Soybean Rust, in specific the Asian soybean rust is a serious disease of soybean caused by the fungal pathogen Phakopsora pachyrhizi. Distribution of the disease: Asia and Australia, and more recently from Africa, North America and South America. The disease was first observed in Japan in 1902. The pathogen was distributed throughout Asia and Australia through America. Soybean rust cannot overwinter in areas with freezing temperatures, but it can spread by wind rapidly over such large distances, its development can be so explosive, and it can cause such rapid loss of leaves that it is now one of the most feared diseases in the world’s soybean-growing areas.
Symptoms and Signs
The first symptoms of soybean rust caused by Phakopsora pachyrhizi begin as very small brown or brick-red spots on leaves. In the field, these spots usually begin in the lower canopy at or after flowering, although seedlings can be infected under certain circumstances. Often the first lesions appear toward the base of the leaflet near the petiole and leaf veins. This part of the leaflet probably retains dew longer, making conditions more favorable for infection. Lesions remain small (2-5 mm in diameter), but increase in number as the disease progresses. Pustules, called uredinia, form in these lesions, mostly on the lower leaf surface, and they can produce many urediniospores.
The raised pustules can be seen with the unaided eye, especially when sporulating. Even though the lesions are small, each lesion often has several pustules (uredinia) . Lesions can be completely covered in urediniospores when the pustules are active. Soybean rust urediniospores are pale yellow-brown to colorless, with an echinulate (short spines) surface ornamentation. This coloration is different from many other rust pathogens whose spores are often reddish-brown (rust colored). Germination of P. pachyrhizi urediniospores occurs through an equatorial (central) pore, producing a germ tube that ends in an appressorium, which the fungus uses to penetrate the host directly or through a stoma.
As more and more lesions form on a leaflet, the affected area begins to yellow, and eventually the leaflet falls from the plant. While soybean rust usually begins in the lower canopy, it quickly progresses up the plant until all of the leaves have some level of disease. Severely diseased plants may become completely defoliated. The loss of effective leaf tissue results in yield reductions from both fewer and smaller seed. Yield losses as high as 30 to 80% have been reported, but the amount of loss depends on when the disease begins and how rapidly it progresses. Besides leaves, soybean rust can also appear on petioles, stems, and even cotyledons, but most rust lesions occur on leaves.
Lesions may be either tan or red-brown. Tan lesions have many pustules that produce numerous urediniospores. Red-brown lesions, thought to be a moderate resistance reaction, have only a few pustules that produce only a few urediniospores. As will be discussed in the Disease Management section, this lesion type depends on the strain of the pathogen, and may appear on the same leaf with tan lesions, or tan lesions may turn red-brown with age. Symptoms and signs on other hosts, such as kudzu, are similar, although lesion size may differ. As pustules age, they may turn black. This is caused by the formation of a layer of teliospores in the pustules, turning pustules from uredinia into telia.
Teliospores have two functions: survival of the fungus in the absence of a living host (overseasoning) and sexual reproduction. The thick walls of the teliospores protect the fungus from the environment and attack by other organisms. In rusts, the teliospores germinate forming a basidium and four basidiospores during which sexual recombination occurs. Germination of P. pachyrhizi teliospores has been observed only in the laboratory and does not seem to make a significant contribution to the perpetuation of this disease in the field.
Pathogen Biology
There are two closely related fungi that cause rust on soybean: Phakopsora pachyrhizi, sometimes referred to as the Asian or Australasian soybean rust pathogen, but which now also occurs in the western hemisphere, and P. meibomiae, the so-called New World soybean rust pathogen, which is found only in the western hemisphere. Except for a few minor characteristics, the two fungi appear morphologically identical, but P. pachyrhizi is much more aggressive on soybean than P. meibomiae. To date, P. meibomiae has not been documented to cause significant yield losses in Central and South America. The two species can be distinguished by using DNA analysis protocols.
Like other rusts, the soybean rust pathogens are obligate parasites that require a living host to grow and reproduce. They can survive away from the host as urediniospores for only a few days under natural conditions. Both soybean rust pathogens, to the best of our knowledge, produce only two types of spores: urediniospores and teliospores (Figure 15). This contrasts with other rusts, which can have up to five spores stages (for example, wheat stem rust). For soybean rust, like most rusts, the uredinial stage is the repeating stage. This means that urediniospores can infect the same host on which they were produced (soybean) during the same season. Epidemics can develop quickly from only a few pustules because spore-producing pustules are produced in as little as 7 to 10 days after infection, and each pustule can produce hundreds of urediniospores. Teliospores are produced in old lesions, but they do not appear to germinate in nature, and no alternate host, nor aecia or spermogonia are known.
Without germination of teliospores, sexual reproduction cannot take place. Lack of sexual reproduction should limit variability of the rust fungus, but nevertheless there is substantial variability in P. pachyrhizi with respect to virulence. This has limited the use of single genes for resistance in soybean, because in a short time new isolates of the pathogen arise that overcome the resistance gene. It is not known how this variability originates in P. pachyrhizi. Wheat stripe rust, Puccinia striiformis, has a similar life cycle as P. pachyrhizi with no functioning telial stage and therefore no sexual reproduction, but has many races. It may be that each resistance gene is so specific that a single mutation in the right gene of the fungus allows it to be virulent on hosts with the new resistance gene.
Epidemiology
Soybean rust epidemics begin with the arrival of airborne inoculum (urediniospores). This pathogen is unique among rusts because it has many alternative hosts, which may serve as sources of inoculum. Alternative hosts are other plants that can become infected with the same pathogen, but are not required to complete the pathogen’s life cycle. Alternative hosts are not to be confused with alternate host, which is a plant other than the principal host, that is needed for a pathogen to complete its life cycle. In frost-free areas, such as South America, Central America, the Caribbean basin, southern Texas, and Florida, the inoculum source could be nearby on volunteer soybean plants, kudzu, or some other alternative host. In areas that experience frost, such as the Midwestern United States, inoculum must be blown in from over-wintering sources that may be hundreds of miles away.
Re-introduction of obligate pathogens into a distant region occurs with several other diseases, such as wheat stem rust and downy mildews, e.g. blue mold on tobacco. Because spores of P. pachyrhizi are sensitive to ultraviolet radiation, long distance movement of these rust spores probably occurs in storm systems where clouds protect the spores from the sun. Once viable spores have landed on the leaf surface of a suitable host, infection and subsequent epidemic development are dependent on environmental conditions. Generally, infection occurs when leaves are wet and temperature is between 8°C and 28°C, with an optimum of 16°C to 28°C. At 25°C, some infection occurs in as little as 6 hours of leaf wetness, but 12 hours are optimal. After infection, lesions and pustules with urediniospores can appear within 7 or 8 days, and the next infection cycle is set to begin.
This short life cycle means that, under the right conditions, soybean rust epidemics can quickly build up from almost undetectable levels to very high levels. Soybean rust epidemics can progress from below detectable levels to defoliation within a month. Epidemics may seem to progress even faster than that, because early infections occur in the lower canopy and are hard to find. Besides the environment, plant age affects soybean rust epidemics. Usually, rust lesions are not found on soybean until flowering, unless there are high inoculum levels early in the season. This may be due to greater susceptibility of plants to rust as the host enters the reproductive stages, it may be because in lower parts of the canopy spores are more protected from UV radiation, or it may be because conditions in the canopy become more humid as the canopy closes. In any event, lesions can form at any growth stage, but major increases in disease do not occur until after flowering.
Two models for soybean rust are described and calculated in FieldClimate:
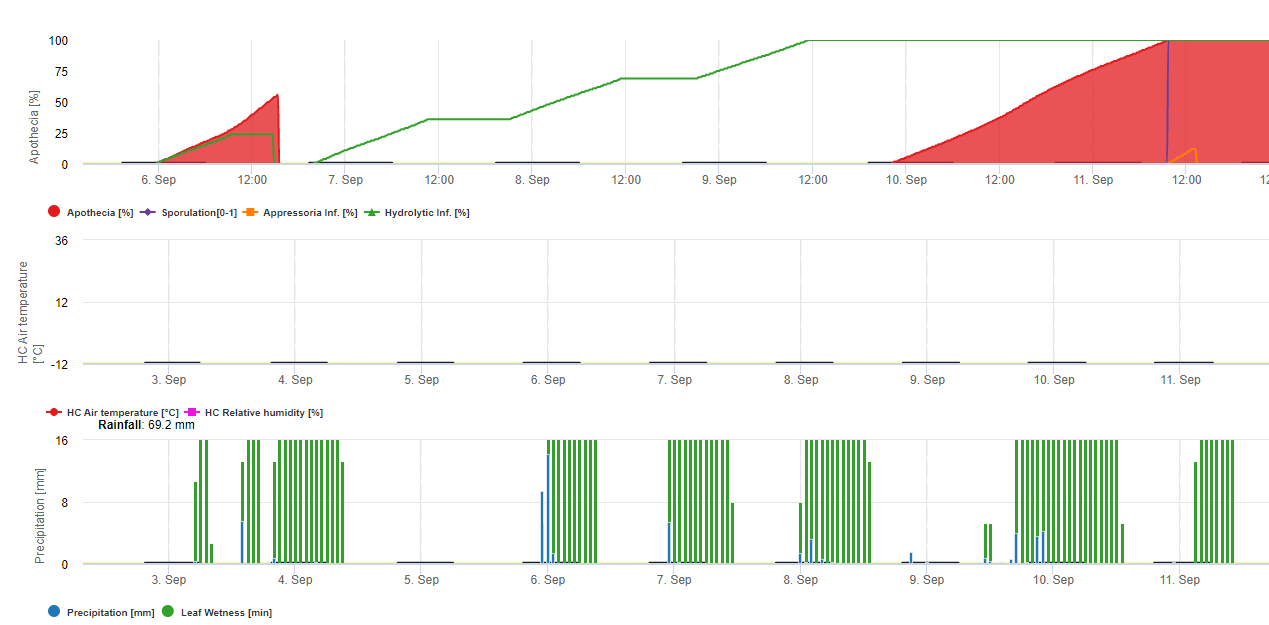
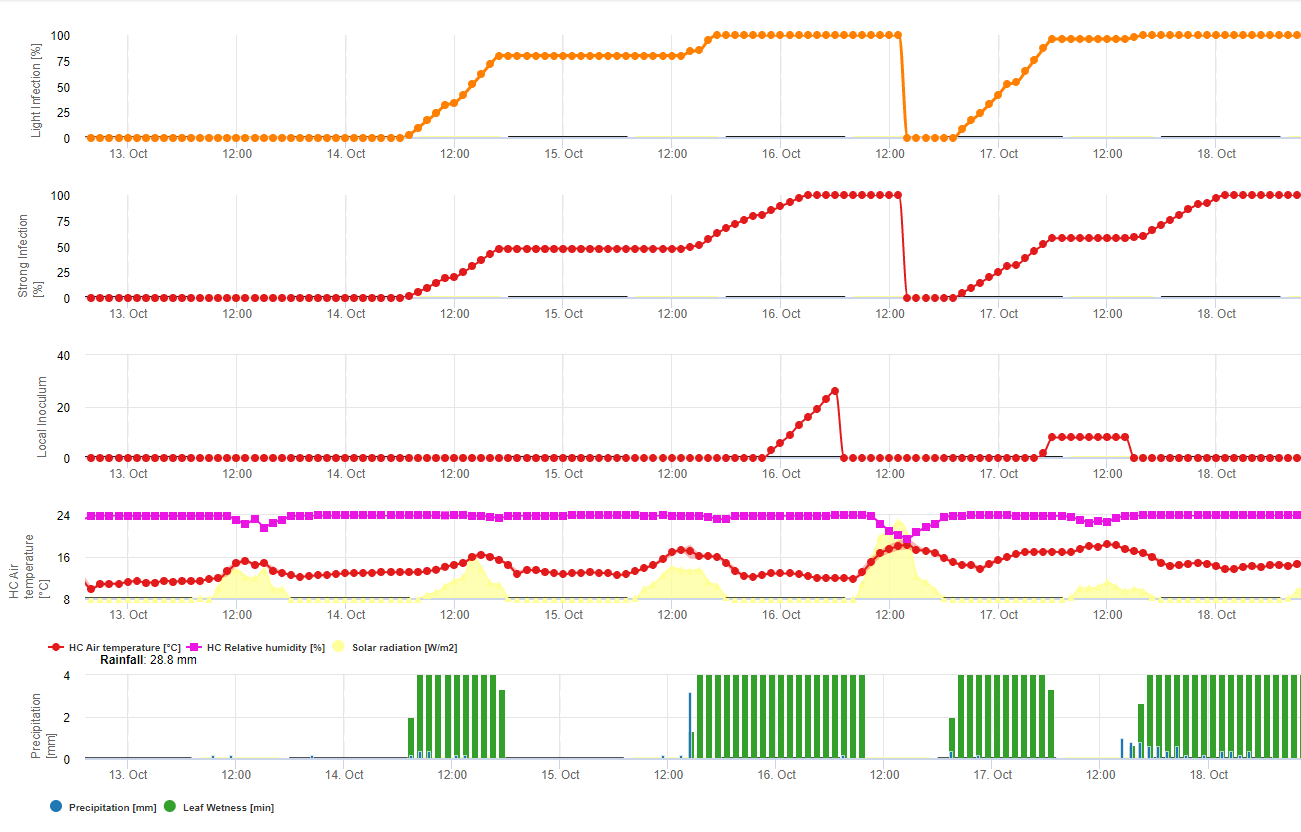
Model 1: Main factors for calculation of the infection and severity of the soybean rust are the leaf wetness and the temperature (be careful infection has to be 100% till severity rates (from 1-3) start to be calculated) Leafs have to be wet and temperature has to be between 8 and 28°C – infection starts to calculate (sum of 7200 °minutes is 100%). If infection is 100% and in dependance of temperature and duration of leaf wetness the severity classes from 1- 3 start to be calcuated.
Model 2: Here much more factors are included into the calculation of Soybean rust: Temperature, Leaf wetness, relative Humidtiy, Precipitation and solar radiation (fungal hyphae are able to enter the stomata during day time).
When infection of soybean rust reached 100% the severity starts to be calculated. If both (infection and severity are 100%) the occurrence (establishment?) of local inoculum is predicted.
Literature:
- Ivancovich, A. y Botta, G. 2002. La roya de la sojo en la Argentinia. EEA Pergamino. RevistadeTEchnologia, Agropecuaria Vol. 7 (21), pp 16-17.
- Melching, J. S., Dowler, W. M., Koogle, D. L., and Royer, M. H. 1989. Effects of duration, frequency, and temperature of leaf wetness periods on soybean rust. Plant Dis. 73:117-122.
- X.B. Yang, E.M. Del Ponte, and A.P. Dias. Knowing the risk of soybean rust by comparison with Brazil. Department of Plant Pathology, Iowa State University
- Yang, X. B. 1995. Assessment and management of the risk of soybean rust. Proceeding of the soybean rust workshop, 9-11 August 1995. J. B.Sinclair and G. L. Hartman, eds. National Soybean Research Laboratory, Urbana, IL
Sclerotinia stem rot or white mold of soybeans is a disease caused by the fungus Sclerotinia sclerotiorum. The disease can cause major seed yield reductions or even completely destroy a crop when soybeans are planted in infested soil and there is a dense plant canopy with prolonged periods of wet weather. Yield losses usually occur when incidence of disease is 15 % or greater. In most years throughout North Dakota, Sclerotinia stem rot is only a minor to moderate problem for growers, and the disease is rarely observed during drought years. Wet weather is a major factor in disease development. It has been a persistent problem in irrigated soybeans. Besides seed yield reductions, the disease also results in reduced seed quality and seed contaminated with the black sclerotia of the fungus. Seed contamination can be a serious problem for exported seed since it may result in rejection of the seed lot at foreign ports of entry. Furthermore, sclerotia returned to the soil can affect other crops in the rotation. Sclerotinia stem rot can be managed by growers through an understanding of the pathogen and disease cycle.
Symptoms
The symptoms are usually not observed until the crop canopy between rows has closed, creating a humid microclimate. Wilting and withering of leaves followed by death of plants are usually the first symptoms observed. A close inspection under the plant canopy will reveal a cottony, white mycelial (fungus threads) growth on stems, leaves or pods . Lesions develop on main stems and side branches. Eventually, lesions girdle the stems and the plant parts above die. Stems appear bleached and sometimes shredded from advanced decay. Large, black sclerotia of varying shapes and sizes will form from the white mycelium growing on plant tissue. Sclerotia also form in the stem pith and have a characteristic cylindrical shape. Seeds in diseased pods are usually shriveled and may be infected by the fungus, or replaced by black sclerotia. The seed is usually contaminated with sclerotia when infected plants are harvested.
Pathogen Biology
The fungus has an extensive host range of over 370 plant species and causes diseases on a wide variety of crops such as sunflower, dry bean, canola (rapeseed), potato, alfalfa, buckwheat, lupine, mustard, Jerusalem artichoke, safflower, lentil, flax, field peas and many vegetables. In North Dakota it is rare to find this pathogen causing serious damage on some of these crops such as flax and potato. There are also many common broadleaf weed hosts such as marsh elder, lambsquarter, pigweed, Canada thistle and wild mustard. The fungus that causes white mold on soybean is the same one that causes white mold or Sclerotinia disease of sunflower, dry beans, canola, and other crops. Sclerotinia sclerotiorum overwinters principally as sclerotia in soil. The sclerotia germinate to form small tan to brown mushrooms called apothecia (about one-eighth to one-fourth inch in diameter) . These produce spores termed ascospores which initiate the disease on soybean and other susceptible crops.
Disease Cycle
Moisture and flowering are critical factors in disease development. Disease usually does not occur before the closing of the crop canopy, because a dense canopy promotes cool temperatures and a humid microclimate around the stems and maintains high soil moisture following rain or irrigation. Initiation of disease is also closely associated with flowering. Following seven to 14 days of high soil moisture, sclerotia within the upper several inches of soil will germinate to form the mushroom-like apothecia. A single sclerotium can produce several apothecia. The apothecia will forcibly eject their ascospores into the air where they will be carried by air currents to soybean plants. The most important source of ascospores is from apothecia produced within the field, but ascospores can blow in from adjacent or nearby fields. One apothecium can produce enormous quantities of ascospores over several days. Ascospores survive for short periods on plant tissue but do not overwinter. Ascospores require a film of water and a food base such as dead or senescing flower tissues to germinate and grow before they infect the plant. Flower tissue is the most important food base for initiating infections. Often infections will start in stem axils where senescing flower tissues have fallen and lodged. Infections may also occur through wounds caused by hail or other injury. A film of water on the plant surface promotes development of lesions and increases the amount of tissue damage. Initial disease development generally requires greater than 40 hours of continuous plant surface wetness, but once disease is started, shorter periods of wetness will allow lesion development. This is the reason disease is associated with lengthy periods of cloudy, humid, rainy weather. As plant surfaces dry, the progress of disease is slowed. Cool temperatures between 59 and 75 degrees Fahrenheit favor disease development. The greater the density of the plant canopy the more favorable the environmental conditions are for disease. Solid seeded soybeans and lodged crops are therefore more prone to disease development. Sclerotia will be formed as the mycelium grows in and on the plant tissues. These sclerotia will not germinate to form more apothecia during the season, but rather will be returned to the soil during harvest and tillage operations and will over-winter to become inoculum (source of infective fungus) for a future susceptible crop. Sclerotia are highly resistant structures and survive for long periods in soil.
Source: http://www.ndsu.edu/pubweb/~bernelso/soydiseases/sclerotinia.shtml
In FieldClimate the model for Sclerotinia is calculated in dependence of a rainy periods, the relative humidity and temperature as well as leaf wetness. During long wet periodes an infection through the establishing of an appressorium by the fungal pathogen is recommended. Another way of infection is called “hydrolytic infection”. This way is based on the release of hydrolytic enzymes that sequentially degrade the plant cuticle, middle lamellae, and primary and secondary cell walls and entire the plant. Model described in detail on this page.