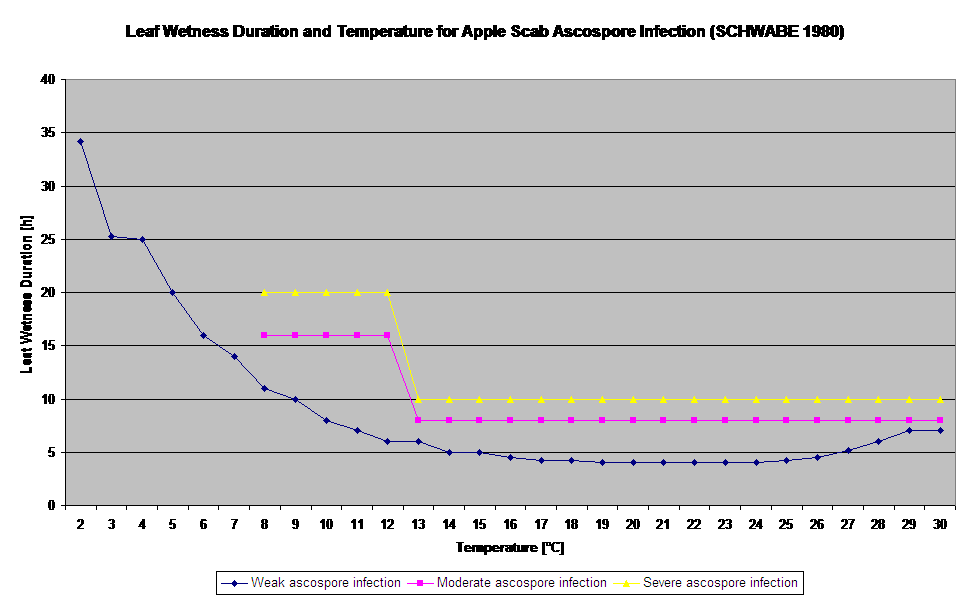
Germination and Penetration of ascospores of Venturia inaequalis on leaves or fruits depend on leaf wetness periods (wet leaves or rel. humidity >80% (holds the infection level till under 80% r.h.). and air temperature (above 2°C). Higher temperatures lead under wet conditions to faster penetration, while at lower temperatures longer leaf wetness periods are needed. This relationship was first published by MILLS and LAPLANTE (1945). Our calculation is carried out based on the publications of SCHWABE (1980). SCHWABE described the severity of scab infections depending on the temperature. These observations are in congruence with our own observations.
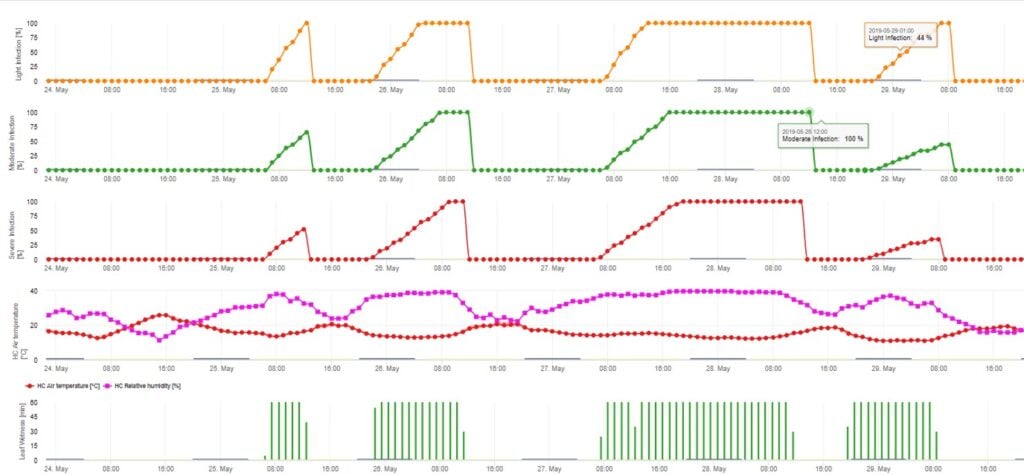
The Ascospore Infection Model calculates the progress of infection, depending on leaf wetness and temperature, for weak, moderate and severe infections. To split the prediction into three “severity classes” is very traditional for apple scab models. The progress of infection is displayed as a graphic for hourly values. An infection has already taken place as soon as the progress curve for a weak infection reaches 100%. A moderate or severe infection is complete if the progress curves for this severity classes are reaching (100%). Often in practice or in reality an infection reaches 70% or 80% early in the morning and in dependence of the actual weather you will be able to know if this infection will finish or not. Displaying the daily maximum values of the infection process curve gives an overview over the expected infection dates in the season.
Primary use of the model
For the market it is absolutely important to have disease free apple fruits to get a good price. Therefore, growers tend to have no scab tolerance at all in their orchards. The model should be implemented into a curative application program (application of sprays containing compounds of cyprodinil or pyrimethanil at pre-bloom) when the model pointed out an infection (no matter if weak or severe). In the bloom and early post bloom period, severe infections are most frequent and all humid periods have been covered by protective sprays. Further on plant tissue grows very fast, therefore a protection may only work for 4 days. In this case a curative spray after an incomplete infection period might be also needed. But in this case, it is relevant if it has been a weak or severe infection and also if there was a low or high number of ascospores ready for infection.
Literature
- Schwabe WFS, 1980. Wetting and temperature requirements for apple leaf infection by Venturia inaequalis in South Africa. Phytophylactica, 69-0.
- Schwabe WFS, 1980. Weather favouring apple scab infection in South Africa. Phytophylactica 12, 213-217.
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Secondary infections by conidia
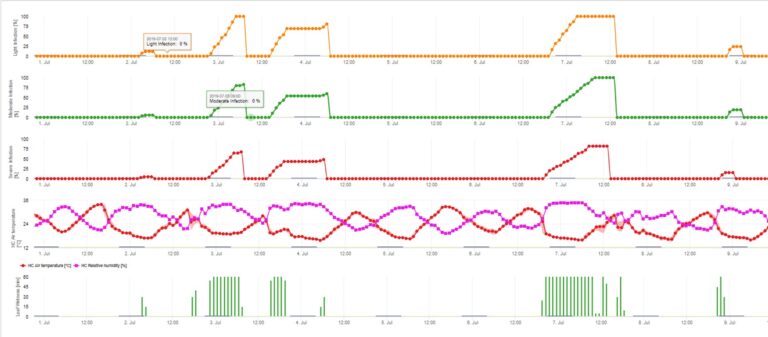
Infections of apple scab by conidia (asexual) depend on the same factors as infections caused by ascospores. The difference is, that conidia are able to be discharged under light and dark conditions and conidial infections take mostly place during summer time.
Depending on air temperature regimes the conidia of Venturia inaequalis need longer or shorter leaf wetness periods for germination and penetration into leaves or fruits of the apple tree. This relationship was first published by MILLS and LAPLANTE (1945). Our calculations are carried out based on the publications of SCHWABE (1980). Where Mills and LAPLANTE (1945) only assumed that conidia need a certain time period of leaf wetness; SCHWABE (1980) also included the importance of the temperature in the infection process.
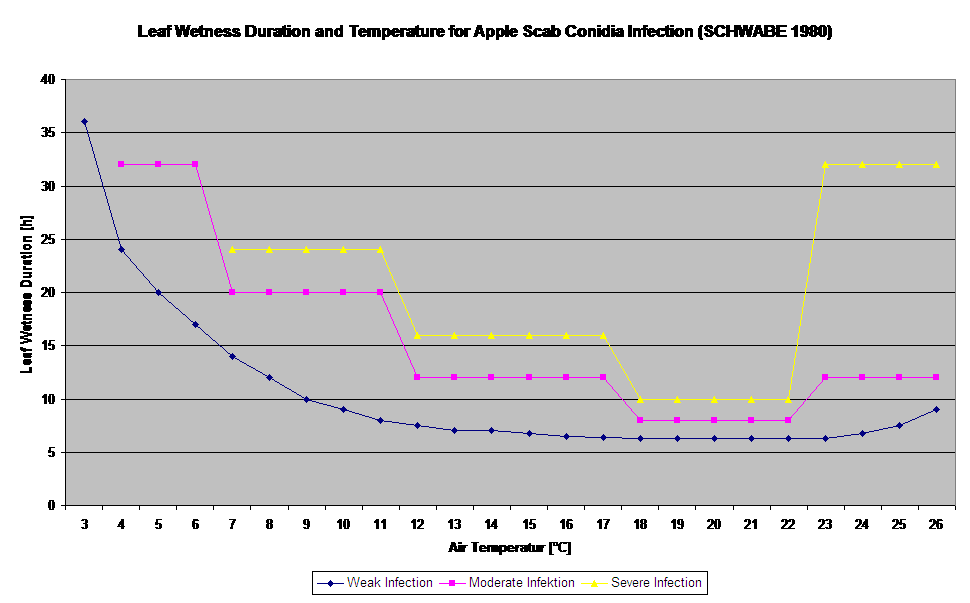
The Conidia Infection Model calculates the progress of infection depending on leaf wetness and temperature for weak, moderate and severe infections. To split the prediction into “three severity classes” is very traditional for apple scab models. The progress of infection is displayed as a graphic for hourly values. An infection has already taken place as soon as the progress curve for a weak infection reaches 100%. A moderate or severe infection is complete if the progress curves for this severity classes are reaching 100%. Often in practice or reality an infection reach 70% or 80% early in the morning and in combination with the actual weather you will be able to know if this infection will finish or not. Displaying the daily maximum values of the infection progress curve gives an overview over the expected infection dates in the season.
Literature:
- Venturia inaequalis infection, pointing out dates of ascospore and conidia infection (SCHWABE, W. 1980)
- Mac Hardy W.E., 1996. Apple scab biology, epidemiology and management. St. Paul, MN: Am. Phytopathol. Soc.: 545.
Graphical presentation of the model
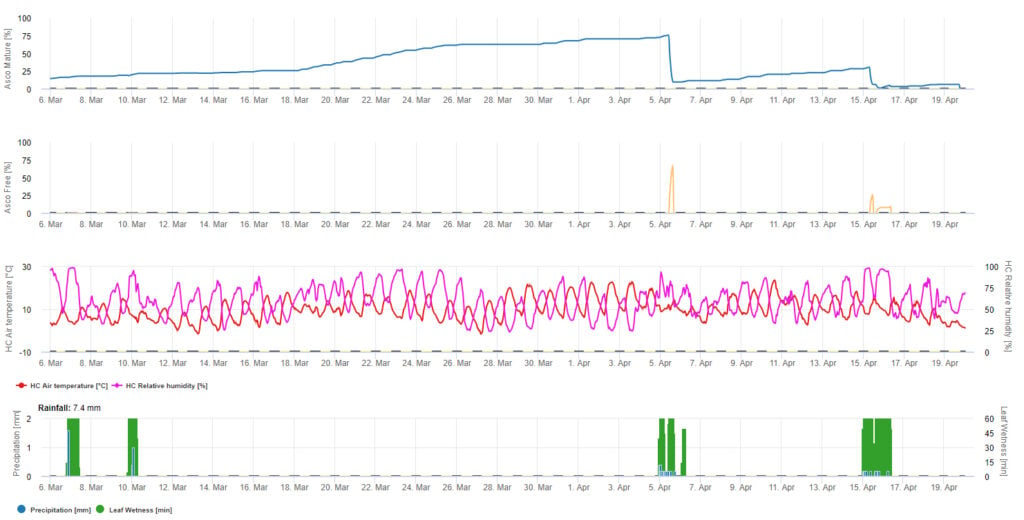
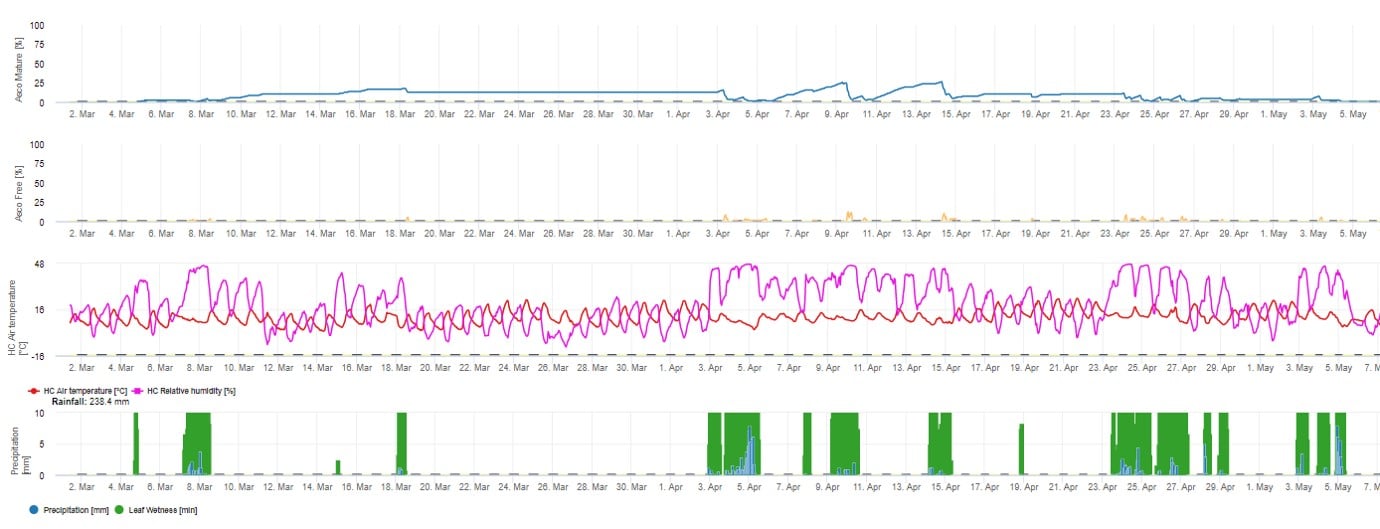
For the graphical presentation of the Ascospore maturation, formation and discharge model daily values are used. It shows that leaf wetness periods lead to ascospore releases and curves for mature and released ascospores. The severity of the release and the numbers of mature ascospores can not be assessed in absolute values. Values between 0 and 100% are indicating the relative amount of mature and released Ascospores in dependence of the climatic conditions.
1. Ascospore maturation, formation and discharge:
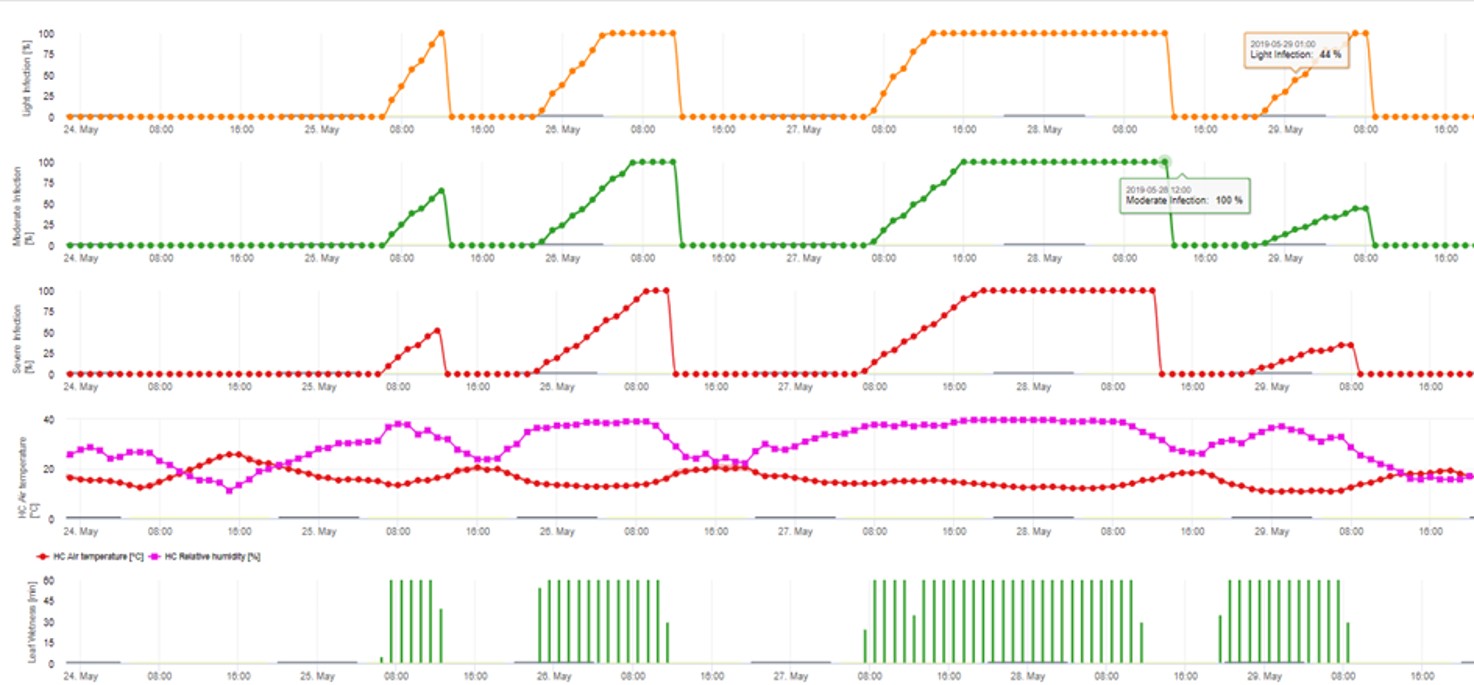
2. Ascospore Infection showing in three different severity classes (weak: orange, moderate: green, severe: red). Whenever 100% infection was reached optimal conditions for the ascospore to infect plant tissue was determined. So plant protection measurements (before infection prophylactic, after 100% infection curative) have to be taken into account:
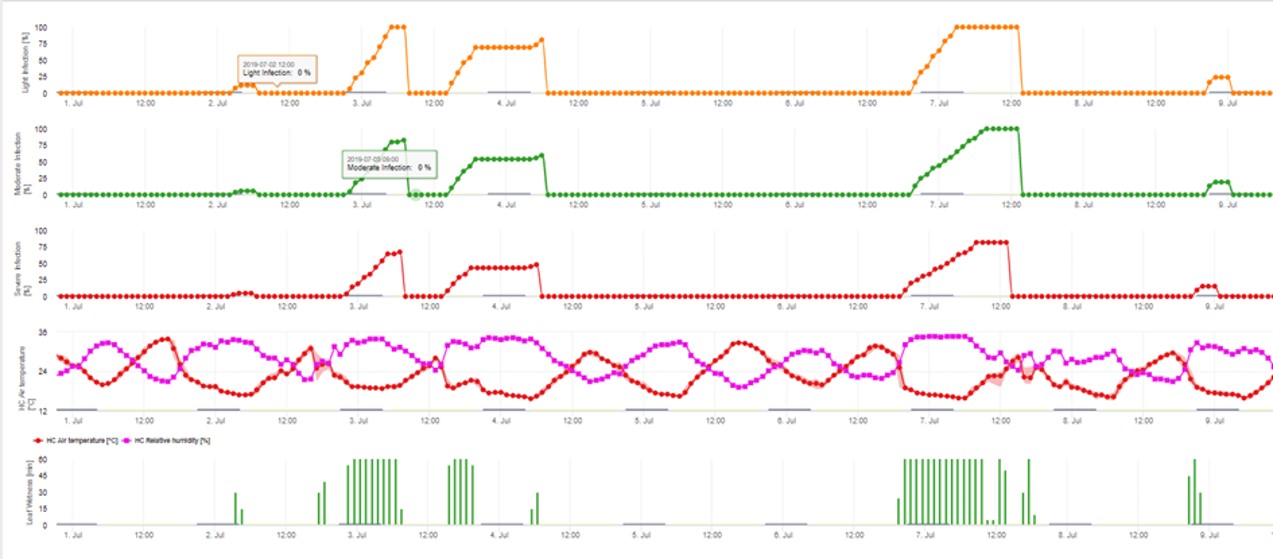
3. Conidia Infection showing in three different severity classes (weak: orange, moderate: green, severe: red). Whenever 100% infection was reached optimal conditions for the ascospore to infect plant tissue was determined. So plant protection measurements (before infection prophylactic, after 100% infection curative) have to be taken into account:
Commercially grown apples have to be free of scab. Scabbed apples will only be sold for processing. Therefore, it is the aim of all plant protection activities in conventional as well as in organic growing to have scab free fruits. Models, which show the apple scab ascospore discharge and ascospore/conidia infection are very important tools to reach this goal.
Two basic types of fungicides against apple scab are used in conventional growing systems: a) Preventative products like Captan, Mancozeb, Dithianone and Strobilurins or b) curative products like Cyprodinil (Chorus) or Pyrimethanil (Scala) or for application in the later warmer periods of the season the DMI fungicides. Actually, most of the growers follow a preventative strategy. Nevertheless, a practical preventative strategy is not able to protect apple trees completely because the apple tree grows and develop blossoms, fruits and leaves. Therefore, the preventative spray only protects for a period of 4 to 7 days in dependence of the actual growth of the tree. Such narrow spray intervals are not manageable; therefore growers will integrate their experience on the local climate, weather forecast and apple scab models in their spray management. They will schedule the preventative sprays on base of their experience and weather forecast. The apple scab infection models will show them the exact date of infection (weak, moderate and severe) as well as the ascospore/conidia discharge model and with their experience they are able to estimate the importance of an infection. This gives the possibility to act with a curative product if an apple scab infection was too long after the last preventative spray.
In organic apple production lime sulfur pointed out to be the most effective control agent against scab. The optimal control can be achieved if it is sprayed short before infection or into the beginning of infection. This has to be planned on the base of the weather forecast. Sometime we will miss this optimum period and we have to spray into the wet leaves of a nearly complete scab infection. This will still give a good efficacy. The apple scab models are helping to decide if an emergence spray into a nearly complete infection is needed.
Symptoms of Fire Blight
Overwintering cankers, harboring the fire blight pathogen, are often clearly visible on trunks and large limbs as slightly to deeply depressed areas of discolored bark, which are sometimes cracked about the margins. The largest number of cankers, however, are much smaller and not so easily distinguished. These occur on small limbs where blossom or shoot infections occurred the previous year and often around cuts made to remove blighted limbs. Since many of these cankers are established later in the season, they are not often strongly depressed and seldom show bark cracks at their margins. Also, they are often quite small, extending less than 2.5 cm, with reddish to purple bark that may be covered with tiny black fungus fruiting bodies (most notably Botryosphaeria obtusa, the black rot pathogen of apple).
Blossom blight symptoms most often appear within one to two weeks after bloom and usually involve the entire blossom cluster, which wilts and dies, turning brown on apple and quite black on pear. When weather is favorable for pathogen development, globules of bacterial ooze can be seen on the blossoms. The spur bearing the blossom cluster also dies and the infection may spread into and kill portions of the supporting limb. The tips of young infected shoots wilt, forming a very typical “shepherd’s crook” symptom. Older shoots that become infected after they have been developed about 20 leaves may not show this curling symptom at the tip. As the infection spreads down the shoot axis, the leaves first show dark streaks in the mid-veins, then wilt and turn brown, remaining tightly attached to the shoot throughout the season. As with blossom infections, the pathogen often invades and kills a portion of the limb supporting the infected shoot. The first symptom on water sprouts and shoots that are invaded systemically from nearby active cankers is the development of a yellow to orange discoloration of the shoot tip before wilting occurs. In addition, the petioles and mid-veins of the basal leaves on such sprouts usually become necrotic before those at the shoot tip.
Depending on the cultivar and its stage of development at the time infection occurs, a single blossom or shoot infection can result in the death of an entire limb, and where the central leader or trunk of the tree is invaded, a major portion of the tree can be killed in just one season. In general, infections of any type that occur between petal fall and terminal bud set usually lead to the greatest limb and tree loss. In addition, heavily structured trees tend to suffer less severe limb loss than those trained to weaker systems for high productivity. Where highly susceptible apple rootstock’s (M.26, M.9) become infected, much of the scion trunk and major limbs above the graft union very typically remain symptom less, while a distinct dark brown canker develops around the rootstock. As this rootstock canker girdles the tree, the upper portion shows symptoms of general decline (poor foliage color, weak growth) by mid to late season. In some instances, the foliage of trees affected by rootstock blight develop early fall red color in late August to early September, not unlike that often associated with collar rot disease caused by a soil borne fungus. Some trees with rootstock infections may not show decline symptoms until the following spring, at which time cankers can be seen extending upward into the lower trunk.
Disease cycle
The bacterial pathogen causing fire blight overwinters almost exclusively in cankers on limbs infected the previous season. The largest number of cankers and, hence, those most important in contributing inoculum, occur on limbs smaller than 38 mm in diameter, especially around cuts made the previous year to remove blighted limbs. During the early spring, in response to warmer temperatures and rapid bud development, the bacteria at canker margins begin multiplying rapidly and produce a thick yellowish to white ooze that is elaborated onto the bark surface up to several weeks before the bloom period. Many insect species (predominantly flies) are attracted to the ooze, and subsequently disperse the bacteria throughout the orchard. Once the first few open blossoms are colonized by the bacteria, pollinating insects rapidly move the pathogen to other flowers, initiating more blossom blight. These colonized flowers are subject to infection within minutes after any wetting event caused by rain or heavy dew when the average daily temperatures are equal to or greater than 16 °C while the flower petals are intact (flower receptacles and young fruits are resistant after petal fall). Once blossom infections occur, early symptoms can be expected with the accumulation of at least 57-degree days (DD) greater than 13 °C which, depending upon daily temperatures, may require 5 to 30 calendar days.
With the appearance of blossom blight symptoms, the number and distribution of inoculum sources in the orchard increase greatly. Inoculum from these sources is further spread by wind, rain, and many casual insect visitors to young shoot tips, increasing the likelihood for an outbreak of shoot blight. Recent research conducted in Pennsylvania indicates that aphid feeding does not contribute to shoot blight. More research is needed to determine whether or not leafhopper’s play a role in the incidence of shoot blight. Most shoot tip infections occur between the time that the shoots have about nine to ten leaves and terminal bud set, when sources of inoculum and insect vectors are available, and daily temperatures average 16 °C or more.
In years when blossom infections do not occur, the primary sources of inoculum for the shoot blight phase are the overwintering cankers and, in particular, young water sprouts near these cankers, which become infected as the bacteria move into them systemically from the canker margins. Such systemic shoot infections, called canker blight, are apparently initiated about 111 DD greater than 13 °C after green tip, although visible symptoms may not be apparent until the accumulation of at least 157 DD greater than 13 °C after green tip. In the absence of blossom infections, the development of shoot blight infections is often localized around areas with overwintering cankers.
Model blossom blight
The model requires the user to recognize specific and ever-changing local events and aspects of their orchard that may increase or decrease fire blight risk relative to other orchards in the region. The model requires the user to assume there is a risk of fire blight infection whenever blossoms are present on the trees, especially during the petal fall and “post bloom” period, when scattered blossoms may remain on many apple and pear varieties. The model user is asked to carefully assess the situation on their specific site and to initiate control measures if blossoms are present, risk levels are “High” or “Extreme,” and blossom wetting is likely to occur sometime during the next 24 hours.
Model Structure: Temperatures and Wetness: The key Fire Blight process that must be modeled is the potential for bacterial growth on the stigmas of apple and pear flowers. This growth is temperature dependent, so dependable prediction of infection risk requires the use of a measurement method that most accurately reflects the growth of Erwinia amylovora colonies. The main disagreement among modelers is how this should be done.
The Cougar blight model estimates bacterial growth rate with degree hours based on a specific growth rate curve. This growth curve is based on the growth rate of E. amylovora bacteria in laboratory tests. The degree hour values are accumulated each hour of the day that temperatures are over 15 °C. The hourly values increase as temperatures rise from 15 °C to 29°C, decline at higher temperatures, and reach zero for any hour with temperatures over 40°C.
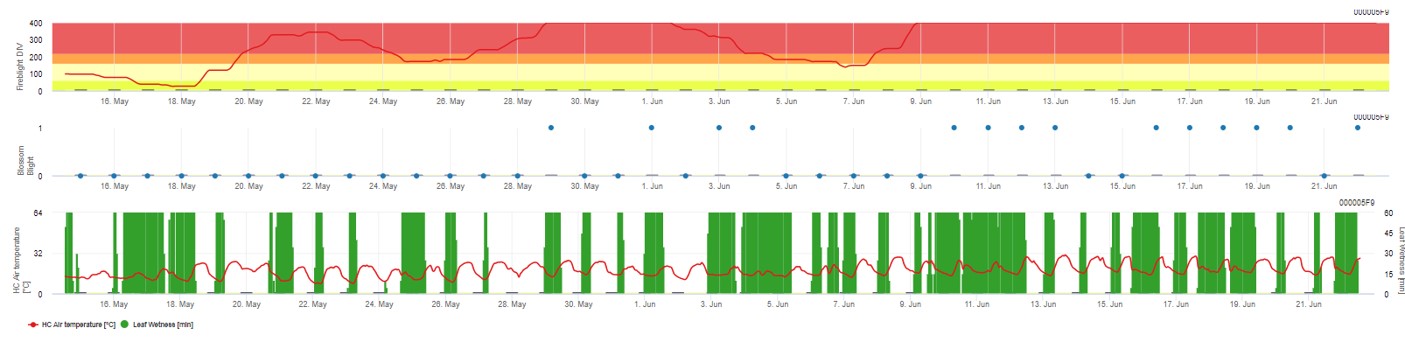
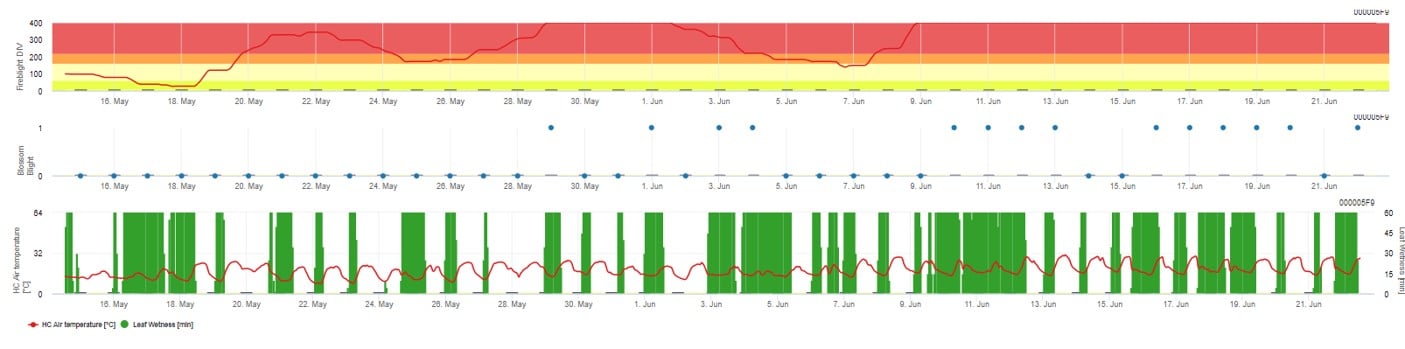
In FieldClimate.com both models are displayed in the same figure. The Fireblight DIV model is based on Courgarblight and the Blossomblight Model is based on Maryblight.
Model cougar blight
The incidence of blossom blight is one of the most sporadic aspects of fire blight epidemics. The model was built on the assumption that there is an abundance of inoculum and that, for a blossom infection event to occur, four strict conditions must be met in sequence. These conditions are:
1) flower must be open with stigmas and petals intact, stigmas have to be exposed for colonization, flowers in petal fall are resistant;
2) accumulation of at least 110 °C hours > 18.3°C within the last 44°C days > 4.4°C defines the epiphytic infection potential for the oldest open and hence most colonized flower in the orchard
3) a wetting event occurring as dew or 0.2 mm of rain or 2.5 mm of rain the previos day allows movement of bacteria from colonized stigmas to the nectarthodes
4) an average daily temperature of >= 15.6°C: This may influence the rate at which the bacteria migrate into the nectarthodes as well as the multiplication of bacteria needed to establish infections.
Graphical presentation of the models
Both models are displayed on the same page. The graphical presentation of the Fire Blight models is best to use with values aggregated on the daily base. The graph shows the climate values related to this disease. The Cougar Blight result named Fire Blight DIV and the Blossom Blight results. To interpret the Cougar Blight results the graph is underlaid by 5 different colors. The distribution of this colors is made on base of the settings on the first blight history of the orchard (see button on the right upper side) . The 5 colors are indicating the risk class for the DIV values.
Interpretation of values (in dependence of the fire blight occurrence the last years, basic set up):
Active cankers present nearby
Set the settings for fire blight history with the blue bottom “Settings” on the right upper side:
Displaying the results of the Fire Blight model by 1) Modell Cougar Blight (called Fireblight DIV based on history and background colors) and 2) Modell Blossom Blight
| Potential for Pathogen Presence | Low | Moderate | High | Extreme |
| Potential for Pathogen Presence | Low | Moderate | High | Extreme |
| No fireblight past two years | 0-350 | 350-500 | 500-800 | 800+ |
| Fire blight in local area two seasons | 0-300 | 300-500 | 500-750 | 750+ |
| Fire blight in local area two seasons | 0-250 | 250-450 | 450-700 | 700+ |
| Fire blight in orchard last year | 0-200 | 200-350 | 350-500 | 500+ |
Action threshold:
Control is advised if a high or extreme risk infection period is detected. Factors determining the damage are: numbers of flowers, tree aged, vigor and variety,… too.
The opportunity of an Blossom Blight infection is displayed by a bar from 0- 1 (so all conditions are met or not).
Aim of the Fire Blight models is to assess the probability of an infection by Erwinia amyloflora in the orchard.
The Mary Blight model which is evaluating for blossom blight is very well indicating infection situations of high economic impact. By this reason it is quite frequent used to indicate the use of antibiotics against this pathogen. Cougar blight is giving information about the risk of fire blight infections do to the overall propagation possibilities of the pathogen. Its weighting done by the history of an orchard is very helpful to indicate us how carefully we have to check the orchards for fire blight symptoms even in situations where Mary Blight will not indicate an infection.
Biology of Codling moth
The C. pomonella spends the winter as a mature larva spun up in a cocoon that is located on protected areas on the tree (bark crevices or cracks), which is the preferred location, or at the base of the tree in ground litter. In modern high-density apple orchards there are very few cocooning sites on trees because of the smooth bark. Pupae form in the cocoon spun by the mature larva.
The adult stage emerges from the pupal case and after a short period to harden the exoskeleton it flies into the tree. Most literature indicates that moth activity occurs in the upper third of the tree canopy. Adults are active at dusk for a couple hours with an average activity threshold of between 13°C to 15°C. Mating usually occurs on the host and egg laying occurs directly afterwards. It is possible on warm evenings for a female moth to emerge, mate, and lay eggs in a two-day period. A C. pomonella moth can live over a 20-day period under ideal conditions, however in fields it is likely they survive a much shorter time, especially in the hot summer. Under laboratory conditions a female moth lays up to 100 eggs, but the actual fecundity in the field is likely less and most are laid in the first few days after emergence and mating.
Eggs are laid individually on the upper surface of the leaf, on twigs or on fruit. In the first-generation eggs are laid only on leaves until about two weeks after fruit have set when the fuzziness of the fruit wares off. In the second generation about 65% of the eggs are laid on leaves but these are usually very close to a fruit. Eggs are creamy white when first laid, have a red-ring mid-way though development and when near hatch the black head capsule can be seen.
A newly hatched larva wander until they locate a fruit. It chews through the skin and feeds on the apple flesh beneath the skin for a few days. In the process of chewing through the skin and feeding beneath the larvae pushes frass out the entry area. There is often a red ring surrounding the entry, especially in young fruit. After the larva molts to the next stage it bores toward the core where it will feed on seeds until mature. When nearly mature the larva will make a tunnel from the core to the exterior of the apple and will plug the hole with frass (excrement) and silk. When ready to spin a cocoon, the larva leaves the fruit, wondering across the tree or dropping to the ground on a silk thread, and finds a protected location.
Model for Codling moth
Our model is based on the accumulation of the effective temperatures between >= 10°C and <=32°C. In the Northern hemisphere this accumulation starts with the first of January. In the Southern hemisphere it starts at the first of July. We accumulate every measurement in its equivalent for the day (see graph below). The following thresholds are used for the insect development:
*Complete Generation= 650
| *Egg Laying to Egg Hatch | = 88 |
| *First Larvae Stage | = 60 |
| *Second Larvae Stage | = 65 |
| *Third Larvae Stage | = 55 |
| *Forth Larvae stage | = 45 |
| *Fifth Larvae Stage plus Pupae | = 335 |
The graph produced by the model displays the first appearance and duration of the larvae instar 1 to 5 and the presence of adults in 5 levels.
| *Level 1: | Flight activity, non-egg laying |
| *Level 2: | Flight activity with possible egg laying. |
| *Level 3: | Flight activity with some egg laying. |
| *Level 4: | Flight activity with reasonable to strong egg laying. |
| *Level 5: | Flight activity with strong egg laying. |
Presentation of the Codling Moth Adult stages in 5 different levels (first graph) as well as the accumulated degree days (red line, second graph) for adult development and the larval stages shown in blue (third graph).
Several aphid species are present in apple orchards. The green apple aphid, the Oat apple aphid or Apple-grass aphid and the Rosy apple aphid. In the private gardens mostly, rosy apple aphids have some importance. This aphid leads to heavy hormonal changes in the tips of the shoots with infestations. The leaves become curled and the shout becomes shorted. Later the leaves become yellow and die. This aphid is mostly affecting young shoots. The Rosy apple aphid can produce severe damage already in a small abundance. Whereas the green apple aphid and the apple grass aphid can occur in higher abundance without leading to economic damage. The apple grass aphid has to be rated to be of non-economic importance at all.
Rosy apple aphid
This is an old European species introduced into the United States about 1870. It became a major pest of apples near the end of the 19th century. To thrive, this species must have an abundance of its summer host plant, the narrow-leaved plantain. A remarkable parallelism exists between the introduction and spread of rosy apple aphid and the narrow leaved plantain. This aphid occurs throughout all fruit-growing areas worldwide.
Life stages
Egg: The egg is oval and slightly flattened on the side next to the bark. The length varies from 0.49 to 0.56 mm. When first laid, it is bright yellow and covered with a glutinous substance that hardens with age. The color gradually changes to greenish yellow and finally to a shiny jet black. The time required for this change in color varies under normal outdoor conditions from about nine days to more than two weeks.
Nymph: The individuals that hatch from the eggs are all viviparous wingless females. There are five instars. The last is the mature stem mother, which shortly after the fourth instar, begins to produce living young parthenogenetically. She produces an average of five to six young per day. The nymphs of the second generation, all of which are females, reach maturity in two to three weeks. The great majority of the nymphs begin to reproduce on apple, though a few may develop wings and migrate to the plantain.
The third generation is produced in June and early July. The majority of this generation develop wings and migrate to the narrow-leaved plantain. In some seasons, wingless females of the third generation produce a fourth generation on apple. In recent years, it has been observed in some areas that damaging populations of rosy apple aphid have persisted in orchards until midsummer or later. It is not known whether this change is due to the selection process, changes of habit or lack of need for an alternative host.
Adult: The adult varies considerably in color markings. The general color is rosy brown, with a pinkish cast due to a powdery covering. Some of the older adults are purple, while the younger adults are decidedly reddish pink.
Host range
Apple is the preferred host, but the aphid also feeds on pear and hawthorn. Cortland, Golden Delicious, Rhode Island Greening and Ida Red are all particularly susceptible apple varieties. The aphid is found in all fruit-growing areas of the United States and Canada.
Injury or Damage: Rosy apple aphid feeding often causes apple leaves to curl, starting at petal fall. These leaves may later turn bright red. Feeding on the leaves around fruit clusters often results in the bunching, stunting and malformation of the developing fruit. These abnormalities become worse as fruit develops and can eventually render the fruit unsalable.
Large aphid populations may produce large amounts of honeydew as waste from the sap on which they feed. Honeydew excreted onto fruit will serve as a growing ground for sooty mold fungus, which will affect the finish of the apple. Toxins in the aphids’ saliva also serve as a “stop drop,” preventing the fruits’ abscission (natural separation from the tree) at normal harvest.
Life History: The aphid passes the winter in the egg stage. Hatching occurs early in the spring, about a week to 10 days later than the apple grain aphid and at about the same time as the apple aphid. The eggs hatch when the buds start opening in the spring, over a period of two weeks. As soon as they hatch, the young seek out the opening buds of the apple; they seem to prefer the fruit buds. They feed on the outside of the leaf bud and fruit bud clusters until the leaves begin to unfold. Then they work their way down inside the clusters and begin sucking the sap from the stems and newly formed fruits.
Their feeding causes the leaves to curl, protecting the aphids from sprays and some enemies. The severe curling of the foliage caused by this species is probably the most characteristic feature of its work. A single stem mother located on the underside of a leaf near the midrib will cause the leaf to fold tightly. It takes only a few stem mothers to cause a severe curling of all leaves surrounding an opening flower bud, providing ideal protection to the rapidly developing aphids. The stem mothers reach maturity when apple trees are coming into bloom.
The mature stem mothers are very inactive. They settle down and feed and produce young at a rapid rate. When distributed, they quickly remove their beaks from plant tissues and seek out another spot in which to continue. The stem mothers mature about two weeks after hatching. The length of time depends largely on weather conditions. The production of young usually begins two or three days after the last molt and continues without interruption for more than a month.
The total production by a single female averages about 185. Normally, the period of reproduction extends from about early May to June. Usually, the maximum period of reproductive activity is around the last week of May and the first week of June, when the young fruits are beginning to set and start active growth. Rosy apple aphid is rarely found attacking the young and rapidly growing shoots. It restricts itself to the foliage, the flower stalks and the young fruits.
One of the characteristic features of this species is the congregation of the young about the mother. Each individual stem mother or group of mothers will have massed about it hundreds of young. The infested leaves may soon be covered-in some cases, by more than one layer of aphids. This habit of congregating soon kills the infested leaves and causes the forced migration of the aphids. The young move actively and hurriedly, seemingly anxious to locate a suitable feeding ground. They are frequently found during this period congregated on the forming fruits or attacking the new, succulent unfolding foliage.
The second generation required four to 40 days to reach maturity and produce young. The majority of the second generation is wingless females. The average total production of each individual in the second generation about 119. The habits and activities of the third generation do not differ from those of the second. The aphids congregate in immense numbers on the undersides of the foliage, causing severe curling. They also attack the setting and developing fruits, producing characteristic injuries.
The majority of this third generation acquires wings and migrates to narrow-leaved plantain, the summer host plant. A fourth generation may be produced. All of these produce wings and migrate to the narrow-leaved plantain. After the last molt, the winged adults are very tender and inactive. They remain secreted in the curled leaves for two or three days before venturing on their migratory flight. Just before flying, they become very active and nervous, running about or moving their wings up and down in anticipation of their flight.
Green apple aphid
The green apple aphid, Aphis pomi (DeGeer), is widely distributed in all apple growing areas. In the northern hemisphere it first appears in apple orchards in late May to early June. The insect sucks sap from the leaves on water sprouts and succulent terminal growth. Green apple aphids are usually found close to the major veins on the under surface of the leaf. The green apple aphid also attacks pear, hawthorn, quince, crab apple and spiraea.
Description of green apple aphid
The eggs of the green apple aphid are oval and shiny black. The nymphs and adults are dark or light green. The green apple aphid has a green head, uniformly green or yellowish green abdomen, black legs and long black cornicles – the “tail pipes” (at the rear end of the insect). Adult females are dark green to black and winged. During the summer these females, or alates, give birth to live young parthenogenically.
Biology of green apple aphid
The green apple aphid overwinters as an egg on suckers, at the base of buds and on leaf scars of terminal shoots. The eggs usually hatch when the buds burst and the first leaves are unfolding. The nymphs begin to feed immediately on the developing leaves. They are initially present on the terminal shoot and later move to older cluster leaves. Adult female aphids produce wingless females in two weeks, each female being capable of producing 50-100 live offspring. Young aphids develop in seven to ten days. Green apple aphid populations usually build on apple slowly in early spring (bloom, petal, fall), and more rapidly as average daily temperatures increase. Winged aphids, or alates, and wingless aphids are produced during the summer. Green apple aphids are most numerous during July and early August. There are more nymphs that alate adults in the initial stages of an infestation. The percentage of alates increases as the population grows. Green apple aphid colonies can double in a week if conditions are favorable. If the aphids do not have succulent new growth and they have to feed on older leaves, then the number of young produced can drop by up to 50%. If daily temperatures are 30-32°C and higher, females do not reproduce well. When temperatures are high for several days, aphids will die. Several generations of the green apple aphid develop during the summer, and winged forms disperse throughout the orchard. Sometimes the green apple aphid and the rosy apple aphid, Dysaphis plantaginea Pars., have colonies on the same leaf. The rosy apple aphid is pink or purple. In the fall, wingless males and females develop and mate, and the females lay eggs for the winter. The greatest numbers of eggs are 15-20cm from the tips of twigs. Eggs are seldom on the large scaffold limbs or on the trunks of apple trees.
Damage
The green apple aphid sucks sap from the leaf. Heavy infestations reduce vigor and growth of shoots. This is of particular concern in nurseries and young, non-bearing orchards. The insect can reduce bud size and internode length, and cause leaf curling. Aphid feeding may also stimulate lateral branch growth that can affect tree shape. Leaf curling and weakened terminals are susceptible to winter injury. Honeydew produced by the aphids may drip onto fruit allowing sooty fungi to grow. The fungus blemishes the fruit and lowers the market value. When infestations are heavy, the green apple aphid may feed on immature apples and cause russeting.
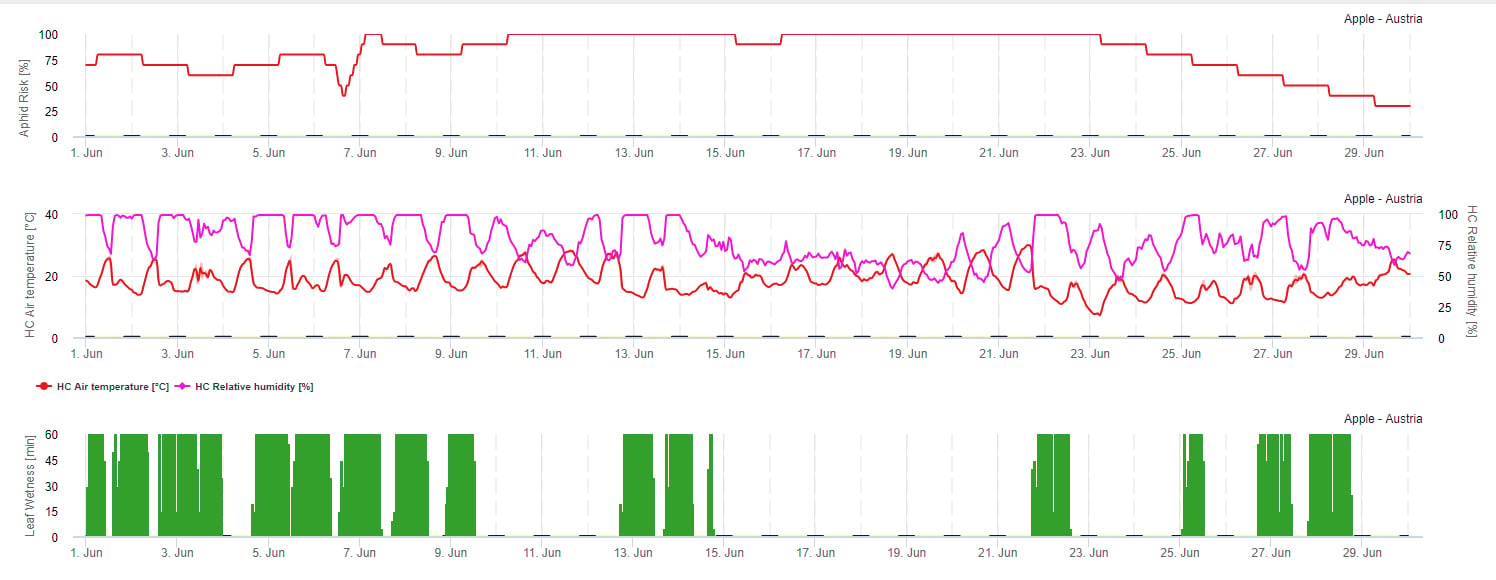
Propagation risk model
Aphid propagation is mainly influenced by temperature. As more hours are close to the optimum temperature (20-32°C) and there is a food source available as higher will be the propagation rate. Whereas eggs and young instars are susceptible against dry air. Low relative humidity’s can damage aphid populations too (below 30% r.h.). Beside of insecticides, aphids are controlled by predators and parasitic fungi. The parasitic fungi will be depending on moisture. Most of this species like Beauvaria bassiana are favored by free moisture.
We increase risk when the optimal temperature is between 20 and 32°C and relative humidity between 30 and 95% r. h. and decrease the risk when it is too wet (leaf wetness, rain, or relative humidity above 95%), too cold (temperatures lower than 20°C) or too hot (temperatures above 32°C), wet nights (leaf wetness in night hours).