en las hojas cercanas al cordón viejo.
2. En el gráfico anterior se muestra el riesgo de oídio según el modelo de California y el modelo para zonas de clima moderado. Los periodos con riesgo deben cubrirse con aplicaciones de fungicidas. En periodos con riesgo superior a 60 puntos se recomienda pulverizar con mayor frecuencia especialmente alrededor y poco después de la floración.
La podredumbre negra de la vid afecta a muchos viticultores de Estados Unidos y de la mayoría de las zonas cálidas y áridas del mundo. Para controlar la enfermedad, es importante conocer su ciclo de vida y las condiciones ambientales. Una vez que se produce la infección, existen diferentes métodos para controlar la enfermedad.
Biología
El patógeno de la podredumbre negra de la uva pasa el invierno en muchas partes de la vid y también puede pasar el invierno en el suelo, especialmente en las momias. Además, el patógeno puede pasar el invierno durante "al menos 2 años dentro de las lesiones de los sarmientos infectados que se conservan como sarmientos o espolones".
Una vez, las lluvias podrían liberar las esporas invernantes (ascosporas) que se forman dentro de las momias en el suelo y pueden ser dispersadas por el viento. Algunas de las momias en el suelo pueden tener una descarga significativa de ascosporas que comienza unas 2 o 3 semanas después de la brotación y madurará 1-2 semanas después del inicio de la floración. También puede formarse un segundo tipo de esporas (conidios) dentro de las lesiones de la caña o de las momias que han quedado dentro del "enrejado, y éstas se dispersan a distancias cortas (de centímetros a metros) por salpicaduras de gotas de lluvia". La infección se produce cuando cualquiera de los dos tipos de esporas aterriza en el tejido verde de la uva y el tejido permanece húmedo durante un "periodo de tiempo suficiente, que depende de la temperatura." El periodo durante el cual estas esporas invernantes pueden causar la infección depende de la fuente. Si la fuente de infección es grande, la infección se establecerá pronto. En presencia de humedad, estas ascosporas germinan lentamente, tardando entre 36 y 48 horas, pero acaban penetrando en las hojas jóvenes y los tallos de los frutos (pedicelos). Las infecciones se hacen visibles al cabo de 8 a 25 días. Cuando el clima es húmedo, las ascosporas se producen y liberan durante toda la primavera y el verano, proporcionando una infección primaria continua. (Wilcox, Wayne F. "Podredumbre negra Guignardia bidwellii." Ficha de identificación de la enfermedad nº 102GFSG-D4. 2003. Cornell. 24 oct. 2010).
Así pues, el hongo de la podredumbre negra requiere un clima cálido para crecer de forma óptima; el clima frío ralentiza su crecimiento. También se requiere un periodo de 2 a 3 días de lluvia, llovizna o niebla para la infección." (Ries, Stephen M. "IPM : Reports on Plant Diseases : Black Rot of Grape". Manejo Integrado de Plagas en la Universidad de Illinois. Dic. 1999. 24 oct. 2010). Las gotas de lluvia transfieren estas esporas moviéndolas a diferentes partes de la planta, especialmente a las hojas jóvenes susceptibles. Si hay agua, las conidias germinan en 10 a 15 horas y penetran en el tejido joven. Las nuevas infecciones de podredumbre negra continúan a finales de la primavera y en verano durante periodos prolongados de tiempo cálido y lluvioso. Los conidios son capaces de germinar y causar infección varios meses después de haberse formado. Durante el mes de agosto, los picnidios se transforman en una fase de hibernación que, a su vez, da lugar a pseudotecas dentro de las cuales se producen las esporas de primavera (ascosporas). Las investigaciones han demostrado que las ascosporas son una fuente importante de infecciones primarias en primavera. En primavera, cuando el tiempo es húmedo, los "picnidios de los tejidos infectados absorben agua y los conidios son expulsados. Las conidias son salpicadas aleatoriamente por la lluvia y pueden infectar cualquier tejido joven en menos de 12 horas a temperaturas entre 60-90 grados. (Ellis, Michael A. "Fact sheet Agricultural and Natural Resources: Grape Black Rot". Departamento de Patología Vegetal. Extensión Estatal de la Universidad de Ohio. 2008).
Uso práctico
Para nuestro modelo la base es la literatura publicada por Spotts (Spotts. R. A. 1977. Effect of leaf wetness duration and temperature on the infectivity of Guignardia). A través de discusiones con Daniel Molitor modificamos el modelo original de Spotts hacia un modelo separando en infecciones leves, moderadas y severas similar al diseño de los modelos de la sarna del manzano. (Daniel Molitor: Untersuchungen zur Biologie und Bekämpfung der Schwarzfäule (Guignardia bidwellii) an Weinreben. Disertación Justus-Liebig-Universität Gießen, 2009).
Una infección que cumpla los "criterios Spotts" se considera una infección leve. Las infecciones que cumplen los "criterios de Spotts" por 150% se consideran moderadas y las infecciones que cumplen los "criterios de Spotts" por 200% se consideran graves.
El modelo muestra las fechas de infección para infecciones leves, moderadas y graves. Cuando 100% de una infección se han cumplido las condiciones para la descarga de esporas, la germinación y la infección del tejido vegetal. La gravedad de la infección depende del periodo de temperatura óptima y del periodo de humedad del hongo. Dependiendo de la presión de la infección, las infecciones deben cubrirse preventivamente o debe aplicarse una pulverización curativa poco después de la infección. En el clima semiárido moderado del Mosela o de la mayoría de las zonas vitivinícolas austriacas no sería recomendable una pulverización tras la determinación de una infección leve.
Lobesia botrana (Totricidae, Olethreutinae) se describió por primera vez en Austria, pero procedía del sur de Italia y emigró a toda Europa, el norte y el oeste de África, Oriente Próximo y el este de Rusia. Más recientemente se introdujo en Japón, y en 2008 se informó de su presencia por primera vez en Chile.
Anfitriones
Uva (Vitis vinifera) y el laurel tártago (Daphne gnidium) son los hospedadores preferidos, pero también se ha descrito en la zarzamora (Rubus fruticosus), grosella espinosa (Ribes sp.), grosella negra y roja (Ribes nigurm), oliva (Olea europaea), cereza (Prunus avium), podar (Prunus domestica), el caqui (Diospyrus kakis), kiwi (Actinidia chinensis), granada (Punica granatum), clavel (Dianthus spp.) y otros huéspedes salvajes.
Daños
En mayo y junio, las larvas de primera generación tejen redes y se alimentan de los racimos florales. Las larvas de segunda generación (julio-agosto) se alimentan de bayas verdes. Las larvas jóvenes penetran en las bayas y las ahuecan, dejando la piel y las semillas. Las larvas de tercera generación (agosto-septiembre) son las que causan mayores daños, ya que actúan como telarañas y se alimentan dentro de las bayas y de los racimos, que se contaminan con excrementos. Además, los daños causados a las bayas por la alimentación favorecen la infección por Botrytis y otros hongos secundarios como Aspergillus, Alternaria, Rhizopus, Cladosporium, y Penicillium.
Biología y ciclo vital
La polilla adulta mide aproximadamente 6-8 mm de longitud, con una envergadura de 11-13 mm. La hembra es ligeramente mayor. Tanto los machos como las hembras tienen alas con un patrón similar en mosaico. El primer par de alas (las delanteras) es de color crema tostado, moteado con manchas gris azuladas, marrones y negras. El segundo par de alas es gris con un borde con flecos. En reposo, las alas se mantienen en forma de campana sobre el abdomen. Las hembras ponen los huevos por separado. Los huevos son elípticos y planos, de aproximadamente 0,6-0,8 mm de diámetro. Estos huevos en forma de lenteja son visibles y de color blanco crema iridiscente, tornándose amarillos a medida que se desarrolla el embrión y más tarde negros cuando se forma la cabeza de la larva en desarrollo. Hay cinco estadios larvarios. El primer estadio mide aproximadamente 1 mm y el último entre 12 y 15 mm. Las larvas tienen la cabeza negra, se vuelven marrón amarillento y el cuerpo toma el color de su alimentación (contenido intestinal). El quinto y último estadio larvario hila un capullo de silicona grisáceo para pupar. La pupa mide entre 4 y 9 mm de largo.
En nuestra zona climática, la polilla europea de la vid tiene de dos a tres generaciones al año. En las regiones más cálidas de España, Grecia y Egipto se registran cuatro generaciones. La fase de pupa pasa el invierno dentro de los capullos de silicona (diapausa) bajo la corteza o en grietas del suelo o en lugares ocultos y protegidos. Cuando la temperatura del aire aumenta por encima de 10°C durante unos 10 a 12 días eclosionan los adultos. Primero se encuentran los machos y al cabo de una semana eclosionan las hembras. Así pues, el primer vuelo de los machos puede comenzar ya en la época de la brotación y el vuelo continúa durante unas 4 ó 5 semanas. Las actividades de vuelo tienen lugar cuando sale el sol y las temperaturas son de unos 12ºC. Durante el vuelo, los insectos se aparean y la mayoría de las hembras sólo lo hacen una vez. La puesta de huevos comienza poco después del apareamiento (en uno o dos días). Depositan los huevos individualmente sobre las flores o cerca de ellas. Una hembra pone unos 100 huevos y su vida adulta dura de una a tres semanas, dependiendo de las condiciones climáticas.
En primavera, la eclosión de los huevos tarda 10 días o más, mientras que en verano los huevos eclosionan en 3 ó 4 días, dependiendo de las condiciones de temperatura. Las larvas de primera generación tejen telarañas en las flores y se alimentan de ellas, pueden penetrar en el pedúnculo y provocar la desecación del racimo. El desarrollo de las larvas requiere entre 20 y 30 días, dependiendo de las condiciones climáticas. Después pupan bajo la corteza o en el suelo o sobre la flor y los adultos emergen de nuevo entre 6 y 14 días después.
Las polillas hembras de segundo y tercer vuelo ponen los huevos individualmente directamente sobre bayas sombreadas. Poco después de emerger, la larva entra en una baya y la ahueca mientras se alimenta. Un solo racimo puede estar infestado con varias larvas. Las telarañas, los excrementos y las infecciones fúngicas pueden provocar una gran contaminación del racimo.
El umbral inferior de desarrollo es de unos 10 °C y el superior de 30 °C. Las condiciones óptimas de desarrollo se sitúan entre 26-29°C y una humedad de 40 a 70%. Si la duración del día disminuye y hace más frío, se inicia la fase de diapausa. Las fases larvarias mueren cuando las temperaturas caen por debajo de los 8°C, pero la pupa en diapausa puede resistir incluso los fríos inviernos del norte de Europa.
La primera generación es más corta que las generaciones de verano. Los huevos eclosionan en unos 66 grados-día Celsius (DDC). Según los informes, las larvas que se alimentan de racimos de flores se desarrollan más rápidamente que las que se alimentan de bayas de uva más tarde en la temporada, y esto influye en el tiempo de generación. Las pupas no diapáticas necesitan unos 130 DDC para desarrollarse. Las hembras adultas pueden poner huevos unos 61 DDC después de emerger.
Las estimaciones de DD para una generación varían considerablemente en la bibliografía, de 427 DDC a 577 DDC en la primera generación a 482 DDC a 577 DDC en las generaciones posteriores.
Seguimiento y gestión
Los machos se sienten atraídos por las feromonas, que se utilizan para controlar los vuelos de los machos. Antes de la brotación, coloque trampas con L. botrana señuelos en lo alto de la copa (feromona iScout). Coloque al menos una trampa por cada 30 acres o por cada bloque de viñedo si es más pequeño. Cambie los señuelos según las recomendaciones del fabricante. Revise las trampas semanalmente, registrando el número de polillas capturadas y retirando las polillas atrapadas del fondo pegajoso de la trampa. Grafique las capturas semanales para determinar el inicio y el pico de vuelo de los machos en cada generación. Continuar el seguimiento con trampas hasta el pico del tercer vuelo.
Las aplicaciones de insecticidas deben programarse para la emergencia de las larvas, por lo que el seguimiento de la puesta de huevos y la determinación de su eclosión son esenciales para la gestión de esta plaga. Para la primera generación, la puesta de huevos debe vigilarse desde el pico hasta el final del vuelo. Buscar los huevos en el pedúnculo de 100 racimos, seleccionando un racimo por cepa. Observe el estadio de la mayoría de los huevos encontrados. Los huevos son blancos cuando están recién puestos, se vuelven amarillos y más tarde negros cuando las larvas están próximas a emerger. Un corion translúcido indica que la larva ha emergido. Tras la eclosión de los huevos, busque telarañas en las partes florales. Abra las telarañas y busque daños de alimentación y larvas.
Comenzar el seguimiento de los huevos de segunda y tercera generación en las bayas una semana después de que las primeras polillas del vuelo respectivo queden atrapadas en las trampas. Continuar la búsqueda de huevos semanalmente hasta una semana después del pico de vuelo. Inspeccionar 100 racimos, seleccionando uno por cepa. Seguir controlando los racimos para detectar daños por alimentación (agujeros o bayas huecas), telarañas y presencia de larvas.
En los países en los que L. botrana se establece, las medidas de control se dirigen a la segunda generación. Esto se debe en parte a la prolongada aparición de la primera generación y a la posible reinfestación procedente de viñedos vecinos no tratados. No obstante, se recomienda el tratamiento de la primera generación si las poblaciones son elevadas o si los tratamientos se realizan en toda la zona. Los insecticidas son menos eficaces después del cierre de los racimos. Varios insecticidas de riesgo reducido están registrados para su uso en uvas para el control de larvas de tortrícidos. Entre ellos se incluyen reguladores del crecimiento de insectos, espinosinas y Bacillus thuringiensis.
La interrupción del apareamiento se ha estudiado en Europa durante varios años. Ha demostrado ser más eficaz cuando las poblaciones de polilla del racimo son bajas y cuando se aplica en grandes superficies de más de 10 acres o en toda la zona.
En la bibliografía europea se citan numerosos depredadores y parasitoides. Entre los parasitoides se encuentran 4 especies de moscas taquínidas y cerca de 100 especies de avispas parásitas de las familias de los icneumónidos, bracónidos, pteromalidos y chalicidoideos. Los parásitos de mayor impacto son los que atacan a la pupa invernante. En España son los pteromálidos Dibrachys affinis y D. cavusque causan una mortalidad de pupas de hasta 70%, mientras que en Italia los icneumónidos Dicaelotus inflexus y Captador Campoplex son los más importantes.
Literatura:
- Briere JF, Pracros P. 1998. Comparación de modelos de crecimiento dependientes de la temperatura con el desarrollo de Lobesia botrana (Lepidoptera: Tortricidae). Environ. Entomol. 27: 94-101.
- Coscollá Ramón R. 1981. Algunas consideraciones sobre la dinámica poblacional de Lobesia botrana Den. Schiff. en las comarcas vitícolas valencianas. Bol. Serv. Plagas 7: 169-184.
- Coscollá Ramón R. 1998. Polillas del racimo (Lobesia botrana Den. Y Shiff.). En Los parásitos de la vid, estrategias de protección razonada. Madrid, España. pp. 29-42.
- Armendáriz I, Campillo G, Pérez-Sanz A, Capilla C, Juárez JS, Miranda L. 2007. La polilla del racimo (Lobesia botrana) en la D.O. Arribes, años 2004 a 2006. Bol. San Veg. Plagas 33: 477-489.
- Del Tío R, Martínez JL, Ocete R, Ocete ME. 2001. Estudio de la relación entre las capturas con trampas de feromonas sexuales de Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) y la acumulación de grados-día en viñedos de Jerez (SW de España). J. Appl. Ent. 125: 9-14.
- Gabel B, Mocko V. 1986. Simulación funcional de la polilla europea de la vid Lobesia botrana Den. Et Schiff. (Lep., Torticidae). J. Appl. Ent. 101: 121-127.
- Gallardo A, Ocete R, López MA, Maistrello L, Ortega F, Semedo A, Soria FJ. 2009. Predicción de la actividad de vuelo de Lobesia botrana (Denis & Schiffermüller) (Lepidoptera, Torticidae) en el suroeste de España. J. Appl. Entomol. 133: 626-632.
- Louis F, Schmidt-Tiedemann A, Schirra KJ. 2002. Control of Sparganothis pilleriana Schiff. and Lobesia botrana (Den. & Schiff). in German vineyards using sex pheromone-mediated mating disruption. Bull. IOBC/WPRS 25: 1-9.
- Maher N. 2002. Sélection du site de ponte chez Lobesia botrana (Lepidoptera: Tortricidae): influence de l'infromation chimique non-volatile présente sur les fruits de plantes hôtes. Thése N° 968, Université de Bordeaux 2. pp. 204 (PDF).
- Maher N, Thiéry D. 2006. Daphne gnidium, una posible planta hospedadora nativa de la polilla europea de la vid Lobesia botrana, estimula su oviposición. ¿Es relevante un cambio de hospedador? Chemoecol. 16: 135-144
- Masante-Roca I, Anton S, Delbac L, Dufour MC, Gadenne C. 2007. Atracción de la polilla de la vid hacia partes de la planta hospedantes y no hospedantes en el túnel de viento: efectos de la fenología de la planta, el sexo y el estado de apareamiento. Entomol. Exp. Appl. 122: 239-245.
- Milonas PG, Savopoulou-Soultani M, Stavridis DG. 2001. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Ent. 125: 515-518.
- Moreau J, Benrey B, Thiéry. 2006. Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 96: 205-212.
- Sáenz-de-Cabezón F, Maron V, Zalom F, Pérez-Moreno I. 2005. Efectos de la metoxifenozida sobre huevos, larvas y adultos de Lobesia botrana (Den & Schiff) (Lepidoptera: Torticidae).
- Thiéry D. 2008. Les Tordeuses nuisibles à la vigne. En Les ravageurs de la vigne. Féret, Bordeaux. pp. 15.
- Torres-Vila LM, Stockel J, Roehrich R, Rodríguez-Molina MC. 1997. Relación entre la dispersión y supervivencia de larvas de Lobesia botrana y su densidad en inflorescencias de vid. Entomol. Exp. Appl. 84: 109-114.
- Xuéreb A, Thiéry D. 2006. ¿Varía el parasitismo larvario natural de Lobesia botrana (Lepidoptera: Torticidae) según los años, la generación, la densidad del hospedador y el cultrivar de la vid? Bull. Entomol. Res. 96:105-110.
- Zangheri S, Briolini G, Cravedi P, Duso C, Molinari F, Pasqualini E. 1992. Lobesia botrana (Denis & Schiffermüller). En Lepidotteri dei fruttiferi e della vite. Milán, Italia. Pp. 85-88.
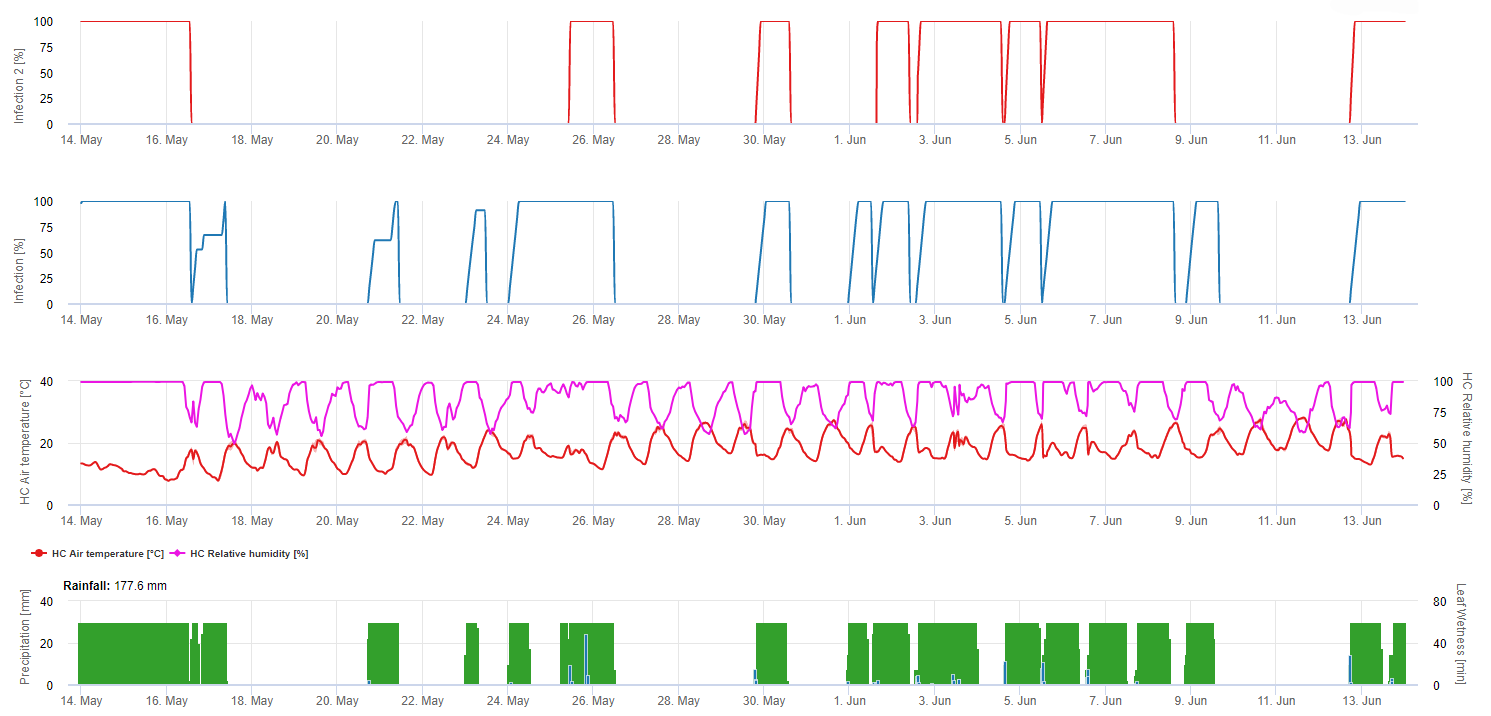
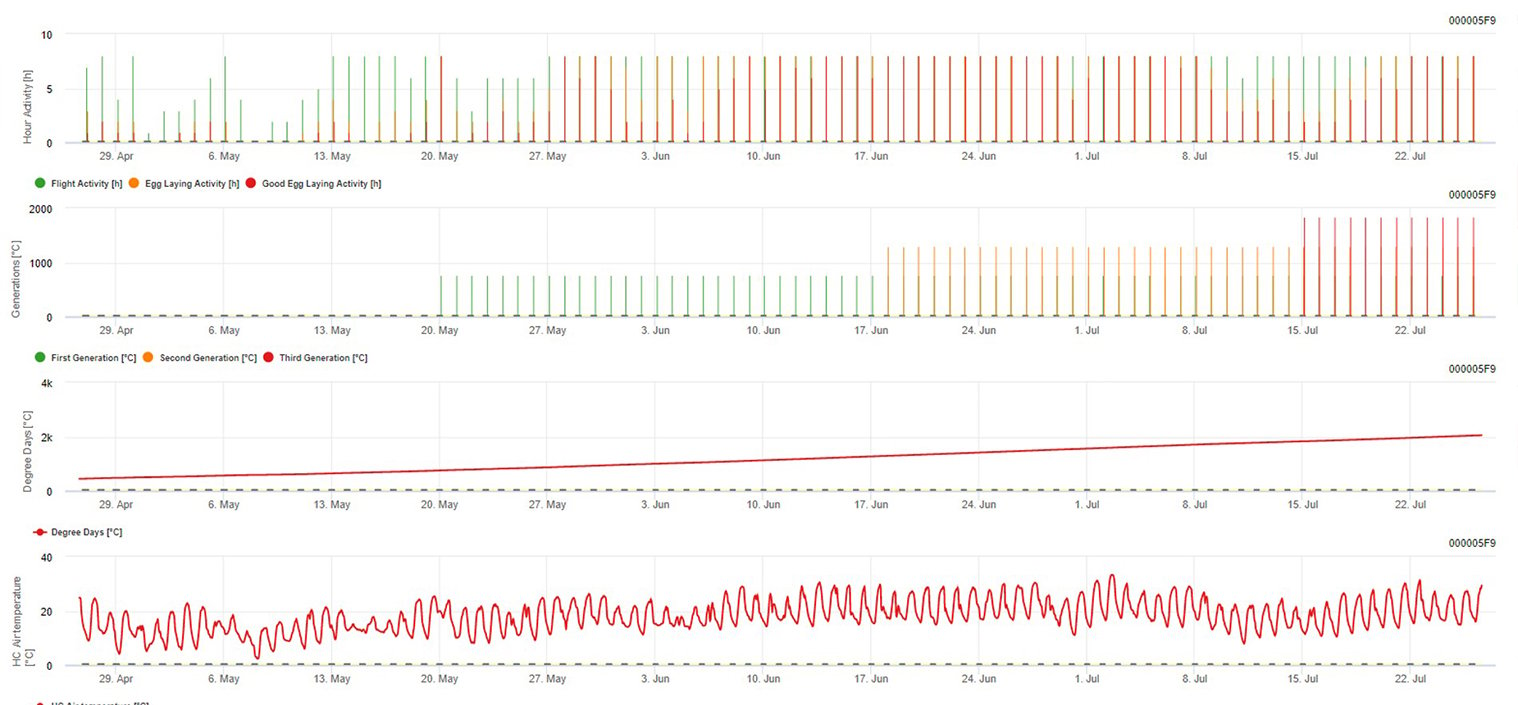
Modelización de la polilla europea de la vid en la plataforma FieldClimate
Sensor necesario: Temperatura del aire
Datos horarios Salida: actividad de vuelo, actividad de puesta de huevos, primera generación, segunda generación y tercera generación, días-grado acumulados
Para la actividad de vuelo, huevo y buena puesta de huevos: comprobamos los datos de las últimas 8 horas (entre las 19:00 p.m. y las 03:00 a.m.) El umbral para la actividad de vuelo es >12°C, para la actividad de puesta > 15°C y para una buena actividad de puesta > 17°C. Si se cumplen todas las condiciones para la actividad de puesta de huevos, se genera la primera generación (= días-grado acumulados). La primera generación +530 DD es la segunda generación (si no se ha previsto debido a las condiciones anteriores). La segunda generación + 530 DD es la tercera generación.
En el gráfico se ve la aparición de la primera generación (buenas condiciones para las tres actividades de vuelo, puesta de huevos y buena puesta de huevos) en unos 469 grados-día acumulados. El vuelo de la segunda generación comienza a mediados de junio con unos 1010 grados-día acumulados y buenas condiciones de temperatura para la puesta de huevos.
Botrytis cinerea es un hongo necrótrofo que afecta a muchas especies vegetales, aunque sus huéspedes más notables pueden ser las uvas. En viticultura, se conoce comúnmente como podredumbre del racimo por botritis; en horticultura, suele denominarse moho gris o moho gris.
El hongo da lugar a dos tipos diferentes de infecciones en las uvas. La primera, la podredumbre gris, es el resultado de condiciones constantemente húmedas o mojadas y suele provocar la pérdida de los racimos afectados. La segunda, la podredumbre noble, se produce cuando las condiciones más secas siguen a las húmedas y puede dar lugar a vinos dulces de postre característicos, como el Sauternes o el Aszú de Tokaj. El nombre de la especie Botrytis cinerea deriva del latín y significa "uvas como cenizas"; aunque poético, lo de "uvas" se refiere al agrupamiento de las esporas del hongo en sus conidióforos, y "cenizas" sólo se refiere al color grisáceo de las masas de esporas. El hongo suele denominarse anamorfo (forma asexual) porque rara vez se observa la fase sexual. El teleomorfo (forma sexual) es un ascomiceto, Botryotinia cinerea.
Biología de B. cinerea
En frutas y uvas, las infecciones florales seguidas de latencia están teniendo un gran impacto en la epidemiología del moho gris. Se pueden distinguir varias vías de infección de la flor a la fruta. En uvas, kiwis y frambuesas se han postulado infecciones a través del estilete hasta el óvulo. En el óvulo, el patógeno permanece latente, lo que parece ser el resultado de una estrategia de defensa del huésped preformada (similar al contenido de resveratrol de la baya de uva joven). En las uvas, los kiwis y las fresas, las infecciones a través de los estambres, los pétalos o los sépalos son importantes. En las uvas, los estudios han demostrado que B. cinerea puede infectar los estambres y crece basalmente para infectar el receptáculo y, a continuación, crece sistémicamente hasta el pedicelo y los tejidos vasculares de las bayas.
Un proyecto de investigación de 6 años en la zona del Cabo demostró que las bayas de uva pueden infectarse a través del estoma y las lenticelas del pedicelo y en menor medida del raquis. Las infecciones del pedicelo también son posibles durante el periodo de floración. Posteriormente, este tejido aumenta la resistencia contra B. cinerea infecciones.
Otras vías de infección postulan el crecimiento saprofítico del patógeno en los restos florales y la posterior infección de las bayas cuando la susceptibilidad aumenta con la maduración o por daños de las bayas causados por insectos o granizo. Otra tesis es la acumulación de conidios en la fruta durante el verano y la infección de bayas susceptibles más adelante en la temporada. La infección conidial de la fruta madura es posible a partir de cualquier fuente de inóculo. Lo más probable es que se forme un número bajo de bayas infectadas de forma latente, que muestran una esporulación extensa cuando la susceptibilidad de las bayas aumenta con la madurez. Se puede suponer que las bayas se vuelven susceptibles a partir de un contenido de azúcar de 6%.
En el kiwi, las condiciones de recolección influyen mucho en la aparición de B. cinerea. Las frutas recogidas con la superficie húmeda pueden infectarse por B. cinerea en las microlesiones provocadas por los dedos de los recolectores.
En el control práctico de B. cinerea tenemos que separar dos periodos importantes de infección: La floración y la senescencia. 1) Durante la floración tenemos frutos jóvenes susceptibles, en los que la infección va seguida de un periodo de latencia. 2) Mientras que la infección en frutos maduros (senescentes) dará lugar a síntomas sin periodo de latencia. La importancia de la infección durante la floración en las uvas puede cambiar de una temporada a otra y de una región a otra. En frutas en las que hay que esperar cierta vida útil (uvas de mesa, kiwis o fresas) los síntomas se observan cuando se almacenan en condiciones refrigeradas en tiendas o almacenes. Control químico de las uvas de vid que muestran buena resistencia a B. cinerea durante la floración no mostrará ningún resultado económico. Por lo tanto, todas las condiciones de riesgo y probabilidad de infección, la susceptibilidad de la fruta y la vida útil, las condiciones de almacenamiento deben tenerse en cuenta en la decisión de una aplicación contra Botrytis cinerea durante la floración.
En las frutas de hueso, la infección por B. cinerea se produce sobre todo durante la floración. En esta época, los tratamientos contra Monilina spp. que también infectan la Botrytis cinerea infección.
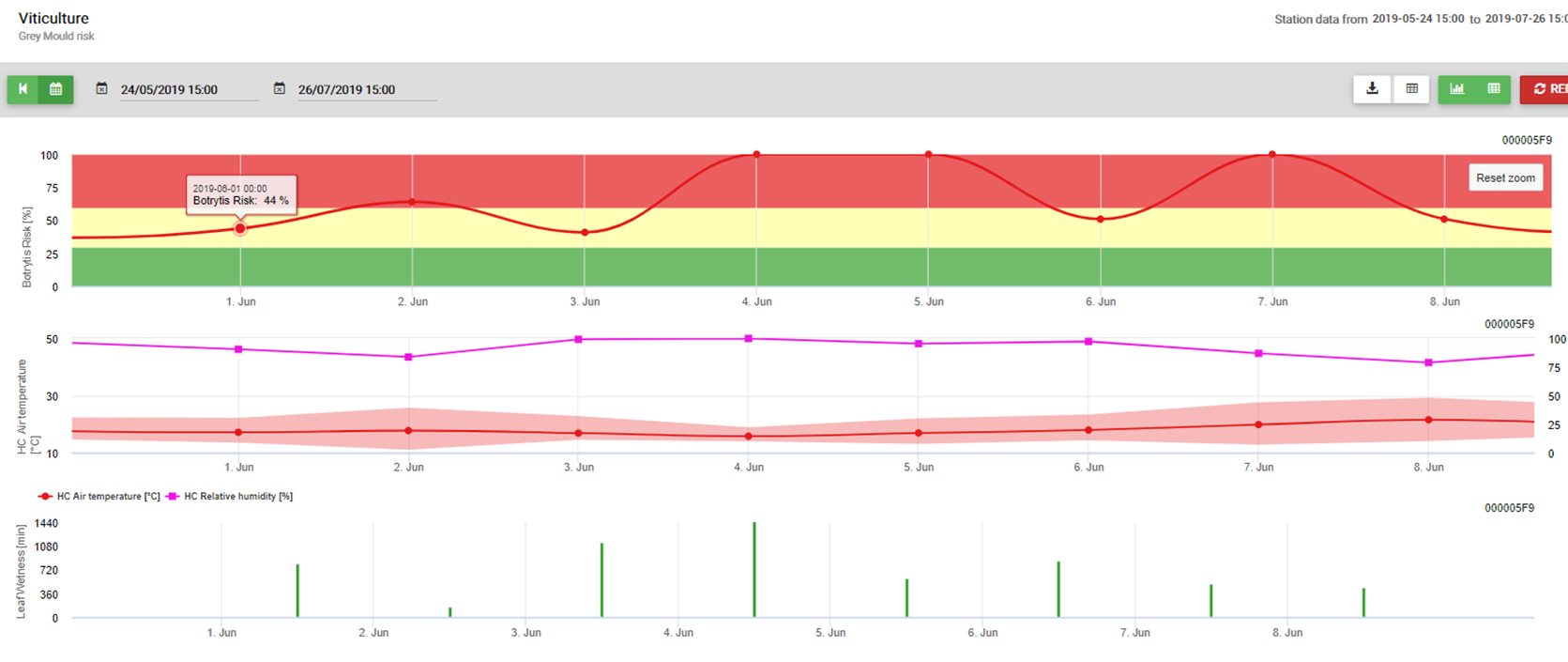
Modelo de B. cinerea y uso práctico
Sensores necesarios: Humedad de la hoja, temperatura, humedad relativa
Botrytis cinerea es un parásito facultativo. Crece también en material vegetal muerto. Por este motivo, siempre está presente en viñedos y huertos. Botrytis cinerea está relacionada con un clima húmedo. Para la infección, necesita una humedad relativa muy alta o la presencia de agua libre (sensor: humedad de la hoja, humedad relativa). El hongo es incapaz de infectar material vegetal adulto sano mediante esporas. La infección se produce en los sarmientos jóvenes de la vid durante los periodos húmedos más largos o en los sarmientos dañados por las tormentas de granizo.
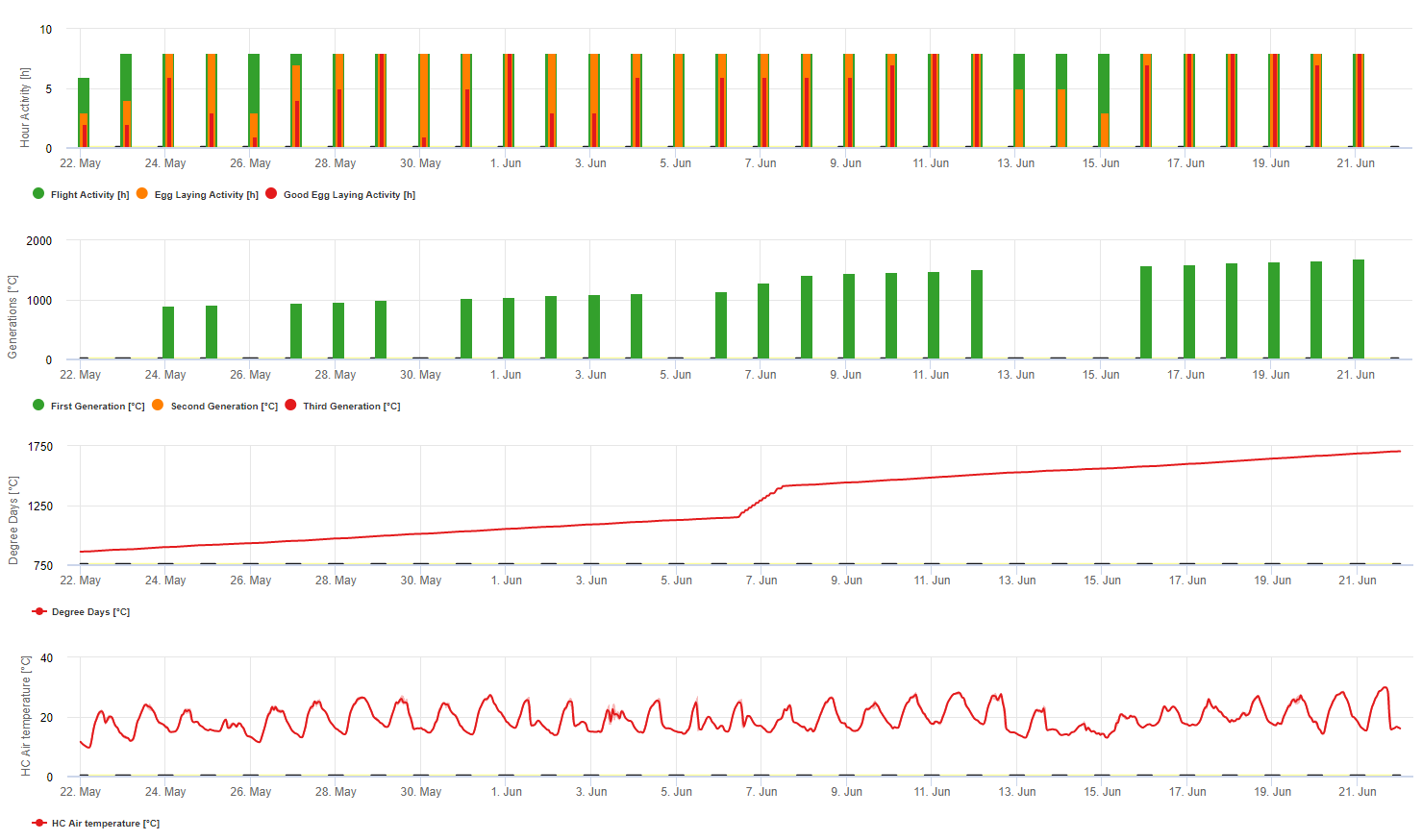
El modelo utiliza la siguiente correlación entre la duración de la humedad de las hojas y la temperatura para calcular el riesgo de infección.
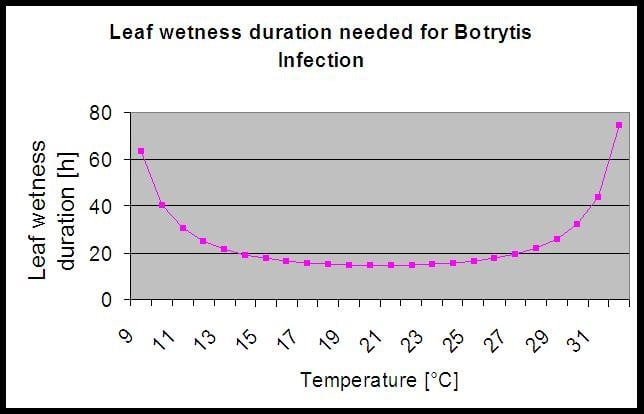
En Botrytis El modelo de riesgo da como resultado un valor de riesgo de 0 a 100%. Este valor indica la presión de B. cinerea en ese momento. Si tenemos un valor de 100% significa que ha habido varias veces un período de humedad lo suficientemente largo como para infectar el tejido susceptible (calculamos los llamados "puntos húmedos" (matriz entre la humedad de la hoja, la temperatura con un máximo de inicialmente 38400 puntos (comienzo de la temporada, que muestra 30% riesgo). Después de este periodo, cada periodo húmedo con unos 4000 puntos húmedos (matriz) aumenta el riesgo con 10% o, por otro lado, cada periodo seco reduce el riesgo en 1/5 del valor anterior. Una aplicación contra
Una solicitud contra B. cinerea depende de la fruta y del objetivo de producción.
La antracnosis, mancha negra, podredumbre de ojo de pájaro, mancha de ojo de pájaro se encuentra en todo el mundo. La enfermedad reduce la calidad y el rendimiento de la fruta, debilita la vid mediante la destrucción significativa de nuevos brotes y hojas. La mayoría de las especies de Vitis son sensibles a la antracnosis.
En las temporadas húmedas de cultivo de la vid, la enfermedad está bien establecida y causa daños en cultivares muy susceptibles tras las lluvias de principios de temporada.
Una vez establecida en los viñedos, la enfermedad puede ser muy difícil de controlar. Sin embargo, en Europa la enfermedad es casi insignificante debido a los fungicidas con compuestos de cobre, ditiocarbamatos, ftalimidas para controlar el mildiú velloso y a la falta de precipitaciones en los climas áridos.
Síntomas
La enfermedad ataca las partes aéreas y suculentas de la vid, incluidos los sarmientos jóvenes, las hojas, los pecíolos, los zarcillos y los racimos; las lesiones más comunes se producen en los sarmientos y las bayas. Los primeros síntomas aparecen como manchas aisladas, pequeñas y circulares, que se vuelven parduscas, hundidas, con centros grises y márgenes oscuros, redondos o angulosos. A veces los síntomas parecen lesiones por granizo. En las hojas aparecen numerosas manchas pequeñas, marrones, que son grises en el centro, el centro necrótico suele caer, creando una apariencia de "agujero de tiro". Las hojas jóvenes son más susceptibles a las infecciones. En las bayas aparecen pequeñas manchas circulares de color marrón rojizo, que se hunden ligeramente. El centro se vuelve gris y está rodeado de márgenes de color marrón rojizo a negro (ojo de pájaro).
Ciclo de vida del patógeno
La enfermedad está causada por el patógeno Elsinöe ampelina (de Bary) Shear.
Las cañas infectadas son la principal fuente de la enfermedad. Los esclerocios o micelios que sobreviven en las lesiones se activan en primavera y producen conidios en condiciones húmedas (lluvia o rocío durante 24 horas) dentro de un régimen de temperaturas de 2°C-40°C. Los conidios son salpicados por la lluvia a nuevos tejidos. Los conidios son salpicados por la lluvia a nuevos tejidos. Germinan y producen nuevas infecciones. El clima cálido reduce el tiempo de mojado necesario para la infección inicial y el periodo de incubación antes de que los síntomas sean visibles en las hojas. La germinación óptima de las esporas se produce a 25-30°C, con un mínimo de 3 a 4 horas de humectación necesaria de las hojas. Además, el tiempo de incubación es más corto dentro de este rango de temperatura (3-4 días en condiciones ideales de humedad). En infecciones a temperaturas en torno a los 10°C, los síntomas de la enfermedad necesitan unos 14 días para ser visibles en las hojas.
Las conidias o ascosporas formadas en bayas infectadas que pasan el invierno en la vid o en el suelo también pueden causar infecciones primarias. Las esporas infectan las hojas nuevas, los brotes, los zarcillos y las bayas jóvenes y producen lesiones en condiciones húmedas. Estas conidias sirven como inóculo secundario y son responsables de nuevas infecciones en la temporada. Las conidias son desalojadas por las gotas de lluvia y dispersadas por el viñedo.
El desarrollo epidémico se debe a la susceptibilidad del tejido de la vid, la dispersión de los conidios y las condiciones meteorológicas imperantes. El factor más importante es la frecuencia de los periodos de lluvia.
Modelización en Fieldclimate
Desarrollo de las esporas de hibernación: Temperatura: 2-40C°, humedad relativa superior a 90% o humedad de la hoja: cuando las esporas se desarrollan (100%) y se mantienen las condiciones de humedad de la hoja y temperatura, se empieza a calcular la infección. El desarrollo de las esporas (humedad relativa inferior a 50%) y la infección dejan de calcularse cuando la humedad es demasiado baja. La gravedad de la infección depende de las condiciones de humedad (evento de lluvia).